
Nasrollah Vali
Department of Animal Sciences, Faculty of Agriculture, Islamic Azad University, Shahrekord Branch, Iran
Asian Journal of Poultry Science
Year: 2010 | Volume: 4 | Issue: 2 | Page No.: 60-66







ABSTRACT
A total of 210, 3-d-old Japanese quail chicks (Coturnix japonica) allocated to 21 cages, each cage containing 10 chicks, that were received seven diets with three replication of each diet. Experimental diets were arranged with five levels 150, 300, 600, 1200 and 2400 phytase enzyme unit (FTU kg-1) with commercial name (Natuphos 500) as treatments1, 2, 3, 4 and 5, respectively and two control groups (positive and negative controls). At the end of each experimental period (each week to 45 days of ages), birds were weighed and feed consumption was recorded for feed efficiency computation. At the age of 45 days, four chicks were randomly selected from each treatment blood samples for subsequent determination of minerals (Ca, P and Mg) and alkaline phosphates (ALP) in serum. Data were subjected to analysis of variance using the General Linear Models (GLM) procedures of SAS software. Data showed that feed efficiency was not affected at periods of 9-16 and 37- 45 days of ages by addition of phytase, but in other periods were significantly different (p<0.05). Liver weight in treatment 5 (4.68±0.23 g) at 45 days of age was more than other groups that had significant difference (p<0.05). Quails of treatment 4 had Calcium (Ca) and Phosphorus (P) 10.19±0.80, 7.11±0.46 mg dL-1, respectively that were more than other groups. The amount of Magnesium on treatment 1(3.27±0.15 g), positive control (3.27±0.15 g) and negative control (3.56±0.15 g) which were not significantly different (p>0.05), whereas treatment1 and control groups had significant differences with other treatments (p>0.05). The Alkaline phosphates of treatment 5 (366.83±38.07 IU L-1) in blood serum at 45 days of age represents the most increase among other treatments, which had significant differences with treatment 1 (124.99±50.80 IU L-1) and control groups, although weren’t significant difference with the treatments 2 (260.26±50.01 IU L-1) and treatments 3 (288.37±40.64 IU L-1).
PDF Abstract XML References Citation
How to cite this article
Nasrollah Vali, 2010. Comparison Difference Levels of Phytase Enzyme in Diet of Japanese Quail (Coturnix japonica) and Some Blood Parameters. Asian Journal of Poultry Science, 4: 60-66.
DOI: 10.3923/ajpsaj.2010.60.66
URL: https://scialert.net/abstract/?doi=ajpsaj.2010.60.66
DOI: 10.3923/ajpsaj.2010.60.66
URL: https://scialert.net/abstract/?doi=ajpsaj.2010.60.66
INTRODUCTION
The primary constituents of diets for poultry are plant-based ingredients which come primarily from the seeds of plants. Most of the stored phosphorus in plants is found in seeds mainly as a component of a molecule called phytin. Phytin-phosphorus is poorly available to poultry and this availability varies both within and among ingredients. The enzyme phytase releases phosphate groups from phytin potentially making this released phosphorus available to the animal. Phytase is the only recognized enzyme that can initiate the release of phosphate from phytin (International Union of Biochemistry, 1979). Nelson et al. (1968) demonstrated that the addition of the enzyme phytase to grains and feeds was an effective way to increase phosphorus (P) availability to poultry.
Numerous reports have shown that phytate phosphorus present in cereal grains could be made available to both poultry and swine by treating the grains or supplementing the diets with the enzyme phytase, reducing the need to supplement these diets with inorganic P and decreasing P excretion (Jongbloed and Kemme, 1990; Simons et al., 1990; Zhu et al., 1990; Ketaren et al., 1993).
