
D.O. Aworinde
Department of Biological Sciences, University of Agriculture, Abeokuta, Ogun State, Nigeria
D.U. Nwoye
Department of Biological Sciences, University of Agriculture, Abeokuta, Ogun State, Nigeria
A.A. Jayeola
Department of Botany and Microbiology, University of Ibadan, Ibadan, Oyo State, Nigeria
A.O. Olagoke
Department of Forestry and Wood Technology, Federal University of Technology, Akure, Ondo State, Nigeria
A.A. Ogundele
Department of Biology, Federal College of Education (Special), Oyo, Nigeria
Research Journal of Botany
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 17-28







ABSTRACT
A detailed morphological study of the leaf epidermis of the tropical genera of some species Acalypha, Bridelia, Euphjorbia, Hura, Jatropha, Manihot and Ricinus in Nigeria is presented. The study revealed several interesting epidermal features some of which have not previously been reported in the genera. Leaf epidermal characters such as pattern of epidermal cells, types of stomata and presence of trichomes are constant in some species and variable in others and thus of great significance in understanding the relationships between and within species. Leaves are amphistomatic in all species except in Bridelia ferruginea, Euphorbia heterophylla, Euhorbia pulcherrima and Jatropha gossypifolia which are hypostimatic. The stomata length, width, density and index also very in different species.
PDF Abstract XML References Citation
How to cite this article
D.O. Aworinde, D.U. Nwoye, A.A. Jayeola, A.O. Olagoke and A.A. Ogundele, 2009. Taxonomic Significance of Foliar Epidermis in Some Members of Euphorbiaceae Family in Nigeria. Research Journal of Botany, 4: 17-28.
URL: https://scialert.net/abstract/?doi=rjb.2009.17.28
URL: https://scialert.net/abstract/?doi=rjb.2009.17.28
INTRODUCTION
Euphorbiaceae is a big family found mostly in the tropics. It consists of trees, shrubs, herbs but are rarely woody climbers. The larger genera are: Euphorbia (about 2,000 species), Croton (700 species), Phyllanthus (500 species), Acalypha (430 species), Jatropha (175 species), Manihot (170 species) (Dutta, 1980). Representatives of these larger genera and other smaller ones are well distributed both in the free area and forest reserves across Nigeria.
In Nigeria, the indigenous people have utilized number of species in the Euphorbiaceae family for various purposes in the crude forms as well as in processed forms. Some usage, however, are hinged on superstition and thus, such usage cannot be explained beyond metaphysical level. Such prominent uses can basically be classified into food and fodder, medicine, hedges, landscape beautification, timber, superstitious use and others; this however does not dispute the fact that some of them are still among the popularly known weed of both the arable farmlands and forest plantation (Akobundu and Agyakwa, 1998).
The foliage of Acalypha ciliata and Acalypha ornata provides grazing for donkeys, cattle, sheep and goat; but horses are said not to take it. Acalypha ornata has also been reported as being useful in that the stems are woven into baskets and fish-traps in Tanganyika and Gbengi. Besides being a staple food in every household, feed for livestock, Manihot esculenta has also gained entrance into the international market and thus form a source of foreign exchange earning in Nigeria.
Many of these Euphorbiaceae species have essential medicinal importance. Acalypha ciliata is applied as dressing to wound, while A. ornata, apart from being a laxative, is also known to have healing application to circumcision wounds. Laboratory experiments on immature female guinea pigs showed that the Euphorbia hirta enhanced mammary development and secretion of milk. It is also used by women as a galactogogue in Nigeria. The plant has been an item of market trade in Northern Nigeria for this purpose. Igbo medicine-men use a cold-water extract of the bark of Bridelia ferruginea with the stem of Costus afer (Costaceae) in the treatment of osenkenu, identified as minor epilepsy. The list of the medicinal potentials of the member of this family is inexhaustible.
Also, Acalypha fimbriata, Euphorbia heterophylla and E. pulcherrima are popularly utilized for ornamental beautification of landscape. Acalypha ciliata occurrence under planted cotton may not be altogether vicarious for perhaps magical ends it is used to adopt good results in southern Nigeria (Oliver, 1960). There is a superstitious invocation not to beat a friend with stem of tapioca, for if he is a sorcerer, he will die. Manihot plant also enters into a Yoruba incantation to get money quickly.
