
Necda Eldeniz Cankaya
Black Sea Agricultural Research Institute, Samsun, Turkey
Osman Kaftanoglu
Department of Animal Science, Faculty of Agriculture,
cukurova University, Adana, Turkey
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1282-1286







ABSTRACT
The mated bumblebee queens overwinter in the soil and they make nests under the ground they come into contact with many ground dwelling pathogens and parasites. These pathogens and parasites may kill the queen. This study was conducted to investigated the occur some diseases and parasites on 578 mated Bombus terrestris L. queens collected from Adana, Mersin, Antalya and Bodrum regions in Turkey. For Nosema bombi, the highest infestation ratios were found in Bodrum ecotype (17.48%), for Apicystis bombi, the highest infestation ratios were found in Bodrum ecotype (8.54%), for Crithidia bombi, the highest infestation ratios were found in Bodrum ecotype (9.76%), for bacteria, the highest infestation ratios were found in Mersin ecotype (39.09%), for fungus, the highest infestation ratios were found in Mersin ecotype (17.27%), for internal parasites (Locustacarus buchneri), the highest infestation ratios were found in Bodrum ecotype (13.01%) and for external mites (Kutzinia laevis, Scutacarus acarorus, Parasitus fucorum) the highest infestation ratios were found in Antalya ecotype (41.44%, respectively).
PDF Abstract XML References Citation
How to cite this article
Necda Eldeniz Cankaya and Osman Kaftanoglu, 2006. An Investigation on Some Diseases and Parasites of Bumblebee Queens (Bombus terrestris L.) in Turkey. Pakistan Journal of Biological Sciences, 9: 1282-1286.
DOI: 10.3923/pjbs.2006.1282.1286
URL: https://scialert.net/abstract/?doi=pjbs.2006.1282.1286
DOI: 10.3923/pjbs.2006.1282.1286
URL: https://scialert.net/abstract/?doi=pjbs.2006.1282.1286
INTRODUCTION
Bumblebee breeding and using them in greenhouses especially during the off seasons has become an essential part of the modern agriculture. With the developments on the breeding techniques they are being widely used as pollinators in the greenhouses to increase the yield and the quality of the vegetables. Thousands of colonies are reared in the laboratories and they are being shipped to different countries every year for the last ten years. The health of the colonies or the introduction of unwanted diseases or parasites was one of the major concerns for the importing countries.
The natural bumblebee queen and colonies suffer from many pathogens (viruses, bacteria, fungi, protozoa) internal and external parasites. Heavily infested queens can not establish colonies. Catching naturally mated and hibernated queens in the spring and fall and bringing them to the breeding stations may cause the introduction of the pathogens and parasites into the facilities.
Moreover the commercial colonies may suffer from the stress during transportation and quarantine and develop some diseases if the pathogens exist in the colonies. Infected colonies may not show any symptoms before transportation since the brood area is covered with cotton layer in the nest box and they may enter other countries without being noticed. Therefore the identification and control of bumblebee diseases and parasites are very important for preventing the spread of diseases and parasites, as well as the rapid growth of the colonies and pollination efficiency. Diseased colonies can not develop as fast and they will have smaller number of bees than the health colonies.
There are very few researches on the distribution, symptoms and control of bumblebee diseases and internal enemies compared to that of honeybees. The most common bumblebee diseases and parasites can be summarized as follows.
MATERIALS AND METHODS
A total of 578 bumblebee queens were collected from Adana, Mersin, Antalya and Bodrum regions in Turkey in order to investigate the occurrence of pathogenic diseases and parasites. The queens were caught by using insect net during October and January. They were placed in plastic queen cages containing queen candy which was made by mixing powdered sugar and honey.
The queens were placed in breeding boxes and then temperature and humidity of breeding room were kept 28°C and 60-65%, respectively. This temperature and the humidity are also suitable for the growth of many pathogenic viruses, bacteria, fungi and protozoa. The queens were fed up with sugar syrup (1:1) and fresh pollen one bumblebee pupa and/or bumblebee workers were placed in the boxes in order to stimulate oviposition. The queens were checked for the diseases and parasites within one month if they did not statu oviposition. These queens were anesthetized with CO2 and killed in deepfreeze. Then they were dissected under a stereo-zoom microscope after they checked external mites. The second tergum was separated from abdomen to expose the air sacs. The air sacs and trachea were checked for the determination of the internal mite, which is Locustacarus buchneri, parasites under a stereo zoom microscope. Their abdomen checked for fungus and searched for bacteria, its midgut, malphigian tubes and fat body searched for Nosema bombi, Apicystis bombi and Crithidia bombi spores. A piece of midgut were placed on a microscope slide and the pathogens were observed at 400 magnification and under the compound microscope. For the sure Nosema bombi, Apicystis bombi and Crithidia bombi. We had paint Örösipal methods for sure Nosema bombi spores or yeast.