The P is an essential mineral for laying hens in formation of egg shell and metabolism (Frost and Roland, 1991; Summers, 1995; Usayran and Balnave, 1995). Only 20-50% of plant P is available to broilers and the rest of P is in the form of phytate (myo-inositol hexaphosphate), which is poorly used by broilers (Ravindran et al., 1995). Ravindran et al. (1998) and Sebastian et al. (1998) reported that poultry can not produce enough amounts of endogenous phytase to hydrolyze and release P from phytate. To meet dietary P requirement of laying hens, inorganic P (Dicalcium phosphate) or exogenous phytase enzymes are commonly added to commercial corn-soy layer diets. However, Dicalcium phosphate supplementation is not only expensive but also leads to environmental problems by over-supplementation. Excess P from the feces of hens is easy to access ground water, rivers, lakes and oceans and can lead to mortality of aquatic animals by stimulating algae growth.
Considerable research has demonstrated that phytase supplementation (from 100 to 300 phytase unit (FTU kg-1 of feed) to diets containing 0.1% dietary nonphytate phosphorus (NPP) has positive effects on egg production, egg mass, egg weight, egg specific gravity, bone ash and eggshell quality by improving P utilization (Boling et al., 2000a, b; Jalal and Scheideler, 2001; Roland et al., 2003; Keshavarz, 2003). Phytase supplementation decreased P excretion in the manure and reduced the potential environmental problems (Jalal and Scheideler, 2001; Biehl and Baker, 1997).
Through consideration authentic resources the phytase enzyme was narrow in diets of quail so, the target of this study is comparison difference levels of Phytase enzyme in diet of Japanese quail (Coturnix japonica) for investigation of feed consumption and some blood factors.
MATERIALS AND METHODS
A total of 210, three days old Japanese quail chicks (Coturnix japonica) were housed in pen with the newspaper litter until 12 days of age. Then birds were moved into one-tier cage (60x50x40 cm) in an environmentally controlled room, with 23 h constant overhead fluorescent lighting. The temperature was decreased gradually from 35 to 20°C on the weekly bases. The chicks were allocated to 21 cages, each cage containing 10 chicks, that were received seven diets with three replication of each diet. The experimental diets are presented in Table 1 and were formulated to meet the requirements of quail as established by the NRC (1994). Experimental diets were arranged with five levels 150, 300, 600, 1200 and 2400 phytase enzyme unit (FTU kg-1) with commercial name (Natuphos 500) as treatments1, 2, 3, 4 and 5, respectively and two control groups (positive and negative controls). One FTU is defined as the amount of enzyme that liberates micromole inorganic phosphorus per minute from 0.0051 mol L-1 sodium phytate at 37°C and at pH = 5.5 (Hall et al., 2003). Small amounts of the basal diet were first mixed with the respective amounts of phytase as a small batch and then with a larger amount of the basal diets until the total amounts of the respective diets were homogeneously mixed. Diets in mash form and water were provided ad libitum from 3 to 45 days of ages. At the end of each experimental period (each week to 45 days of ages), birds were weighed and feed consumption was recorded for feed efficiency computation.
| Table 1: | Ingredients and nutrient composition of the experimental diets |
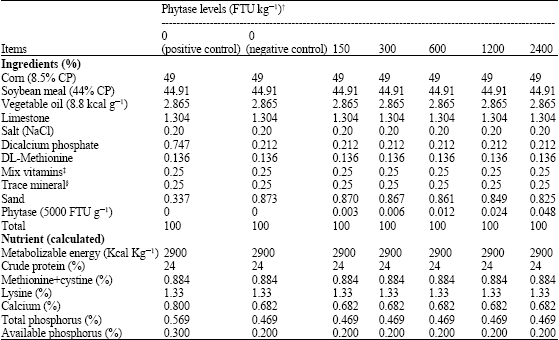 | |
†Danisco animal nutrition, Carol stream, IL, FTU kg-1 feed = phytase units/kg feed. One phytase unit (FTU) is defined as the amount of enzyme that liberates micromole inorganic phosphorus per minute from 0.0051 mol L-1 sodium phytate at 37°C and at pH = 5.5. ‡Provided per kilogram of diet: vitamin A (as retinyl acetate), 8,000 IU: Cholecalciferol, 2,200 ICU: Vitamin E (as DL-a-tocopheryl acetate), 8 IU: Vitamin B12, 2800.02 mg: Riboflavin, 5.5 mg: D-calcium pantothenic acid, 13 mg: Niacin, 36 mg: Choline, 500 mg: Folic acid, 0.5 mg: Vitamin B1 (thiamin mononitrate), 1 mg: Pyridoxine, 2.2 mg: D-biotin, 0.05 mg: Vitamin K (menadione sodium bisulfate complex), 2 mg.§Trace mineral mix provided the following (per kg of diet): manganese (MnSO4_H2O), 60 mg: Iron (FeSO4_7H2O), 30 mg: Zinc (ZnO), 50 mg: Copper (CuSO4_5H2O), 5 mg: Iodine (ethylene diamine dihydroiodide), 1.5 mg | |
At the age of 45 days, four quails were randomly selected from each treatment and weighed and were killed, bled and plucked for liver weight and blood samples for subsequent determination of minerals (Ca, P and Mg) and alkaline phosphates (ALP) in serum. A total 84 blood samples (8-10 cc) collected for to assigning Ca, p, Mg elements by photometric system and alkaline phosphates enzyme (ALP) by photometric system by method Deutsche Gesellschaft für klinische Chemie (1972).