In spite of the numerous usages to which members of Euphorbiaceae family have been put in the time past, much research have not been done on the use of anatomical features in the delineation and classification of the various taxa in this family as a whole. Previous works basically delved into the propagation, growth and physiology (David and Ruth, 1988; Kuhn et al., 1996; Schurr, 1998; Walter and Schurr, 2000; Buckeridge et al., 2000; Qin et al., 2004 etc.), developmental anatomy (Heckenberger et al., 1998; Oparka and Turgeon, 1999) and medicinal potentials of members of the family (Anselm, 2004; Tolu, 2006 etc.).
The use of data generated from leaf epidermal surfaces in resolving the taxonomy of taxa has gained much recognition for a long time. Some of the prominent researchers that have solved taxonomic problems among plant taxa with such data are Ogundipe and Olatunji (1991), Olowokudejo (1993), Das and Ghose (1993), Illoh (1995), Isawumi (1996), Croxdale (2000), Das (2002) and Ayodele and Olowokudejo (2006). The present study described the leaf epidermal morphology in some members Euphorbiaceae family with the aim of providing useful taxonomic data that would give further insight into proper classification, delineation and identification of the studied taxa.
MATERIALS AND METHODS
Sources of Material
Plant materials used for this investigation was obtained from the herbaria of the Forestry Research Institute of Nigeria, Ibadan (FHI) and Department of Botany and Microbiology, University of Ibadan (UIH). Field collections were also made. Both the herbaria and field collection were conducted between the space of April to December 2007. The field collection was ensured to cover both the rainy and dry seasons in a bid to make provision for any variation in the leaf micro-morphology that may be due to seasonal changes. All taxa studied are listed in Table 1.
Measurement of Quantitative Morphological Parameters
Twelve dried leaf materials of the Euphorbiaceae family consisting of the genus: Acalypha, Bridelia, Euphorbia, Hura, Jatropha, Manihot and Ricinus were collected from Forestry Research Herbarium, Ibadan (FHI).
Observation of Qualitative Character
The macromorphological characters were assessed on mature leaves at comparative position include leaf length, leaf width at the widest point, petiole length, blade length, leaf apex, leaf margin, leaf shape, leaf surface and leaf base, others are derived ratios of the length and width of the leaf, length and width of the petiole. Three to five specimens were assessed for the micro characters in each species based on their geographical distribution except in cases where the specimens were limited or collected from the same locality. The micro characters assessed include number of epidermal cells per view (x400), thickness of cell wall, size of epidermal cells at widest point, number of stomata per view (x400), length and width of stomata and stomata index. Others are trichome type, stomata type, shape of epidermal cells and anticlinal wall pattern.
| Table 1: | Leaf morphological feature of some species of family Euphorbiaceae |
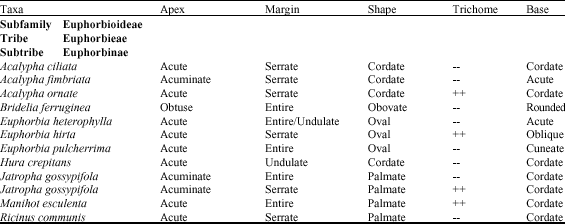 | |
| --: Denotes glabrous leaf ; ++: Denotes pubescent leaf | |
Preparation of Leaf Epidermal Surfaces
Leaf epidermal morphology was studied using fresh and dried herbarium materials. About 5 mm2-1 cm2 leaf portions were obtained from the standard median portion of the leaves.
Three to five specimens of each species were used depending on the geographical spread of the species except for those known from only one or two localities. Herbarium specimens were first dehydrated by boiling in water for about 10-29 min. The leaf fragments were then soaked in concentrated HNO3 in capped specimen bottles for a period of about 18-24 h. They were transferred into water in a Petri dish with a pair of forceps to strip off thin slices of epidermis of the leaves into water in a Petri-dish. Upper and lower epidermises were carefully and completely isolated from the mesophyll using pair of fine forceps and dissecting needles. The epidermises were cleared with a camel hair brush and placed in distilled water for rinsing for 5 min. The epidermises were later stained in aqueous safranin for 5 min. Each membrane was dehydrated by passing though different grades of alcohol viz., 30, 50, 70 and 95% for about 15 min each. After the dehydration, the membrane was stained in aqueous fastgreen for 3 sec and then cleared in xylene for about 15 min. The membranes were then placed in clove oil for 15 min and again cleared in xylene for another 15 min and then mounted on clean slides with DPX® Mountant. The slide were labeled appropriately and examined under the light microscope while photographs of the micro-morphological features were take at a magnification of x400 using a photomicrograph with an installed digital camera optics.
