
Audifax N. Iwu
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
C.S. Ebere
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
I.P. Ogbuewu
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
I.F. Etuk
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
M.N. Opara
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
M.C. Uchegbu
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
I.C. Okoli
Department of Animal Science and Technology, Federal University of Technology, Owerri, Imo State, Nigeria
F.C. Iheukwumere
Department of Animal Science, Abia State University, Umuahia, Abia State, Nigeria
U. Herbert
College of Animal Science, Michael Okpara Federal University of Agriculture, Umudike, Abia State, Nigeria
Journal of Medical Sciences
Year: 2013 | Volume: 13 | Issue: 7 | Page No.: 563-570







ABSTRACT
Forty eight crossbred male and female rabbits aged 12-14 weeks were used to study the effects of supplementing Coconut Shell Ash (CSA) on reproductive organ characteristics and serum sex hormone concentrations of growing rabbits. The rabbits were divided into 24 males and females, respectively. Each group was divided into four experimental treatments T1, T2, T3 and T4 of six animals and was further replicated three times with two animals per replicate in a Completely Randomized Design (CRD). The animals were fed a commercial grower rabbit ration which was supplemented with CSA at the rate of 0, 1, 2 and 3 g kg-1 b.wt., respectively. At the end of 12 weeks feeding, three rabbits were selected from each group, slaughtered and used to determine reproductive organ characteristics. Blood samples from the slaughtered animals were collected from the jugular vein and assayed for serum reproductive hormones concentrations. Data collected were subjected to analysis of variance. Testicular morphometry result showed that 1 to 2 g kg-1 b.wt. CSA supplementation could improve the weight of testis, size and epididymal weights indicating that up to 2 g kg-1 b.wt. CSA supplementation supported superior testicular development. This was associated with increasing serum testosterone concentration indicating probably a CSA enhancement of metabolic activity. One gram kg-1 b.wt. was the optimal CSA supplementation rate in female rabbits, since this led to improved weight of whole reproductive tract, weight of ovary, oviduct and uterus. This was equally associated with increased estrogen concentration, which increased with increasing CSA supplementation.
PDF Abstract XML References Citation
Received: March 26, 2013;
Accepted: May 04, 2013;
Published: July 04, 2013
How to cite this article
Audifax N. Iwu, C.S. Ebere, I.P. Ogbuewu, I.F. Etuk, M.N. Opara, M.C. Uchegbu, I.C. Okoli, F.C. Iheukwumere and U. Herbert, 2013. Coconut Shell Ash as a Mild Agonist of Reproductive Organ Development and Sex Hormone Release in Growing Rabbits. Journal of Medical Sciences, 13: 563-570.
DOI: 10.3923/jms.2013.563.570
URL: https://scialert.net/abstract/?doi=jms.2013.563.570
DOI: 10.3923/jms.2013.563.570
URL: https://scialert.net/abstract/?doi=jms.2013.563.570
INTRODUCTION
The domestic rabbit is a herbivorous animal that consumes grains, forages and hay to derive its essential nutrients. Its diets, whether home made or commercially prepared consist mostly of ingredients from plant sources (Herbert, 2001; Nworgu et al., 2001; Biobaku and Dosumu, 2003; Herbert et al., 2005; Omoikhoje et al., 2006). Although minerals are nutrients required in small amounts by animals, they are however more essential to body metabolism than many other food nutrients. For example, essential trace minerals are important constituents of metalloenzymes, which are important in bone formation, lipid metabolism, glucose utilization, iron transport and free radical metabolism (Leach, 1967; Jovanoic-Peterson and Peterson, 1996; Cuanne et al., 1993; Hostetler et al., 2003; Townsend et al., 1994). Short (1980) reported that minerals are particularly essential for growing rabbits, therefore without adequate supply, production will certainly be lowered.