Data were subjected to analysis of variance using the general linear models procedures of SAS software (SAS Institute, 1998). Statements of statistical significance are based on a probability of p<0.05.
RESULTS AND DISCUSSION
The effects of phytase enzyme on growth performance are summarized in Table 2. The main effects data indicated that the body weights at 23 and 45 days of ages weren’t significantly different and at the other ages (16, 30 and 37 days of age) were significantly different, but in all stages positive control with other experiment treatments weren’t significantly different (p<0.05). Body weight at 45 days of age in treatment of 5 was 173.77±13.40 g that showed more than other treatments and treatment of 3 in the same age with 156.00±13.40 g was the lowest among other treatments (Table 2). Data also showed that feed efficiency (Table 3) was not affected at periods of 9-16 (FE1) and 37- 45 (FE5) days of ages by addition of phytase, as a result of simultaneous showed in weight gain, but in other periods were significantly different (p<0.05). In this study, feed efficiency weren’t different between the positive and negative control groups (Table 3) in any periods (p<0.05).
| Table 2: | Least square means for effect treatments and sex with standard error for Body weight average (g) at age of 16 (BW1), 23 (BW2), 30 (BW3), 37 (BW4) and 45 (BW5) days of ages |
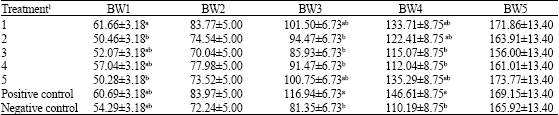 | |
Values with in a column with no common superscript differ significantly (p<0.05) and columns without superscript were not significantly different.1Experimental diets were arranged with five levels 150, 300, 600, 1200 and 2400 phytase enzyme unit (FTU kg-1) with commercial name (Natuphos 500) as treatments 1, 2, 3, 4 and 5, respectively and two control groups (positive and negative controls) | |
| Table 3: | Least square means for effect treatments with standard error for feed efficiency (g g-1) at periods of 9-16 (FE1), 16-23 (FE2), 23- 30 (FE3), 30 37 (FE4) and 37- 45 (FE5) days of ages |
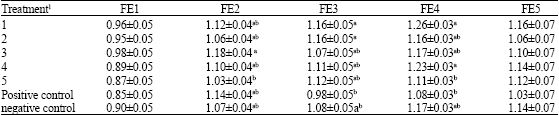 | |
Values with in a column with no common superscript differ significantly (p<0.05) and columns without superscript were not significantly different. 1Experimental diets were arranged with five levels 150, 300, 600, 1200 and 2400 phytase enzyme unit (FTU kg -1) with commercial name (Natuphos 500) as treatments1, 2, 3, 4 and 5, respectively and two control groups (positive and negative controls) | |
| Table 4: | Least square means with standard error for Body weight average (BW), liver weight (lw), Calcium (Ca), Phosphorus (P), Magnesium (Mg) and Alkaline phosphates (ALP) at 45 days of age |
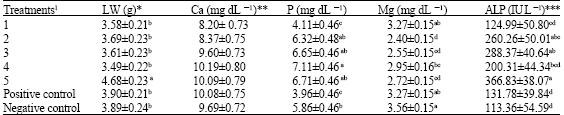 | |
abcdValues with in a column with no common superscript differ significantly (p<0.05) and column without superscript was not significantly different.1Experimental diets were arranged with five levels 150, 300, 600, 1200 and 2400 phytase enzyme unit (FTU kg-1) with commercial name (Natuphos 500) as treatments1, 2, 3, 4 and 5, respectively and two control groups (positive and negative controls). One FTU is defined as the amount of enzyme that liberates micromole inorganic phosphorus per minute from 0.0051 mol L -1 sodium phytate at 37°C and at pH = 5.5. *(g) gram, **(mg dL -1) mille gram per deciliter, *** (IU L -1) international unit per liter | |
These results were similar to those obtained by Perney et al. (1993), Sebastian et al. (1996a, b), Ahmad et al. (2000) and Viveros et al. (2002) those have reported on broiler. Although, Rama Rao et al. (1999) reported improved feed efficiency with supplemental phytase. These contrasting results may be due to a number of factors including phytase source, ingredients (type, source, phytate content) and dietary characteristics (processing, Vitamin D3 level, Ca:P ratio) (Ravindran et al., 1995).