For statistical analysis, 25 epidermal cell and 20 stomata were chosen randomly from each taxon and measured using a micrometer eye-piece. For each quantitative character, the range, mean, standard deviation and standard error were determined for all taxa. The Stomata Index (SI) was calculated using the formula of Metcalfe and Chalk (1979):
where, S denotes the number of stomata per unit area and E is the number of epidermal cells of the same area. Measurement of the epidermal cell width was taken at the widest point on each cell.
| Table 2: | Quantitative characters for the stuied species min (mean±SE) max |
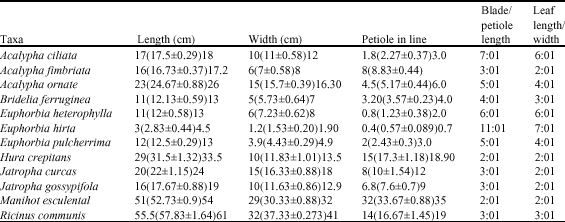 | |
RESULTS
Macromorphological Characters
The leaves are simple, opposite and mostly entire, serrate but rarely undukate. The shape may be cordate, oval, palmate but rarely obovate (Table 2). The leaves are mostly glabrous but a few are pubescent. The apices are acute or acuminate but rarely obtuse while the bases are mostly cordate but rarely acute, rounded and oblique. The leaf size shows considerable variation within the family with the largest recorded in Ricinus communis and the smallest in Euphorbia hurta. The petiole length ranges from 0.4 cm in Euphorbia hirta to 32 cm in Manihot esculenta (Table 2). The lowest blade/petiole length ratio of 2:1 was found in Hura crepitans and Manihot esculenta while the highest of 7:1 was recorded in Acalypha ciliata. The leaf length/width ratio ranges from 2:1 in many species to 7:1 in Euphorbia hirta.
Micromorphological Characters
Epidermal Cells
Leaf epidermal cells are isodiametic, irregular or more often polygonal (Table 1, Fig. 1). Anticlinal wall patterns are straight or curved irregular cells are usually restricted to the abaxial surface of most species although they occur on the adaxial surfaces in a few taxa. The irregular cells are inter mixed with polygonal cells. Isodiametric cells occur only on the adaxial surface of Bridelia ferruginea (Table 3, Fig. 1O). The polygonal and irregular epidermal cells usually have straight to curved anticlinal walls but in some few cases they have curved or curved to straight anticlinal walls (Fig. 1).
The epidermal cells are thick ranging from 7.33 μm in Acalypha fumbriata, Euphorbia pulcherrima, Jatropha curcas, Jatropha gossypifolia, Manihot esculenta and Ricinus communis to 10 μm in Acalypha ciliata on the adaxial surface. Abaxial epiderml cell walls are generally less thicker than the adaxial cell walls with Ricinus communis have the thinnest wall.
Considerable variation occurs in the number of epidermal cells on both adaxial and abaxial surfaces even within species. Acalypha fimbriate has the lowest number of epidermal cells per square millimeter in the family (94 on the adaxial surface) while Manihot esculenta has the lowest on the abaxial (79). Manihot esculenta has the largest number of epidermal cells per square millimeter with 198 on the adaxial surface while Ricinus communis has the largest number on the abaxial surface with 179. In most of the taxa, more cells occur on the adaxial surface than on the abaxial surfaces (Table 4).