In animal production, natural clays are inexpensive natural minerals, which have protective effect against diarrheic microorganisms and conditions (Ayyat et al., 1995). Watanable et al. (1971) and Grobner et al. (1982) reported that when zeolite is included in rabbits ration, it helps in reducing incidence of entritis. Alternative mineral sources are however receiving greater attention in animal and human nutrition. Earlier research efforts have employed natural clay properties, such as ion exchange, adsorption and binding properties in enhancement of mineral uptake from the gastro intestinal tract (Ayyat, 2010). Shuerson et al. (2011) however reported that in contrast to the use of inorganic sources of minerals, the use of chelated minerals will prevent degradation of feed vitamins and also improve bioavailability of essential trace elements.
The nutritional requirements for minerals elements are small; however, these nutrients can greatly affect reproduction (Hostetler et al., 2003). The potential of minerals to play significant roles in mammalian fertility and general reproductive wellbeing have been reported. According to Hostetler et al. (2003), minerals categories of particular importance in this regard are the macro elements (Ca, P, K, Na and Mg) and the trace elements (Cu, Co, Se Mn, I and Zn). Thus, physiological concentrations of the elements must always be adequate to ensure proper cellular and reproductive functions in animals (Boland, 2003). Specifically, mineral deficiencies and imbalances were cited by Corah and Ives (1991) and Boland (2003) as important causes of poor reproductive functions; particularly ovarian activity of ruminants is influenced by mineral deficiency.
Minerals also play vital roles in the endocrine system and tissues integrity, therefore making their optimal availability critical to follicular development and fertility in animals. That reproductive failure may be induced by deficiency of single or combined elements and by their imbalances was particularly highlighted in the reports of Graham (1991) and Maas (1993). However, much remains to be understood about the effects of mineral deficiencies and imbalances on development and sustenance of reproductive structures and functions in animals.
Several cultural reports have highlighted the value of highly alkaline plant ash in the improvement of the nutritional value of indigenous foods (Kyarisiima et al., 2004; Hayes, 2009; Webber, 2011). Ochetim (1988) suggested the use of wood ash as a source of minerals for poultry, while Oso et al. (2011) reported occurrence of lameness and poor gaits in broilers fed diet containing wood ash as Calcium source and therefore concluded that wood ash inclusion in broiler diets should be discouraged since it results in poor production results. However, recent studies by Iwu (2012) and Ebere (2012) showed that supplementation with coconut shell ash at the level of 1 g kg-1 b.wt. was optimal for growing rabbits fed commercial diet, while higher levels of supplementation resulted in some mildly inferior serum biochemical responses.
After the combustion of wood or any plant part, the inorganic and organic residue remaining is termed wood ash. The physical and chemical properties of wood ash have been reported to vary significantly across many factors, such as tree species, soil type, climate and level of material combustion (Greene, 1988; Iwu et al., 2012; Nwogu et al., 2012). Temperate-climate wood was reported by Campbell (1990), to yields 0.1-1.0% ash, while tropical and subtropical wood yields up 5%. The average ash yield for most woods however is about 6-10%.
Ash has a small particle size of 230 μm (Etiegni and Campbell, 1991). Completely ashed (total elimination of carbon) coconut shell yielded an ash with a bulk density of 0.6512 g cm-3, average water holding capacity of 210.54 g mL-1 and specific gravity of 0.65 g cm-3 (Iwu et al., 2012). Ash density generally varies with carbon content in the form of organic matter.
This study was designed to determine the effects of coconut ash supplementations on the reproductive organs characteristics and hormonal profiles of male and female growing rabbits.
MATERIALS AND METHODS
Test materials: Coconut shell was collected from different markets and households within Owerri zone of Imo State, Nigeria. The coconut ash (CSA) was prepared a customize ashing technique was developed (Iwu et al., 2012). Briefly, the customized ashing technique required an overnight initial ashing of the coconut shells in an industrial bread oven and a subsequent re-ashing in an open clay pot on a kerosene stove till the ash glowed red hot. This process eliminated all carbon components of the ash, thus concentrating the minerals (Iwu et al., 2012). The ash was collected and preserved in a sterile container for subsequent use.