The effect of these treatments on Liver Weight (LW), Calcium (Ca), Phosphorus (P), Magnesium (Mg) and alkaline phosphates (ALP) blood serum at 45days of age are presented in Table 4. Liver weight in treatment 5 (4.68±0.23 g) at 45 days of age was more than other groups that had significant difference (p<0.05). Quails of treatment 4 had Calcium (Ca) and Phosphorus (P) 10.19±0.80, 7.11±0.46 mg dL-1, respectively that were more than other groups. The main effects of data indicated that the increase in phytase content in the diet were not significantly different (p>0.05) for Ca retention at 45 days of age, but for P retention were significantly different (p<0.05) (Table 4). The amount of Magnesium on treatment 1 (3.27±0.15 g), positive control (3.27±0.15 g) and negative control (3.56±0.15 g) which were not significantly different (p>0.05), whereas treatment1 and control groups had significant differences with other treatments (p<0.05) (Table 4). The Alkaline phosphates of treatment 5 (366.83 ± 38.07 IU L-1) in blood serum at 45 days of age represents the most increase among other treatments, which had significant differences with treatment 1 (124.99 ± 50.80 IU L-1) and control groups, although, weren’t significant difference with the treatments 2 (260.26±50.01 IU L-1) and treatments 3 (288.37±40.64 IU L-1) (Table 4). These results were similar to those obtained by Saricicek et al. (2005) in laying quails, but more than those obtained by Can et al. (2007) in Japanese quails. The decrease in serum alkaline phosphates associated with the diets supplemented with phytase might reflect the down regulation of this enzyme resulting from the increased availability of phosphorus (Huff et al., 1998).
Phytates (Phytic acid, mineral complexes), as well as very limited providing P availability to the animal in the absence of phytase, can also have other effects which can reduce nutrient digestibility and consequently, animal performance (i.e., phytates in feed act as anti-nutrients). At neutral or basic pH, phytic acid can form complexes with divalent and trivalent cations. These minerals and trace elements attach themselves easily onto the phosphate molecule of phytic acid producing insoluble salts. Once these compounds are produced these cations are unavailable for absorption in the gut and, like phosphorus, these valuable nutrients are then also lost to the animal. If these insoluble salts react with fatty acids in the gut, insoluble metallic soaps are produced. It has been suggested that these soaps may lower fat digestibility and consequently may have an effect on energy availability. Phytic acid can also form complexes with proteins and starch. These can occur in the plant (native complexes) or during digestion in the gastrointestinal tract. In the highly acidic stomach region, amino acids are bound directly to phytate phosphorus creating an insoluble complex. In the less acidic intestine, multiple bonded mineral cations act as a bridge between phytate and protein. These complex compounds are more resistant to enzymatic digestion than their unbound counterparts. Moreover, phytates can also potentially decrease protein and starch digestibility by inhibiting, or binding to, digestive enzymes produced by the animal (e.g., pepsin and trypsin for protein degradation and alpha-amylase for starch degradation). The ability of phytase to improve P availability by hydrolyzing phytate-bound P in poultry diets is well documented (Broz et al., 1994; Coelho and Kornegay, 1996; Yi et al., 1996; Qian et al., 1996, 1997). Increased utilization of P from phytate can therefore reduce supplementation of diets with inorganic P sources while maintaining normal growth of the bird.