| Table 3: | Epidermal characters of some species of family euphorbiaceae |
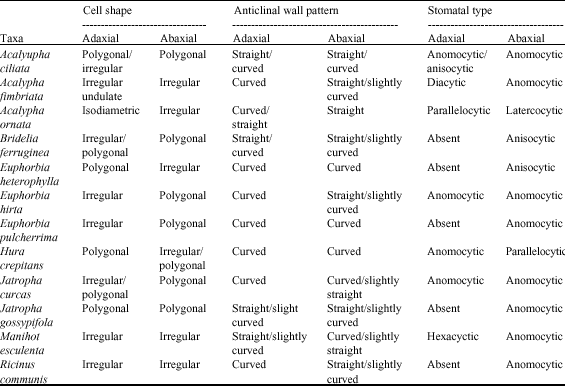 | |
| Table 4: | Variation in epidermal cell size and cell wall thickness of some species of family euphobiaceae |
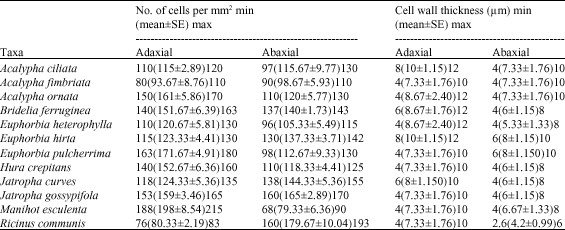 | |
Stomata Structure and Density
All species of the family are amphistomatic except for Bridelia feruginea, Euphorbia heterophylla, Euphorbia pulcherrima and Jatropha gossypifolia which are hypostomatic. A wide range of stomatal types are recorded for the family. These are, Anisocytic with three cells which may be of unequal size enclosing the guard cells, Anomocytic with epidermal cells, around the guard cells not being distinguishable from other epidermal cells, Diacytic where stomata is enclosed by one or more pairs of subsidiary cells whose common walls are at right angle to the guard cells, Hexacytic with a modified tetracytic type with an additional pair of lateral subsidiary cells, Laterocytic where stomata is flanked by three or more subsidiary cells all bordering on the lateral sides of the guard cell pair and parralelocytic with stomata having an alternating complex of three or more C-shape subsidiary cells of graded sizes parallel to the guard cells (Table 3, Fig. 1). In certain cases, more than one type of stomata may be present on the same surface of the same species e.g anomocytic and anisocytic in Acalypha ciliata (Fig. 1U, Table 3). Moreover, the stomatal types found on the adaxial surface of one species may differ from those on the abaxial surface of the same species. The most common types of stomata in the family are the anisocytic and anomocytic (Table 3, Fig. 1). In the amphistomatic taxa, stomata are more frequent on the abaxial surface. The stomata number varies from 1 in Manihot esculenta to 36 in Ricinus communis on the adaxial surface with stomata index ranging from 0.45 to 30.17% in the respective species. On the abaxial surface, the number of stomata ranges from 1 in Euphorbia heterophylla and Acalypha fimbriata to 65 in Manihot esculenta and Hura crepitans and a stomata index of 4.35 to 45.99% (Table 5).
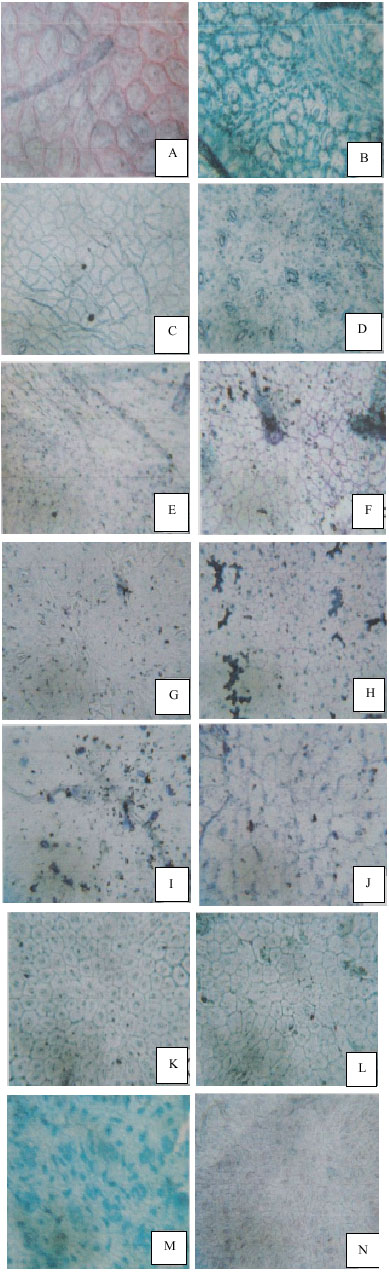 | |
| Fig. 1: | Photomicrographs of leaf surfaces of some members of Euphorbiaceae family (Epidermal cells and stomata). (A and B): Jatropha curcas. A-Adaxial surface showing anomocytic stomata and polygonal epidermal cells. B-Abaxial surface showing anomocytic stomata and polygonal epidermal cells. (C and D): Hura crepitans. C-Adaxial surface showing anomocytic stomata and polygonal epidermal cells. D-Abaxial surface showing parallelocytic stomata and polygonal to