Experimental animals: The animals used in the research were 48 New ZealandxChinchilla cross bred pre-pubertal male and female rabbits, aged 3 months. The animals were housed 2 rabbits per cage in standard wooden/wire mesh cages. Two weeks pre-treatment period was used to acclimatize the animals with their new environment and to get them used to the experimental procedure.
Experimental design: The 48 rabbits were divided into two major groups of 24 males and 24 females. These were each further divided into four treatment groups coded T1, T2, T3 and T4 of six rabbits each, which were then replicated 3 times with made 2 rabbits per replicate in a Completely Randomized Design (CRD).
Experimental feed: The experimental diet used in this study was a commercial mash purchased from a reputable dealer. The nutrient content of the feed was compared (Table 1) and found to be similar to the needs of growing rabbits (Taiwo et al., 2005).
Again, the experimental ration was analyzed for its mineral content with the aid of Atomic Absorption Spectrometer (Bulk Scientific, 205) to determine its concentrations of essential mineral elements.
Feeding experiment: Each male and female treatment group received a determined quantity of the CSA as shown below:
| • | T1 was the control and has 0 g CSA supplementation |
| • | T2 was on 1 g CSA kg-1 b.wt. |
| • | T3 was on 2 g CSA kg-1 b.wt. |
| • | T4 was on 3 g CSA kg-1 b.wt. |
| Table 1: | Composition of experimental diets |
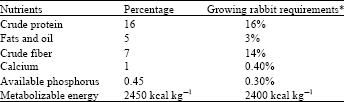 | |
| *Source: Taiwo et al. (2005) | |
The test CSA was offered to the animals by feed supplementation. A small amount of feed was used to mix the CSA and offered to the animals. After the treated feed has been consumed the remaining feed for the day was offered. Feed and water were offered ad libitum and all standard management practices as practiced in the Federal University of Technology Owerri Teaching and Research Farm were observed. The feeding experiment lasted for 12 weeks.
Determination of reproductive organs characteristics: At the end of the 12 weeks feeding experiment, one animal from each of the replicates, making three per treatment group were starved overnight, weighed and slaughtered and thereafter dressed and dissected following standard methods. The reproductive organs were harvested, trimmed free of fats and adhering connective tissues. In the male, the weight of the whole organ was recorded after which the testes was carefully removed from its sac and weighed. The epididymis was also weighed. The weight of each testis was measured and recorded after the epididymis had been trimmed off. The epididymis was weighed after removing it from each testis. All weighing was done using highly sensitive digital balance (Adventurer RS 232).
Similarly, in the female, the whole reproductive tract was separated from the carcass and trimmed free of adhering fats and connecting tissues. The weight of the whole tract was measured and recorded, after which the ovary was carefully removed from the ovarian bursa at the end of the infundibulum and weighed. Again, the infundibulum was removed, followed by the oviduct and weighed, while the uterine horn was removed from the end of the two cervixes to the rosette projection of the utero-tubal junction and also weighed.
Blood collection: Blood was collected from their throat after severing the carotid artery and jugular vein. The rabbit was first removed from the cage by holding it securely on the scuff, held by the hind and fore limbs and the neck from which the blood was to be collected with the neck held downward. The blood was then collected immediately into a set of sterile plastic bottles without anti-coagulant for serum biochemical tests.
Determination reproductive hormones: The blood samples were allowed to coagulate to produce the sera for the hormonal analyses. They were thereafter, taken to the laboratory within 3 h of their collection for analyses. The serum samples obtained were analyzed to determine the concentrations of testosterone and luteinizing in males and leuternizing hormone, follicle stimulating hormone, prolactin, progesterone and estrogen in the females. The analysis was carried out with the aid of the tube-based enzyme immunoassay (EIA) method. The protocol used for the hormone was according to the method of Micaleft et al. (1995) as described for the kit (BioCheck ELISA Assay, USA).