REFERENCES
- Ahmad, T., S. Rasool, M. Sarwar, Ahsan-ul Haq and Zia-ul Hasan, 2000. Effect of microbial phytase produced from a fungus Aspergillus niger on bioavailability of phosphorus and calcium in broiler chickens. Anim. Feed Sci. Technol., 83: 103-114.
CrossRefDirect Link - Biehl, R.R. and D.H. Baker, 1997. Microbial phytase improves amino acid utilization in young chicks fed diets based on soybean meal but not diets based on peanut meal. Poult. Sci., 76: 355-360.
CrossRefPubMedDirect Link - Boling, S.D., M.W. Douglas, R.B. Shirley, C.M. Parson and K.W. Koelkebeck, 2000. The effect of various dietary levels of phytase and available phosphorus on performance of laying hens. Poult. Sci., 79: 535-538.
PubMedDirect Link - Boling, S.D., M.W. Douglas, X. Wang and C.M. Parsons, 2000. The effect of dietary available phosphorus and phytase on performance of young and old laying hens. Poul. Sci., 79: 224-230.
Direct Link - Broz, J., P. Oldale, A.H. Perrin-Voltz, G. Rychen, J. Schulze and C.S. Nunes, 1994. Effects of supplemental phytase on performance and phosphorus utilisation in broiler chickens fed a low phosphorus diet without addition of inorganic phosphates. Br. Poult. Sci., 35: 273-280.
CrossRefDirect Link - Can, A., M. Avci, N. Denek and O. Kaplan, 2007. Effects of microbial phytase on growth performance, carcass yield, biochemical parameters, oxidative stress and faecal phosphorus content of Japanese quails. J. Anim. Vet. Adv., 6: 1031-1035.
CrossRefDirect Link - Deutsche Gesellschaft fur klinische Chemie, 1972. Empfehlungen der deutschen Gesellschaft fur Klinische Chemie (DGKC). Standardisierung von Methoden zur Bestimmung von Enzymaktivitaten in biologischen Flussigkeiten. [Recommendation of the German Society of Clinical Chemistry. Standardization of methods for measurement of enzymatic activities in biological fluids]. Z Klin Chem. Klin Biochem., 10: 182-192.
- Frost, T.J. and D.A. Roland Sr., 1991. The influence of various calcium and phosphoras levels on tibia strength and eggshell quality of pullets during peak production. Poult. Sci., 70: 963-969.
CrossRefPubMedDirect Link - Hall, L.E., R.B. Shirley, R.I. Bakalli, S.E. Aggrey, G.M. Pesti and H.M.Jr. Edwards, 2003. Power of two methods for the estimation of bone ash of broilers. Poult. Sci., 82: 414-418.
Direct Link - Huff, W.E., P.A. Moore Junior, P.W. Waldroup, A.L. Waldroup and J.M. Balog et al., 1998. Effect of dietary phytase and high available phosphorus corn on broiler chicken performance. Poult. Sci., 77: 1899-1904.
CrossRefDirect Link - Jalal, M.A. and S.E. Scheideler, 2001. Effect of supplementation of two different sources of phytase on egg production parameters in laying hens and nutrient digestibility. Poult. Sci., 80: 1463-1471.
CrossRefDirect Link - Jongbloed, A.W. and P.A. Kemme, 1990. Effect of pelleting mixed feeds on phytase activity and the apparent absorbability of phosphorus and calcium in pigs. Anim. Feed Sci. Technol., 28: 233-242.
CrossRefDirect Link - Keshavarz, K., 2003. The effect of different levels of nonphytate phosphorus with and without phytase on the performance of four strains of laying hens. Poult. Sci., 82: 71-91.