RESULTS
Protozoa: There are three sporozoan species such as Nosema bombi, Apicystis bombi and Crithidia bombi that affect the bumblebees.
N. bombi spores (Fig. 1) are 4.6-7.0 μm long and 1.8-4.0 μm wide and longer and wider than Nosema apis (Weiser, 1978; Mcivor, 1990; Eijnde and Vette, 1993; Macfarlane et al., 1995). It is a serious bumblebee disease. It distends and paralyses the abdomen, causes diarrhea and inhibits the mating in bumblebees (Macfarlane et al., 1995). Heavily infected queens can not hibernate and die. Mildly infected queens can survive during hibernation. However it delays the colony initiation and inhibits the growth of the colonies. Infected queens may die sooner during the colony development than the healthy queens.
A. bombi has a sausage shaped spores which are 11.1-14.4 μm long and 3.6-5.4 μm wide (Fig. 2). They may be boomerang shaped and they are about 2-3 times longer and wider than the N. bombi (Lipa and Triggiani, 1992, 1996; Macfarlane et al., 1995). A. bombi is usually found in the fat bodies. Queens heavily infected with A. bombi has white fat bodies, which can be easily detected. The fat bodies of the normal queens are yellow color and they darken and become brown as queens age. The pathogen shortens the life span of the queens and causes mortality in early spring.
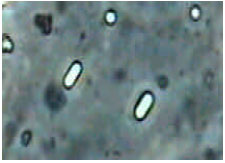 | |
| Fig. 1: | Nosema bombi spores (original) |
 | |
| Fig. 2: | Apicystis bombi spores (original) |
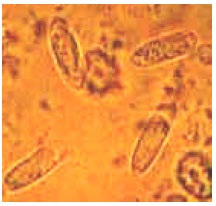
Crithidia bombi (Fig. 3) is also very common in the intestine and in its host of the B. terrestris. The flagellate inhabits the gut and it had the maximum length 8.1 μm, their bodies are pear-like in shape. The flagellum is 8-12 μm long (Lipa and Triggiani, 1988). The infection levels can reach up to 54-81% in queens, 75-80% in workers and 47-71% in males however their effects on the colony development is not known (Skou et al., 1963; Shykoff and Schmid-Hempel, 1991a, b).
Fungi: There are several fungi isolated from the bumblebees and their nests. The Paesilomyces farinosus, Beauveria bassiana, Verticillium lecanii, Chrysossporium pannorum, Metarhizium anisopliae, Aspergillus candidus, Doratomyces putredinis and Hirsutella sp. are the known species of fungi associated with bumblebees. The fungal contamination and inadequate food source are considered to be the major causes for the mortality during and/or after hibernation in early spring. Fungal contamination also causes the mortality of the brood during the colony development (Macfarlane et al., 1995).
Bacteria: Bacterial contamination also cause queen and brood mortality in bumblebee colonies. The midgut of the recently dead queens were found to be dark and contain numerous bacteria.
 | |
| Fig. 3: | Crithidia bombi spore (original) |
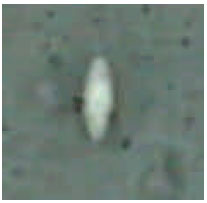
 | |
| Fig. 4: | Gravid female of Locustacarus buchneri (original) |
Some of the dead queens started smelling like rotten meat within 1-2 days. These queens lost the appendages like wings, legs and abdomens during inspection. After microscopic inspections and bioassay studies they showed the typical symptoms of Pseudomonas apiseptica which cause septicemia in honeybees.
