
K. N. Opara
Department of Zoology, University of Uyo, Uyo, Nigeria
I. A. Atting
Department of Medical Microbiology, College of Health Sciences, University of Uyo, Uyo, Nigeria
I. G. Ukpong
Department of Animal and Environmental Biology, University of Port Harcourt, Port Harcourt, Nigeria
A. A. Nwabueze
Department of Animal Science and Fisheries, Delta State University, Asaba, Nigeria
I. I. Inokon
Department of Zoology, University of Uyo, Uyo, Nigeria
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 3 | Page No.: 452-456







ABSTRACT
We assessed the relationship between genetic indices and their susceptibility to malaria in children. A total of 372 children (210 clinically ill and 162 healthy) attending University of Uyo Teaching Hospital, Uyo, Nigeria, were screened for malaria parasitaemia, blood group, haemoglobin genotypes, haemoglobin concentration (Hb) and packed cell volume (PCV), using standard parasitological and haemotological methods of diagnosis. One hundred and fifty six (74.3%) of the clinically-ill children examined were parasitaemic for falciparum malaria. There was no significant difference between infection and sex. There was significant (p<0.05) difference between prevalence of malaria in the children and the expected. Children with genotype AA (92.3%) were more susceptible to malaria parasite than AS (5.1%) and SS (2.6%). The association of haemoglobin genotype with malaria was highly significant (p<0.001). The degree of susceptibility of different blood groups to malaria infection were blood groups O (46.2%); A (26.9%); B (21.8%); AB (5.1%). There was a significant (p<0.05) association between the blood group and malaria infection. Children with malaria parasitaemia had a significantly (p<0.001) lower haemoglobin concentration and packed cell volume than children without parasitaemia.
PDF Abstract XML References Citation
How to cite this article
K. N. Opara, I. A. Atting, I. G. Ukpong, A. A. Nwabueze and I. I. Inokon, 2006. Susceptibility of Genetic Indices to Falciparum Malaria in Infants and Young Children in Southern Nigeria. Pakistan Journal of Biological Sciences, 9: 452-456.
DOI: 10.3923/pjbs.2006.452.456
URL: https://scialert.net/abstract/?doi=pjbs.2006.452.456
DOI: 10.3923/pjbs.2006.452.456
URL: https://scialert.net/abstract/?doi=pjbs.2006.452.456
INTRODUCTION
Malaria is still regarded as a major public health problem in the tropics. It is holoendemic in tropical Africa, claiming over 870,000 lives annually, which totals up to 90% of death attributed to malaria world wide (Claire, 1997; WHO, 2000). Malaria is one of the major causes of morbidity and mortality in African children, most of these deaths occur in the under-fives while, all other age groups suffer serious health and socio-economic consequences (Lackritz et al., 1992; Brewster et al., 1990; Ebrahim, 1978; WHO, 1987). Plasmodium falciparum is the main parasite causing severe illness and presenting mainly with febrile convulsion and severe anaemia (Binka and Adongo, 1997).
There is an increasing interest in research on the association of genetic indices such as ABO blood group, haemoglobin genotypes and diseases. Blood group and haemoglobin genotypes have been known to predispose individuals to or protect them from various diseases or infections (Omoregie et al., 2002) Individuals of blood groups A and B were reported to have shown greater susceptibility in three epidemics of influenza A than those of other groups (Frolov et al., 1976) People with blood groups A and B are known to be more susceptible to eosinophilia (Anand, 1961). Wood and Harrison (1972) reported that anopheles mosquito tends to bite groups O and B persons in preference to group A persons. It has also been reported that group B confers a selective advantage in relation to malaria infection and this partly explains why the distribution of group B tends to correspond with a high degree of malaria endemicity (Athreya and Corriell, 1967). It has been reported by Miller et al. (1973) that the predilection of malaria parasite for different blood groups may depend on the presence of specific molecules on the red cell surface. Plasmodium falciparum is able to invade many different types of human blood groups or red cells, with the exception of Ein(a) erythrocyte which lacks glycophorin A, the major sialogly co-protein on the surface of the normal red cell (Jungery, 1985). No correlation was found between blood group and malaria (Miller et al., 1976; Facer and Brown, 1979; Kazim and Ejezie, 1982).
Population field and clinical studies have provided convincing evidence that where P. falciparum malaria is endemic, non-immune subjects who have sickle-cell traits Hb-AS and Hb-SS have a survival advantage over subjects who have only normal adult haemoglobin Hb-AA as the presence of sickle haemoglobin Hb-S in red cells limit the development and multiplication of the parasite (Luzatto, 1979). The World Health Organisatiopn has defined severe anaemia complicating falciparum malaria as a haemoglobin concentration <50g L-1 or a haematocrit <15% in the presence of a parasitaemia >10,000 μL with a normocytic blood film (Warrell et al., 1990). One of the difficulties in defining anaemia caused by malaria in endemic areas is that a large proportion of children (up to 100%) have parasitaemia without clinical symptoms (Newton et al., 1997).