irregular shaped cells. (E and F): Euphorbia heterophylla. E-Abaxial surface showing anisocytic stomata and irregular shaped epidermal cells. F-Adaxial surface showing no stomata and polygonal epidermal cells. (G and H): Acalypha fimbriata. G-Adaxial surface showing diacytic stomata and irregular/ undulate epidermal cells. H-Abaxial surface showing anomocytic stomata and irregular epidermal cells. (I and J): Euphorbia hirta. I-Abaxial surface showing anomocytic stomata and irregular cells. J-Adaxial surface showing anomocytic stomata and irregular to polygonal shaped cells. (K and L): Euphorbia pulcherrima. K-Adaxial surface showing no stomata and polygonal epidermal cells. L-Abaxial surface showing anomocytic stomata and polygonal epidermal cells. (M and N): Ricinus communis. M-Adaxial surface showing anomocytic stomata and irregular epidermal cells. N-Abaxial surface showing anomocytic stomata and irregular epidermal cells. (O and P): Bridelia ferruginea. O-Adaxial surface showing no stomata and isodiametric cells. P-Abaxial surface showing anisocytic stomata and polygonal shaped cells. (Q and R): Manihot esculenta. Q-Adaxial surface showing hexacytic stomata and irregular to polygonal epidermal cells. R-Abaxial surface showing anomocytic stomata and irregular epidermal cells. (S and T): Jatropha gossypifolia. S-Adaxial surface showing no stomata and polygonal epidermal cells. T-Abaxial surface showing anomocytic stomata and polygonal epidermal cells. (U and V): Acalypha ciliata. U-Adaxial surface showing anomocytic to anisocytic stomata and polygonal to irregular cells. V-Abaxial surface showing anomocytic stomata and polygonal shaped cells. (W and X): Acalypha ornata. W-Abaxial surface showing laterocytic stomata and irregular epidermal cells. X-Adaxial surface showing parallelocytic stomata and irregular epidermal cells |

| Table 5: | Stomata characters of the studied taxa |
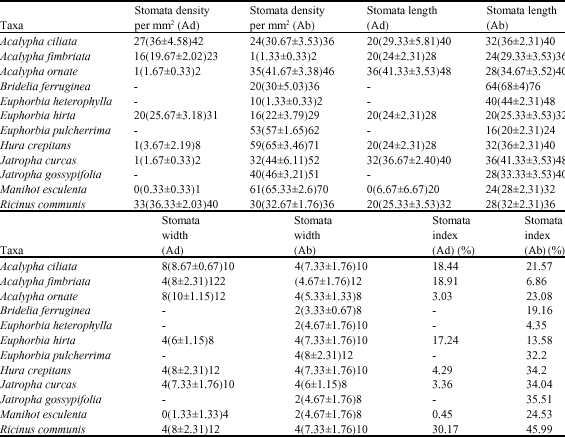 | |
| All measurements in μm min (mean±SE) max, -: Denotes no data; Ad: Denotes adaxial; Ab: Denotes abaxial | |
Mean stomata length is in the range of 6.67 and 41.33 μm in Manihot esculenta and Acalypha ornate, respectively on the adaxial surface. On the abaxial surface, the mean stomata length ranges from 20.00 μm in Euphorbia pulcherrima and Bridelia ferruginea respectively (Table 5). Mean stomata width varies from 1.333 to 8 μm in Bridelia ferruginea and Euphorbia pulcherrimma respectively (Table 5).
Trichomes
Most of the species are glabnrous while a few representatives including Acalypha ornata, Euphorbia hirta, Jatropha gossypifolia and Manihot esculenta are pubescent (Table 1).