Data analysis: Data collected on the different parameters was subjected to Analysis of Variance (ANOVA) (Steel and Torrie, 1980). The treatment mean values were tested for significant differences by Duncan’s New Multiple Range Test (SAS, 2000).
RESULTS AND DISCUSSION
The mineral content of the experimental diet and the recommended standard values by Standard Organization of Nigeria (Standards Organization of Nigeria, 2003) were presented in Table 2. The mineral content of the commercial diet was very low for all the elements considered when compared with the mineral requirements stipulated by Standard Organization of Nigeria for such commercial feeds. This clearly indicates the need for mineral supplementation when using such feeds. The present result agrees with that of Okoli et al. (2012) that commercial feeds produced in Nigeria are significantly deficient in their mineral contents. Again, the values obtained especially for sodium and magnesium were below, the requirements for growing rabbits (Lebas et al., 1986).
| Table 2: | Mineral content analysis of the commercial grower ration |
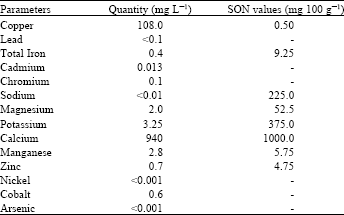 | |
Testicular morphometric characteristics: The data on testicular morphometric characteristics of buck rabbits supplemented varying levels of coconut shell ash were presented in Table 3. The testicular weight of T2 bucks was generally higher than those of the other groups and was specifically superior to those of T3 and T4 (p<0.05). T2 recorded significantly higher right, left and paired values (p<0.05) than T3 and T4 but similar to the control.
From these results, increased supplementation of coconut shell ash beyond 2 g kg-1 b.wt. tended to cause a decrease in the testicular size of the bucks. The left testis was observed to be relatively heavier when compared with right testis. However, these ware not significantly (p<0.05) different. The epididymal weight values of T3 on the other hand were generally higher than those of the other groups. The T3 values were specifically significantly higher than the T4 values (p<0.05). Again, as the coconut shell ash inclusion increased beyond 2 g kg-1 b.wt., the epididymis weights decreased.
The testicular weights have been found to provide valuable information in the evaluation of sperm production, breeding and fertility of the animals. It is specifically a good indicator of the present and future sperm production of an animal (Tohman and Massanyi, 1997; Morris et al., 1978; Perry and Petterson, 2001; Galmessa et al., 2003). Larger testes (without any abnormality have been reported to produce more sperm than smaller testes (Oyeyemi et al., 2002; Brito et al., 2004; Oyeyemi and Okediran, 2007). Morton (1988) reported that decreased weight of testes indicates widespread or diffuse lost of semiferous epithelia cells. The testes which possess greater number of sertoli cells were heavier and produced more sperm than testes with fewer sertoli cells (Berndtson et al., 1987).
Female organ morphometric characteristics: The results of female reproductive organ characteristics were presented in Table 4. The weights of the full reproductive organ were between the values of 7.07-8.35 g. T2-value was significantly higher than T1, T3 and T4.
Thus, the 1 g kg-1 b.wt. supplementation of CSA positively influenced the female reproductive organ characteristics indicating superior reproductive affinity in the group receiving this level of supplementation.