Internal mites: There is only one well-known species of internal mite Locustacarus buchneri parasitizing different species of bumblebees. L. buchneri lives in the air sacs of the first abdominal segments of the adult bumblebees. Females mites overwinter inside the hibernating queens. They pierce the tracheal wall of the host by their mouthparts and feed on haemolymph. When the queen bee becomes active in spring the females develop ovaries and lay eggs (Husband and Sinha, 1970). The gravid females are yellow in color and round in shape (Fig. 4). They are about 0.7 mm long and difficult to see with the naked eye. They can be easily observed with the stereoscope microscope. They lay about 30-40 eggs which give rise to 6 legged male and female larviforms. The male larviforms (Fig. 5) are 0.15 mm long and 0.1 mm wide and transparent colored. The female larviforms (Fig. 6) are 0.4 mm long and 0.2 mm wide.
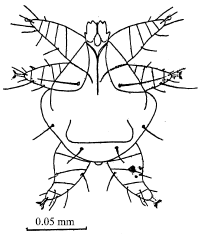 | |
| Fig. 5: | Larviform male of L. buchneri * |
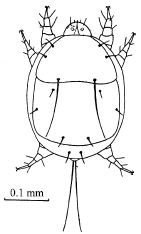 | |
| Fig. 6: | Larviform female of L. buchneri.* (*: from Husband and Sinha, 1970) |
The female mite can deposit up to 50-60 eggs in a cluster. Sometimes several gravid female mites and 5-6 egg clusters can be found in single queen. They also deposit sticky brownish feces in the air sacs of the host. The male and female mites mate in the air sacs and mated female parasites leave the host through the host`s spiracles and infest other bees in the nest. They enter the new hosts through the abdominal spiracles.
External mites: There are also some external mites found on bumblebees or in the nest. The most common external bumblebee mites are Kutzinia laevis (Fig. 7), Parasitus fucorum, Scutacarus acarorum (Fig. 8), Hypoaspis sp. Garmaniella sp. Fuscuropoda and Glycyphagus sp. Most of the external mites are phoretic on bumblebees, feed on pollen and nest debris and not considered to be major parasites in the natural environments (Macfarlane et al., 1995).
The occurrence and the infection ratios of bumblebee (B. terrestris) diseases and parasites in four different regions in Turkey are summarized in Table 1.
| Table 1: | The occurrence of diseases and parasites in different ecotypes of B. terrestris |
 | |
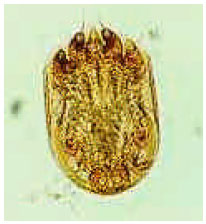 | |
| Fig. 7: | Kutzinia laevis (original) |
 | |
| Fig. 8: | Scutacarus acarorum (original) |
The average Nosema bombi infection rate was found to be 12.96% among the four ecotypes of Bombus terrestris. The infection ratio was the highest in Bodrum ecotype and the lowest in Antalya ecotype (p<0.01). There was no significant difference between the Adana and Mersin ecotypes (p<0.05). N. bombi infection was reported to be between 7-43% in New Zealand, 13.5% in Switzerland and 14% in Italy in B. terrestris queens (Fisher and Pomeroy, 1989; Mcivor, 1990; Triggiani, 1991; Shykoff and Shmid-Hempel, 1991a). The infestation ratio of N. bombi in Turkey is almost the same as in Switzerland and in Italy.
The average Apicystis bombi infection rate was found to be 6.89% among the four ecotypes of Bombus terrestris. The infection ratio was the highest in Bodrum ecotype and the lowest in Antalya and Adana ecotypes (p<0.01). They reported are A. bombi infection was found to be North America and Europa and was not found in New Zealand and Japan (Liu et al., 1974; Lipa and Triggiani; 1992; Macfarlane et al., 1995). The infestation ratio of Apicystis bombi in Turkey is higher than North America (Canada, 2-2.5%) and lower than in Europa (6-16%).
The average infection ratio of Crithidia bombi was found to be 8.99% in Anatolian B. terrestris. Infection ratio was found to be lower in Antalya ecotype and the higher in Bodrum ecotype (p<0.05). There was not a significant difference among the Adana and Mersin ecotypes (p<0.05). The C. bombi infection was reported to be between 54-81% in queens, 75-80% in workers and 47-71% in males in Switzerland (Shykoff and Shmid-Hempel, 1991a, b; Skou et al., 1963).
The average infestation ratio of the internal parasite Locustacarus buchneri was found to be 5.28% among the ecotypes. The infestation ratio was lowest in the Mersin ecotype and highest in the Bodrum ecotype. Antalya ecotype also has higher infestation compared to Adana and Mersin ecotypes (p< 0.05). L. buchneri infestation ranged between 20-24% in B. vagans, B. perplexus and B. bimaculatus in the USA (Goldblatt and Fell, 1984) whereas 2.7% in B. bimaculatus and 1.7% in B. perplexus in Ontario Canada (Liu, 1973; Wyatt, 1970). The infestation level was found to be 0.8% in B. terrestris and 7.0% in B. hortorum in Europe. It was seen that the Anatolian B. terrestris had much higher level of L. buchneri infestation than the Europeans.