Long uniseriate flagelliform trichome (with long whip-like structures) is characteristics of Bridelia ferruginea. Short trichomes are characteristic of Euhorbia heterophylla, Euphorbia hirta and Acalypha ciliate, Euphorbia hirta and Manihot esculentus. Two-to-five armed trichomes are present in Jatropha gossypifolia. Thread like trichomes are also present in Euphorbia heterophylla and Jatropha curcas. The presence of striations is more on the adaxial surfaces of most species and is significant in the fact that they vary in form ranging form slightly undulate to deeply undulate.
DISCUSSION
According to Webster (1987), the taxonomic position of Euphorbiaceae which continues to generate controversy is not unconnected with the great diversity within this family. This is demonstrated in the possession of substantial family characteristics, such as latex, number of seeds/ carpels, basic chromosomes number and so on. The classification dilemma necessitates the quest for additional taxonomic and phylogenetic criteria to improve our understanding of the systematics of this family (Jensen et al., 1994).
In spite of the fact that, vegetative and floral characters are markedly modified in relation to the habitat and pollination mechanisms (Kapil and Bhatnagar, 1994), the preceding observations and the summaries of character variation in Table 3 and 5 indicated that the taxonomic application of the diversity of epidermal morphology in Euphorbiaceae cannot be overemphasized. The cell shape and cell wall pattern vary considerably within the geenra. Based on these two characters, the genera can be conveniently divided onto four groups. Bridelia ferrugunea which has isodiametric and polygonal shape on the adaxial and abaxial surfaces respectively can be separated easily from other species. Also, Euphorbia pulcherrima, Jatropha gossypifolia, Jatropha curcas and Acalypha ciliata with polygonal cell shape and curved wall pattern on both epidermises are distinctly different from other species. Acalypha fimbriata, Acalypha ornata, Euphorbia hirta, Manihot esculentus and Ricinus communis with irregular cell shape and straight to slightly curved or curved wall pattern on both or either side can be separated from other species. However, different combinations of cell shape and anticlinal wall pattern occur in the remaining two-Euphorbia heterophylla and Hura crepitans.
The occurrence of curved walls in some of the species agreed with the suggestion of Stace (1965) that curved wall is a mesomorphic character and that environmental conditions such as humidity play a significant role in determining the pattern of anticlinal cell walls. The cell wall thickness varies form 4.2 to 8 μm and 7.33 to 10 μm on the abaxial and adaxial surfaces, respectively, while the number of epidermal cells per square millimeter varies from 80 to 191 and 79 to 179 on the adaxial and abaxial surfaces respectively. Based on these two characters, the species cannot be easily separated.
The relative abundance and variation of the trichomes is of taxonomic significance in the genera. Almost all the species in their various genera have simple unicellular trichomes either on the abaxial or adaxial surfaces of both making them difficult to be distinguished from one another. However, the presence of simple, long, interwoven trichomes which covers the epidermal cells and stomata in Euphorbia hirta make them easily distinguished from others. In Euphorbia pulcherima, the many short trichomes makes it easy to be separated from other species in the genera. Although, the absence of trichomes in some species is of little importance in distinguishing them. Metcalfe and Chalk (1979) hold that trichome frequency and size are environmentally controlled, while Stace (1965) believes that hairs are constant in a species when present and showed a constant range of form and distribution useful in diagnosis.
The presence of both parallelocytic and Laterocytic stomata in Acalypha ornate distinguishes it from other species. Also, the presence of diacytic and hexacytic stomata in Acalypha fimbriata and Manihot esculenta respectively, distinguishes them form other species. Stomata type is of no diagnostic importance in other species except Bridelia ferruginea, Euphorbia heterophylla and Ricinus communis (which have anisocytic stomata ontheir abaxial surfaces) because they all have anomocytic stomata. However, stomata index could be used to separate the species into the three groups. Those with stomata index less than 10% on the adaxial surface include Acalypha ornate, Hura crepitans, Jatropha curcas and Manihot esculenta while on the abaxial, Acalypha fimbriata and Euphorbia heterophylla are the only two species. Acalypha ciliata, Acalypha fimbriata and Euphorbia hirta have stomata index less than 20% on the adaxial surface while Bridelia ferruginea, Euphorbia hirta and Ricinus communis have stomata index less 20% on the abaxial surface. The last group are those with stomata index greater than 20% and only Ricinus communis falls into this group on the adaxial surface while Acalypha ciliata, Acalypha ornate, Euphorbia pulcherrima, Hura crepitans, Jatropha curcas, Manihot esculenta and Jatropha gossypifolia fall into the group on the abaxial surface.