| Table 3: | Effect of coconut shell ash supplementation on testicular values of buck rabbits |
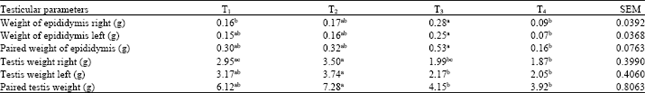 | |
| Means within a row with different superscripts differ significantly (p<0.05) | |
| Table 4: | Effect of coconut shell ash supplementation on female reproductive organ characteristics |
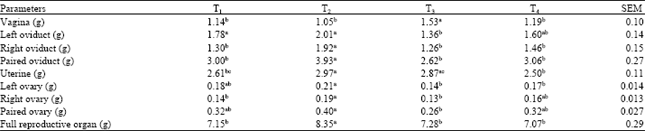 | |
| Means within a row with different superscripts differ significantly (p<0.05) | |
| Table 5: | Serum hormones characteristics of buck rabbits supplemented coconut shell ash (Cocos nucifera L.) diet |
| Means within a row with different superscripts differ significantly (p<0.05) | |
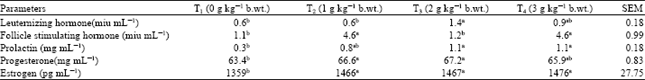
| Table 6: | Serum hormone concentrations of female rabbits fed varying levels of CSA |
 | |
| Means within a row with different superscripts differ significantly (p<0.05) | |
Specifically, the T2 values for right ovary, paired ovary and whole reproductive organ were significantly higher (p<0.05) than the values recorded for the other treatments. T2 values for left ovary was also significantly higher (p<0.05) than the values recorded for T3, while T3 on the other hand recorded significantly higher vaginal weight (p<0.05) than the rest. However, the ovarian weights recorded in this study compared favorably with the values of reported by Bitto (2007), Osuagwu (2004) and Ogbuewu (2008) in rabbits.
Male reproductive hormones profile: The effect of CSA supplementation on serum sex hormone concentrations of buck rabbits were presented in Table 5. Values for the two hormones assayed rose as a result of 1 g kg-1 b.wt. CSA supplementation but dropped at 2 g kg-1 b.wt. and subsequently increased again at 3 g kg-1 supplementation (p<0.05).
Serum sex hormone concentrations provide valuable information in rabbit breeding program. The increase in the values of some of these hormones indicates that CSA supplementation may be promoting hormone syntheses in the growing rabbits. The serum testosterone level were particularly above the normal range of 0.5-10 mg mL-1 (Castro et al., 2002), while the luteinizing hormone levels were below the normal range of 15-20 mg mL-1 reported by Moor and Younglai (1975). The increasing concentration of testosterone in the blood circulation plays a role in metabolic functions throughout the body by stimulating protein synthesis and muscle growth (Osinowo, 2006). According to Kerr and Sharpe (2006) the circulating LH in the male is responsible for maintaining normal plasma testosterone concentration.
Female reproductive hormones profile: The serum concentrations of the female hormones were presented in Table 6. The values of the LH were within the range 0.6 to 1.4 μ mL-1. At 2 g kg-1 b.wt. (T3) supplementation of CSA, the value of LH doubled, while subsequent increase in CSA supplementation caused decrease in values, which was however still higher that the control and T2 values.
Luteinizing hormone stimulates the secretion of sex steroids from the gonads (Osinowo, 2006). The reports of Gallo (1981) and Hashimoto et al. (1987) among several others have demonstrated that the LH surges at the pro-estrous stage are responsible for ovulation. Therefore, any substance capable of enhancing its release would improve ovulation of mature follicles (Benie et al., 2003; Ogbuewu, 2012).
The values of the Follicle Stimulating Hormones (FSH) were within the range of 1.1 to 4.6 μ mL-1. T2 and T4 groups recorded significantly higher (p<0.05) values of 4.6 μ mL-1 than T1 and T3. The values of the prolactin were within the ranges of 0.3 to 1.1 mg mL-1. However, increasing levels of CSA supplementation resulted in increased serum values that stabilized at 1.1 mg mL-1 in T3 and T4. FSH is essential for gonadal development and maturation at puberty and gamete production during the pubertal phases of development (Simoni and Nieschlag, 1995).
The serum concentrations of progesterone were within the range 63.4 to 67.2 mg mL-1. The values increased with CSA supplementation up to 2 g kg-1 b.wt. supplementation (67.2 mg mL-1) but declined slightly to 65.9 mg mL-1 in the T4 groups. The values of estrogen were also within the range of 1359 to 1476 pg mL-1 and increased with increasing values of CSA supplementation.