CrossRefDirect Link - Ketaren, P.P., E.S. Batterham, E.B. Dettmann and D.J. Farrell, 1993. Phosphorus studies in pigs. 3. Effect of phytase supplementation on the digestibility and availability of phosphorus in soya-bean meal for grower pigs. Br. J. Nutr., 70: 289-311.
CrossRefDirect Link - Yi, Z., E.T. Kornegay, V. Ravindran and D.M. Denbow, 1996. Improving phytate phosphorus availability in corn and soybean meal for broilers using microbial phytase and calculation of phosphorus equivalency values for phytase. Poult. Sci., 75: 240-249.
CrossRefDirect Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA.
Direct Link - Nelson, T.S., T.R. Shieh, R.J. Wodzinski and J.H. Ware, 1968. The availability of phytate phosphorus in soybean meal before and after treatment with a mold phytase. Poult. Sci., 47: 1842-1848.
PubMedDirect Link - Perney, K.M., A.H. Cantor, M.L. Straw and K.L. Herkelman, 1993. The effect of dietary phytase on growth performance and phosphorus utilization of broiler chicks. Poult. Sci., 72: 2106-2114.
CrossRefPubMedDirect Link - Qian, H., H.P. Veit, E.T. Kornegay, V. Ravindran and D.M. Denbow, 1996. Effects of supplemental phytase and phosphorus on histological and other tibial bone characteristics and performances of broilers fed semi-purified diets. Poult. Sci., 75: 618-626.
PubMedDirect Link - Qian, H., E.T. Kornegay and D.M. Denbow, 1997. Utilization of phytate phosphorus and calcium as influenced by microbial phytase, cholecalciferol and the calcium: Total phosphorus ratio in broiler diets. Poult. Sci., 76: 37-46.
CrossRefDirect Link - Rao, S.V.R., V.R. Reddy and V.R. Reddy, 1999. Enhancement of phytate phosphorus availability in the diets of commercial broilers and layers. Anim. Feed Sci. Technol., 79: 211-222.
CrossRefDirect Link - Ravindran, V., W.L. Bryden and E.T. Kornegay, 1995. Phytates: Occurrence, bioavailability and implications in poultry nutrition. Poult. Avian Biol. Rev., 6: 125-143.
Direct Link - Roland, Sr. D.A., H.A. Ahmad, S.S. Yadalam and T. Sefton, 2003. Effect of nongenetically modified phytase supplementation on commercial Leghorns. J. Applied Poult. Res., 12: 257-263.
CrossRefDirect Link - Saricicek, B.Z., U. Kilic and A.V. Garipoglu, 2005. Replacing soybean meal Sbm by canola meal CM the effects of multi enzyme and phytase supplementation on the performance of growing and laying quails. Asian-Aust. J. Anim. Sci., 18: 1457-1463.
Direct Link - Sebastian, S., S.P. Touchburn, E.R. Chavez and P.C. Lague, 1996. The effects of supplemental microbial phytase on the performance and utilization of dietary calcium, phosphorus, copper and zinc in broiler chickens fed corn-soybean diets. Poult. Sci., 75: 729-736.
PubMed - Sebastian, S., S.P. Touchburn, E.R. Chavez and P.C. Lague, 1996. Efficacy of supplemental microbial phytase at different dietary calcium levels on growth performance and mineral utilization of broiler chickens. Poult. Sci., 75: 1516-1523.
PubMed - Sebastian, S., S.P. Tounchburn and E.R. Chavez, 1998. Implication of phytic acid and supplemental microbial phytase in poultry nutrition a review. World's Poult. Sc. J., 54: 27-47.
Direct Link - Simons, P.C.M., H.A.J. Versteegh, A.W. Jongbloed, P.A. Kemme and P. Slump et al., 1990. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Br. J. Nutr., 64: 525-540.
CrossRefDirect Link - Usayran, N. and D. Balnave, 1995. Phosphorus requirements of laying hens fed on wheat based diets. Br. Poul. Sci., 36: 285-301.
PubMedDirect Link - Zhu, X.S., P.A. Seib, G.L. Allee and Y.T. Liang, 1990. Preparation of a low-phytate feed mixture and bioavailability of its phosphorus to chicks. Anim. Feed Sci. Technol., 27: 241-251.
CrossRefDirect Link