Although it is widely known that malaria is a severely infectious disease responsible for a great deal of morbidity and mortality among different age groups all over the world. There is a dearth of information on the relationship between malaria and genetic indices of children in Uyo, Nigeria. In this report we present the findings on the susceptibility of genetic indices to malaria parasite in children from Uyo metropolis, Nigeria.
MATERIALS AND METHODS
The study was carried out in Uyo, South Eastern Nigeria. The town serves the dual purpose of a local government headquarters and capital of Akwa Ibom State. It lies within the tropical rainforest belt of the country on Lat 5°201 and 5°32 East of the Greenwich Meridian. There are two distinct seasons namely the wet season (March-October) and dry season (November-February). The detailed description of the study area has been reported previously (Opara et al., 2004).
Study population: The study was conducted between March, 2003 and October 2003, in Uyo, Nigeria. Two Hundred and ten infant and children aged between 1 month to 16 years attending the University of Uyo Teaching Hospital who presented cases of symptoms of malaria, such as fever, headache, joint pains and cold were examined for malaria parasite, while 162 were healthy aparasitaemic children of the same age bracket. This group acted as control, since they were afebrile, not on routine antimalaria prophylaxis and showed no parasites in their peripheral blood films.
Blood collection: Blood samples for parasite estimation, blood group determination, haemoglobin estimation, genotypes assay and packed cell volume, were collected from the children using standard venepuncture method as described by Cheesbrough (1992). About 2 mL of blood was collected from each subject and dispensed into EDTA bottle. For children between the ages of 1 month and 4 years thumb-pricking method was employed.
Preparation of thick film: A drop of various blood was placed on the slide and a thick film was made air dried and read after application of the Giemsa staining method. The slides were read in accordance with the WHO (1991) protocol.
Haemoglobin (Hb) concentration: The haemoglobin concentration was measured spectrophometrically by the Cyanmethahaemoglobin method (Hall and Malia, 1991). About 0.02 mL of well mixed blood was added to 4 mL of modified drabkin solution. The mixture was allowed to stand for 3 min and the Hb concentration was read spectrophotometrically.
Blood group: Blood groups were determined using the commercially available anti-A, anti-B and anti-D blood grouping sera as described by Manson et al. (1977).
Genotypes: Haemoglobin electrophoresis was performed on cellulase acetate as previously reported by Dacie and Lewis (1991).
Packed cell volume: Packed cell volume was determined according to the method of Dacie and Lewis (1991).
Ethical clearance: Informed consent of the management of University of Uyo Teaching Hospital and parents of the children was sought and obtained.
Statistical analysis: Differences in proportion were analysed statistically using the Chi-square tests.
RESULTS
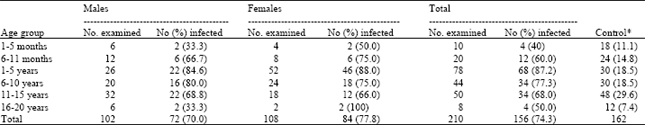
Prevalence: Out of the 210 children examined for malaria parasites 156 (74.3%) were infected. Females (77.8%) were more infected than males (70%). It was not significant (χ2 = 0.73; (Table 1). The highest prevalence of (87.2%) was observed in 1-5 years age group. The least prevalence of 40% was observed in 1-5 months age groups (Table 1).
Blood group susceptibility: The degree of susceptibility of the different blood group to malaria infection shows that blood group O (46.2%) was the highest, while the least was observed among children with blood group AB (5.1%). There was a significant association between ABO blood group and malaria infection (χ2 = 9.32, p<0.05) (Table 2).
| Table 1: | Sex and age distribution of Malaria parasites in 210 children screened and control |
 | |
| Table 2: | ABO blood groups in Southern Nigerian children with and without malaria infection |
 | |
| Numbers in parenthesis represent percentages | |
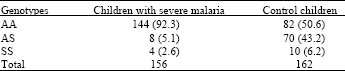
| Table 3: | Haemoglobin genotypes in Southern Nigerian children with and without malaria infection |
 | |
| Numbers in parenthesis represent percentages | |
| Table 4: | Haemoglobin concentrations in Southern Nigeria children with and without malaria infection |
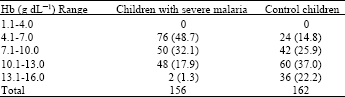 | |
| Numbers in parenthesis represent percentages | |
| Table 5: | Packed Cell Volume (PCV%) in Southern Nigerian children with and without malaria infection |
 | |
| Numbers in parenthesis represent percentages | |
Haemoglobin genotypes: The degree of susceptibility of different genotypes to malaria are AA (92.3%); AS (5.1%); SS (2.6%). There was a significant (χ2 = 28.72; p<0.001) association in the degree of susceptibility (Table 3).