In non pregnant females undergoing normal reproductive cycle, the progesterone level remains relatively constant throughout the luteal phase of the cycle and then increases rapidly following the ovulation (Uboh et al., 2010). The increase in the concentration of blood estrogen with increasing supplementation of CSA in this study could therefore be attributed in part to increased metabolic activity in the higher supplemented animals, probably induced by increased mineral uptake and assimilation earlier observed in these animals (Iwu, 2012; Ebere, 2012). It is however important to note that the levels of these hormones are influenced by the cyclic stage of the female animals and stage of development, (pre-pubertal, pubertal and post pubertal).
CONCLUSION
The result of testicular morphometry showed that 1 to 2 g kg-1 b.wt. CSA supplementation could improve the weight of testis, size and epididymal weights indicating that up to 2 g kg-1 b.wt. CSA supplementation would support better testicular development of rabbits. This was associated with increasing serum testosterone concentration indicating probably a CSA inducement of improved metabolic activity.
The present study also showed that 1 g kg-1 b.wt. was the optimal CSA supplementation rate in female rabbits, since this led to improved weight of whole reproductive tract, weight of ovary, weight of oviduct and uterine weight. This was equally associated with increased estrogen concentration, which increased with increasing CSA supplementation.
REFERENCES
- Ayyat, M.S., I.F.M. Marai and A.M. Alazab, 1995. Copper-protein nutrition of New Zealand white rabbits under Egyptian conditions. World Rabbit Sci., 3: 113-118.
Direct Link - Benie, T., T. Duval and M.L. Thieulant, 2003. Effects of some traditional plant extracts on rat oestrous cycle compared with clomid. Phytotherapia Resour., 17: 748-755.
CrossRefDirect Link - Biobaku, W.O. and E.O. Dosumu, 2003. Effects of supplementing diets based maize and rice bran with three different improved forages on feed intake, digestibility and growth in rabbits. Proceedings of the Conference on Nigerian Society of Animal Production, Volume 22, March 17-21, 2003, Zaria, pp: 179-184.
- Boland, M.P., 2003. Trace minerals in production and reproduction in dairy cows. Adv. Dairy Technol., 15: 319-330.
Direct Link - Brito, L.F.C., A.E.D.F. Sliva, M.M. Unanian, M.A.N. Dode, R.T. Barbos and J.P. Kastelic, 2004. Sexual development in early and late maturing Bos indicus and Bos indicus x Bos taurus crossbred bulls in Brazil. Theriogenology, 62: 1198-1217.
Direct Link - Castro, A.C.S., W.E. Berndtson and F.M. Cardoso, 2002. Plasma and testicular testosterone levels, volume density and number of Leydig cells and spermatogenic efficiency of rabbits. Brazil. J. Med. Biol. Res., 35: 493-498.
CrossRefDirect Link - Corah, L.R. and S. Ives, 1991. The effects of essential trace minerals on reproduction in beef cattle. Vet. Clin. North Am. Food Anim. Pract., 7: 41-57.
PubMedDirect Link - Cuanne, S.C., L.S. Harbige and M.A. Crawford, 1993. The importance of energy and nutrient supply in human brain evolution. Nutr. Health J., 9: 219-235.
CrossRefDirect Link - Etiegni, L. and A.G. Campbell, 1991. Physical and chemical characteristics of wood ash. Bioresour. Technol., 37: 173-178.
CrossRef - Gallo, R.V., 1981. Pulsatile LH release during the ovulatory surge on proestrous in the rat. Biol. Reprod., 24: 100-104.
CrossRefDirect Link - Galmessa, U., V.S. Raina, A.K. Gupta and T.K. Mohanty, 2003. Testicular biometry and semen quality in Karan fries bulls. Ind. J. Diary Sci., 56: 317-319.