Ricinus communis and Acalypha ciliata with stomata density per square millimeter of 36.33 and 36, respectively, can be separated from the remaining species on the adaxial surface while Manihot esculentus and Hura crepitans with 65.33 and 65 can be separated from the remianbing species on the abaxial surface. Also, stomnata length values are found to be reliable in distinguishing between Manihot esculenta with 6.67 μm from all other species which Bridelia ferruginea with 68 μm can be distinguished from other which have values lower than that on the abaxial surface. Also, the stomata width value can be used to distinguish Manihot esculena with 1.33 μm from all other species on the adaxial surface with values greater than its value while Bridelia ferniginea with 3.33 μm can be distinguished form others with greater values on the abaxial surface. The importance of stomatal complex in taxonomy especially in the identification of small leaf fragments, the state at which most plant specimens occur in West African herbal markets, has been emphasize by Stace (1965).
CONCLUSION
Leaf epidermal characters are of taxonomy significance in the members of some genera of the family Euphorbiaceae. With this, they can be separated and distinguished based on their stomata, epidermal cells and trichomes as these features which are being influenced by environmental factors are present on almost every leaf surface. Therefore, the stomata, epidermal cells and trichomes are micromorphological features on leaves epidermal surfaces and can be used to identify, separate or distinguish different plant species hence epidermal studied of leaf surfaces are recommended for different plants for the purpose of distinguishing them from other species.
REFERENCES
- Ayodele, A.E. and J.D. Olowokudejo, 2006. The family of polygonaceae in West Africa: Taxonomic significance of leaf epidermal characters. South Afr. J. Bot., 72: 442-459.
CrossRef - Buckeridge, S.M., N.K. Suda and F.J. Giorgini, 2000. Cell wall hydraolases in the seed of Euphorbia heterophylla during germination and early seedling development. Brazilian J. Plant Physiol., 15: 3-3.
CrossRef - Das, S., 2002. On the ontogeny of stomata and glandular hairs in some Indian mangroves. Acta Botanica Croatica, 61: 199-205.
Direct Link - David, W. and S. Ruth, 1988. Axillary bud inhibition induced by young leaves or bract in Euphorbia pulcherrima. Ann. Bot., 62: 435-440.
Direct Link - Illoh, H.C., 1995. Foliar epidermis and petiole anatomy of four species of Celosia L. in Nigeria. Feddes Repertorium, 106: 15-23.
CrossRef - Jensen, U., I. Vogel-Bauer and M. Nitschke, 1994. Leguminlike proteins and the systematics of euphorbiaceae. Ann. Missouri Bot. Gard., 81: 160-179.
Direct Link - Kapil, R.N. and A.K. Bhatnagar, 1994. The contribution of embryology to the systematics of the euphorbiaceae. Ann. Missouri Bot. Gard., 81: 145-159.
Direct Link - Ogundipe, O.T. and O.A. Olatunji, 1991. The leaf anatomy of the species of Cochlospermum Kunth. (Cochlospermaceae) in West Africa. Feddes Repertorium, 102: 183-187.
CrossRef - Olowokudejo, J.D., 1993. Comparative epidermal morphology of West African species of Jatropha L. (Euphorbiaceae). Bot. J. Linnean Soc., 3: 139-154.
CrossRef - Wei, Q., W.D. Lu, Y. Liao, S.L. Pan, Y. Xu, L. Tang and F. Chen, 2004. Plant regeneration from epicotyl explant of Jatropha curcas. J. Plant Physiol. Mol. Biol., 30: 475-478.
PubMedDirect Link - Schurr , U., U. Heckenberger, K. Herdel, A. Walter and R. Feil, 2000. Leaf development in Ricinus communis during drought strees: Dynamics of growth processes, of cellular structure and of soil-source. J. Exp. Bot., 51: 1515-1529.
CrossRef
darlington nwoye Reply
well taken and explanatory...please keep it up!