Haemoglobin concentration:
Normal range (10.5-14.0 g dL-1)
Children with haemoglobin concentration of 4.1-7.0 g dL-1 had the highest rate (48.7%) of malaria infection followed by Hb of 7.1-10.0 (32.1%) and 10.1-13.0 g dL-1 (17.9%), while the least rate of infection was observed in Hb concentration of 13.1-16.0 g dL-1 (1.3%) (Table 4).
Packed cell volume:
Normal range (32.0-42.0%)
Subjects with 11-20% PCV had the highest prevalence of (55.1%), followed by PCV of 21-30%, with a prevalence of 24.4%, PCV of 31-40% recorded 19.2% prevalence (Table 5). The least was observed in PCV of 41-50%, with a prevalence of (1.3%).
DISCUSSION
The result of this study indicates that there is active transmission of malaria in the study area. The high prevalence observed might be attributed to the period of study (July to September) which is the period of maximum rainfall in Nigeria. In Nigeria high breeding rate of the vector and high transmission rate occur throughout the year especially during the rainy season (Salako, 1986; WHO, 1995; WHO, 2000). This is consistent with the report of Oparaocha (2003), who obtained 88.8% in his study, Aribodor et al. (2003), recorded a prevalence of 76% in their investigation, while Mbanugo and Ejim (2000), had 85% prevalence rate. Apart from increased number of breeding sites occasioned by incessant rains, other obvious factors like ignorance, poverty, unsanitary environmental conditions, poor behavioural attitudes and inadequately planned socio-economic projects might have contributed to increase transmission (Ejezie, 1983; FMOH, 1989). In all the children observed there was no significant difference between prevalence and sex. Similar result has been documented by Agbonlahor et al. (1993).
There was a significant association between malaria infection and age. The low prevalence observed in 1-5 months of age group is due to persisting maternally derived antibodies that are persistent in each child’s serum from the time of birth (Ebrahim, 1978). The incidence of malaria increases as the age increases, it is highest in the first 2 years of life when maximum morbidity and mortality occur. The markedly increased level of parasitaemia in the 1-5 years age group could be attributed to the gradual loss of these maternally derived antibodies and the development of acquired immunity (Irene et al., 1981; Luzzatto et al., 1969).
The blood group distributions found in this study are similar to those previously reported for the same Nigerian population by Kazim and Ejezie (1982), Worlledge et al. (1974), Opara et al. (2004). Blood group O was the most susceptible to malaria infection than the other blood groups. It has been reported that Anopheles gambie recognizes ABO blood group variation with a preference for blood group O (Wood and Harrison 1972). It is possible that this observation might have accounted for the high percentage of blood group O infected with malaria. The preponderance of blood group O in the population may also be a factor for the high prevalence (Opara et al., 2004). There was a significance association between ABO and malaria infection. This is consistent with the report of Agbonlahor et al. (1993), but contrasts with that of Facer and Brown (1979), Martins et al. (1979) and Kazim and Ejezie (1982). The association observed in this study is striking, if proved in subsequent studies, the association could be exploited in the production of potent drugs and/or vaccines against this deadly disease. We found that the degree of parasitaemia was less among children with Hb AS and Hb SS than among those with Hb AA. According to Luzatto (1979), the presence of sickle haemoglobin (Hb-S) in red cells limits the development and multiplication of the parasite. Population, field and clinical studies have provided convincing evidence that where P. falciparum is endemic, non-immune subjects who have only normal haemoglobin (Hb-AS) have a survival advantage over subjects, who have only normal haemoglobin (Hb-AA) (Fleming et al., 1985). This result agrees with our earlier report by Opara et al. (2004).
In malaria endemic regions of Africa, community surveys have shown the prevalence of anaemia in children to be between 49 and 89% (Crawley, 2001). It has been reported that severe malarial anaemia causes 190,000-974,000 deaths annually in children under the age of 5 years with the highest mortality observed among infants below the age of 1 year. There is a direct link between malaria clinical manifestation and haemoglobin levels (Oparaocha, 2003). Plasmodium infection, being a blood parasitic disease illicits marked invasiveness and destruction of the red blood cells which results into severe anaemia particularly in children (Mbanugo and Ejims, 2000). We assessed the degree of anaemia by investigating the effect of malaria on the mean haemoglobin and packed cell volume of the study population. Children with malaria parasitaemia had a significantly lower haemoglobin (<10 g dL-1) and packed cell volume (<30%) than children without parasitaemia. Anaemia defined as a haemoglobin of 11 g dL-1 or less occurred in 85% of the children. Bradley et al. (1985) obtained anaemia in 87% of the children examined.