Direct Link - Hashimoto, I., N. Isomoto, M. Eto, M. Kawaminami, C. Sunazuka and N. Ueki, 1987. Pre-ovulatory secretion of progesterone, luteinizing hormone and prolactin in 4-day and 5-day cycling rats. Biol. Reprod., 36: 599-605.
PubMedDirect Link - Herbert, U., M.O. Ozoeje and D.O. Adejumo, 2005. Effects of Leucaena and Gliricidia leaf meals on seminal characteristics, testis weight and seminiferous tube diameters of rabbits. Anim. Res., 54: 173-178.
CrossRefDirect Link - Hostetler, C.E., R.L. Kincaid and M.A. Mirando, 2003. The role of essential trace elements in embryonic and fetal development in livestock. Vet. J., 166: 125-139.
PubMedDirect Link - Jovanoic-Peterson, L. and C.M. Peterson, 1996. Vitamin and mineral deficiencies which may predispose to glucose intolerance of pregnancy. J. Am. College Nutr., 15: 14-20.
Direct Link - Kyarisiima, C.C., M.W. Okot and B. Svihus, 2004. Use of wood ash in the treatment of high tannin sorghum for poultry feeding. South Afr. J. Anim. Sci., 34: 110-115.
Direct Link - Leach Jr., R.M., 1967. Role of manganese in the synthesis of mucopolysaccharides. Fed. Proc., 26: 118-120.
PubMed - Micaleft, J.V., M.M. Hays, A. Latif, R. Alhasan and S.B. Sufi, 1995. Serum binding of steroid tracers and its possible effects on direct steroid immunoassay. Ann. Clin. Biochem., 32: 566-574.
PubMedDirect Link - Moor, B.C. and E.V. Younglai, 1975. Variation in peripheral levels of LH and Testosterone in adult male rabbits. J. Reprod. Fert., 42: 259-266.
CrossRefDirect Link - Morton, D., 1988. The use of rabbits in male reproductive toxicology. Environ. Health Perspect., 77: 5-9.
Direct Link - Omoikhoje, S.O., A.M. Bamgbose, M.B. Aruna and R.A. Animashahun, 2006. Response of weaner rabbits to concentrate supplemented with varying levels of Syndrella nodiflora forage. Pak. J. Nutr., 5: 577-579.
CrossRefDirect Link - Oso, A.O., A.A. Idowu and O.T. Niameh, 2011. Growth response, nutrient and mineral retention, bone mineralization and walking ability of broiler chickens fed with dietary inclusion of various unconventional mineral sources. J. Anim. Physiol. Anim. Nutr., 95: 461-467.
CrossRefDirect Link - Oyeyemi, M.O. and B.S. Okediran, 2007. Testicular parameters and sperm morphology of chinchilla rabbits fed with different planes of soymeal. Int. J. Morphol., 25: 139-144.
Direct Link - Shuerson, G.C., T.M. Salzer, D.D. Koehler and M.H. Whitney, 2011. Effects of metal specific amino acids complexes and inorganic trace minerals on vitamins stability in premixes. Anim. Feed Sci. Technol., 163: 200-206.
CrossRefDirect Link - Simoni, M. and E. Nieschlag, 1995. FSH in therapy: Physiological basis, new preparations and clinical use. Reprod. Med. Rev., 4: 163-177.
CrossRefDirect Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Townsend, S.F., K.K. Briggs, N.F. Krebs and K.M. Hambridge, 1994. Zinc supplementation selectively decreases fetal hepatocyte DNA synthesis and insulin-like growth factor II gene expression in primary culture. Pediatr. Res., 35: 404-408. Pediatr. Res., 35: 404-408.
PubMedDirect Link - Uboh, F.E., E.E. Edet, M.U. Eteng and E.U. Eyong, 2010. Comparative effect of aqueous extract of P. guajava leaves and ascorbic acid on serum sex hormones levels in male and female rats. J. Applied Sci. Res., 6: 275-279.
Direct Link