There was a significant difference concerning the infestation of the external mites among the ecotypes. The ratio of infested queen bees were 41.44% for the Antalya ecotype, 34.55% for the Mersin, 19.11% for the Bodrum and 17.12% the Adana ecotype. The Antalya ecotype had the highest and the Adana ecotype had the lowest external mites (p<0.01).
The fungi infestation was significantly different among the ecotypes (p<0.01). It was found that 8.11, 17.27, 9.01% and 6.10% of the Adana, Mersin, Antalya and Bodrum ecotypes, respectively. The infestation ratio was the highest in the Mersin ecotype and the lowest in the Bodrum ecotype.
Almost all the ecotypes had high bacterial infection. Some of the dead queens had septicemia type symptoms and probably died from Pseudomonas apiseptica which also causes septicemia in honey bees.
DISCUSSION
Bumblebee diseases and parasites are very important for the successful breeding of the colonies. Rearing efforts will be successful if healthy queens are selected for breeding. Some antibiotics such as fumagillin can be used to prevent N. bombi infections in the rearing facilities. Different antibiotics can be used to prevent bacterial infections however uses of antibiotics may increase fungal growth in the colonies. Any kind of contamination with pathogens and internal parasites will shorten the colony life and decrease the pollination efficiency of the colonies.
Uses of bumblebee colonies are increasing throughout the world. Thousands of colonies are exported to different countries every year. Some of the countries do not have all these diseases and parasites. Introduction of the diseases and parasites will cause serious problems for the importing countries. Therefore bumblebee breeding companies should be very careful about selecting healthy stocks, breeding healthy colonies and not shipping diseased colonies to other countries.
ACKNOWLEDGMENTS
Thanks to the NATO SFS Programme, TUBITAK and The Cukurova University Research Funds for supporting the project. We also thanks to Dr. Hayo W.W. Velthuis, Dr. M.H. Duchateau, from the Utrecht University; Dr. A. de Ruijter and Institute personnels from Insect Pollination and Beekeeping Research Institute in Hilvarenbeek in Netherlands for technical support.
REFERENCES
- Fisher, R.M. and N. Pomeroy, 1989. Pollination of greenhouse muskmelons by bumble bees. J. Econ. Entomol. Entomol. Soc. Am., 82: 1061-1066.
Direct Link - Goldblatt, J.W. and R.D. Fell, 1984. Parasites and parasitization rates in bumble bee queens, Bombus, sp. (Hymenoptera:Apidae), in Southwestern Virginia. Environ. Entomol., 13: 1661-1665.
Direct Link - Husband, R.W. and R.N. Sinha, 1970. A revision of the genus Locustacarus with a key to genera of the family podapolipidae (Acarina). Ann. Entomol. Soc. Am., 63: 1152-1162.
Direct Link - Lipa, J.J. and O. Triggiani, 1988. Crithidia bombi a flagellated parasite of a bumble-bee (Bombus terrestris L.) (Hymenoptera, Apidae). Acta Protozool., 27: 287-290.
Direct Link - Lipa, J.J. and O. Triggiani, 1992. A Newly recorded neogregarine (Protozoa, Apicomplexa), parasite in honey bees (Apis mellifera) and bumble bees (Bombus sp.). Apidologie, 23: 533-536.
CrossRef - Lipa, J.J. and O. Triggiani, 1996. Apicystis gen nov and Apicystis bombi (Liu, Macfarlane and Pengelly) comb nov (Protozoa: Neogregarinida), a cosmopolitan parasite of Bombus and Apis (Hymenoptera: Apidae). Apidologie, 27: 29-34.
CrossRef - Liu, H.J., R.P. Macfarlane and D.H. Pengelly, 1974. Mattesia bombi (Neogregarinida: Ophrocystidae), Parasite of Bombus (Hymenoptera: Apidae). J. Invertebr. Pathol., 23: 225-231.
PubMed - Shykoff, J.A. and P. Schmid-Hempel, 1991. Incidence and effects of four parasites in natural populations of bumble bees in Switzerland. Apidologie, 22: 117-125.
Direct Link