In view of the high prevalence of malaria and anaemia observed in this study, there is need for promotion of better environmental conditions, such as better town planning and good housing with appropriate and well maintained drainage system, proper waste disposal management, proper nutrition and drastic change in our socio-cultural behaviours. The campaigns on the use and benefits of Insecticide Treated Nets (ITNs) should be intensified. All these are necessary for reduction of morbidity and mortality caused by plasmodium infections.
Finally, the data generated in this study would serve as a baseline data for future studies on the susceptibility of genetic indices to malaria infections in other parts of Nigeria.
REFERENCES
- Aribodor, D.N., O.O. Njoku, C.I. Eneanya and I.O. Onyali, 2003. Studies on prevalence of malaria and management practices of the Azia community, Ihiala L.G.A., Anambra State, South-East Nigeria. Niger. J. Parasitol., 24: 33-38.
Direct Link - Binka, F.N. and P. Adongo, 1997. Acceptability and use of insecticide impregnated bed nets in Northern Ghana. Trop. Med. Int. Health, 2: 499-507.
Direct Link - Bradley, Moore, A.M., B.M. Greenwood, A.K. Bradley and A. Akintunde et al., 1985. Malaria chemoprophylaxis with chloroquine in young Nigerian children. Ann. Trop. Med Parast., 79: 585-595.
Direct Link - Brewster, D.R., D. Kwarkowski and N.J. White, 1990. Neurological sequelae of cerebral malaria in children. Lancet, 336: 1039-1043.
Direct Link - Ejezie, G.C., 1983. The Nigerian environment and parasitic infection. Folia Parast., 30: 89-95.
Direct Link - Fleming, A.F., A. Akintunde, E.D. Attai, A.M. Bradley-Moore and B.M. Greenwood et al., 1985. Malaria and haemoglobin genotype in young Northern Nigerian children. Ann. Trop. Med. Parasitol., 79: 1-5.
PubMed - Irene, G.H., B. Ulrich and L. Luzzato, 1981. Plasmodium falciparum malaria and human red cells II red cell genetics trait and resistance against malaria. Int. J. Epidemol., 10: 16-22.
Direct Link - Jungery, M., 1985. Studies on the biochemical basis of the interaction of the merozoites of Plasmodium falciparum and the human red cell. Trans. Roy. Soc. Trop. Med. Hyg., 79: 591-597.
Direct Link - Kazim, O.O. and G.C. Ejezie, 1982. ABO blood groups in malaria and schistosomiasis haematobium. Acta Tropica, 39: 179-184.
PubMedDirect Link - Lackritz, E.M., C.C. Gamphell, T.R. Ruebus, A.W. Hightower and W. Wakube et al., 1992. Effect of blood transfusion on survival among children in a Kenya hospital. Lancet, 340: 524-558.
Direct Link - Luzatto, L., 1979. Genetics of red cells and susceptibility to malaria. Blood, 54: 961-976.
Direct Link - Luzzatto, L., E.A. Usanga and S. Reddy, 1969. Glucose-6-phosphate dehydrogenase deficient red cells: Resistance to infection by malaria parasite. Science, 164: 839-842.
CrossRef - Martins, S.K., L.H. Miller, C.U. Hicks, A. David-Wost, C. Ugbode and M. Deane, 1979. Frequency of blood group antigens in Nigeria children with falciparum malaria. Trans. Roy. Soc. Trop. Med. Hyg., 73: 216-218.
Direct Link - Miller, L.H., S.A. Dvorak, T. Shiroshi and J. Durocher, 1973. Influence of erythrocyte membrane component on malaria merozoite invasion. J. Exp. Med., 138: 1597-1601.
Direct Link - Miller, L.H., S.J. Mason, D.F. Clycle and M.H. McCinnis, 1976. The resistance factor of Plasmodium vivax in blacks the Duff, blood group FyFy. New Eng. J. Med., 295: 302-304.
Direct Link - Newton, C.R.J.C., P.A. Warn, P.A. Winstanley, N. Peshu, R.W. Snow, G. Pasvol and K. Marsh, 1997. Severe anaemia in children living in a malaria endemic area of Kenya. Trop. Med. Int. Health, 2: 165-178.
CrossRefPubMedDirect Link - Opara, K.N., E.S. Ibanga, N.B. Wali and L.P. Usip, 2004. Falciparum malaria and their susceptibility to genetic markers of pregnant women in Uyo, South Eastern Nigeria. J. Applied Zool. Environ. Biol., 6: 56-60.
Direct Link - Oparaocha, E.T., 2003. The impact of haemoglobin level and concomitant infections on malaria parasitaemia and on-set of fever during malaria attack in Ikwuano local government area of Abia State, Nigeria. Niger. J. Parasitol, 24: 25-32.
Direct Link