
Manuel Gallardo- Cabello
Mexico
Elaine Espino-Barr
Mexico
Arturo Garcia-Boa
Mexico
Esther Guadalupe Cabral-Solis
Mexico
Marcos Puente-Gomez
Mexico
Journal of Fisheries and Aquatic Science
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 131-139







ABSTRACT
The age of the green jack Caranx caballus by the analysis of size frequencies in five age groups and with that of otoliths in four age groups is determined. The growth parameters of von Bertalanffy obtained with the frequency analysis were Loo = 55.38 cm, Woo = 3 841.65 g, k = 0.34, to = -0.0084 and A0.95 = 9 years. The results of the otolith lectures were Loo = 52.02 cm, Woo = 3 205.60 g, k = 0.362, to = -0.085 and A0.95 = 8 years. The growth parameters were compared with those of other species of the same genus. Data were also obtained for the gonadosomatic index which showed their maximum values during the months of June and October, which coincides with the season wherein data of the gastric repletion index increases and the values of the hepatosomatic index diminishes. The data obtained for the condition factor show their largest fluctuations during the period between January and August.
PDF Abstract XML References Citation
How to cite this article
Manuel Gallardo- Cabello, Elaine Espino-Barr, Arturo Garcia-Boa, Esther Guadalupe Cabral-Solis and Marcos Puente-Gomez, 2007. Study of the Growth of the Green Jack Caranx caballus Günther 1868,in the Coast of Colima, Mexico. Journal of Fisheries and Aquatic Science, 2: 131-139.
DOI: 10.3923/jfas.2007.131.139
URL: https://scialert.net/abstract/?doi=jfas.2007.131.139
DOI: 10.3923/jfas.2007.131.139
URL: https://scialert.net/abstract/?doi=jfas.2007.131.139
INTRODUCTION
Caranx caballus (Fig. 1) is an American species of the Pacific Ocean, distributed from the coasts of Mexico to North of Perú (Chirichigno et al., 1982). In the State of Colima, Mexico, it is fished year round and the volume of its catch is only surpassed by the snappers of the Lutjanidae family (Cruz-Romero et al., 1996a, b; Espino-Barr et al., 2003). The green jack represents 6% of the total catch in the State but it is not registered in the fishing statistics of the rest of the country.
This species is captured incidentally during fishing of coastal pelagic species which are commercially important, such as the sierra Scomberomorus sierra. It can be captured with gill net, purse net, seine net, line and hook, spinning tackle or casting net. This species is used locally for human consumption and as bait. Together with other carangid species, it is commercially considered as a second and third class product, with a price ranging from 5 to 8 pesos per kilogram ($ 0.50 to $ 0.80 UDS), therefore it is an accessible food for low income population. The annual catch in the official statistics of the State from 1980 to 2002 varies between 9 and 250 ton, 93 ton average. The fishing effort is 3 000 to 28 000 annual trips. There is an increment in the catch of this species from September to December. The Capture per Unit of Effort (CPUE) is around 4.5 kg/trip, with few variations along the year, the maximum value in May.
 | |
| Fig. 1: | Caranx caballus Günther, 1868 |
Studies on ecology and dynamic of its population have been published by Ramírez-Granados (1977), Castro-Aguirre (1978), Chirichigno et al. (1982), van der Heiden (1985), Cruz-Romero et al. (1986, 1987, 1988, 1989 a, b, 1990, 1993), Espino-Barr (2000) and Gallardo-Cabello et al. (2006).
In terms of population dynamics, age determination is important to ascertain population structure and to analyze changes in its distribution and abundance throughout the year. Study methods on age are either direct or indirect. This latter consists on the analysis of the length frequency to identify polimodal curves corresponding to age groups. Direct methods consider hard structures on which the deposit of materials present a seasonal periodicity according to the availability of the food; these structures show zones of both slow and fast growth which help to determine the age and the calculation of von Bertalanffy (1938) growth parameters equation. On these bases we analyzed the length frequency distribution to obtain the average length for each age and the time of slow and fast growth ring formation in the sagittae. Characteristics of growth in length and weight and longevity were also studied. In addition, the values of the condition factor index, gonadosomatic, hepatosomatic and gastric repletion were obtained to better understand the growth phenomena.
MATERIALS AND METHODS
From January to December 2001, 50 individuals of C. caballus from the commercial captures of the coastal fishery were obtained monthly. The Standard Length (Ls), the total (Wt) and eviscerated weight (We), from the gonad (Wg), stomach (Ws) and liver (Wh) was registered for each organism. Only the standard and total length and total weight of over 200 individuals per month were taken directly in the sampling sites.
The data on length frequencies were analyzed with the FISAT program (Gayanilo et al., 1994) to estimate the average length related to the polimodal curves associated with each age group.
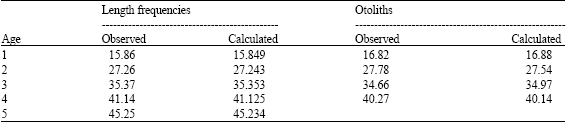
Average length data for each age determined with the sagittae and the asterisci otoliths by Gallardo-Cabello et al. (2006) were used to obtain the growth constants of von Bertalanffy (1938) equation. The observed lengths were: age 1 = 16.82 cm; age 2 = 27.78 cm; age 3 = 34.66 cm; age 4 = 40.27 cm.
The time of growth ring formation was determined analyzing whether the otolith border had a slow or a fast growth. In every case, the otoliths were observed by transparency with transmitted light. The opaque band indicated a fast growth whereas the hyaline band indicated a slow growth, in contrast to what occurs when light is reflected (Blacker, 1974).
There were no differences in the growth between sexes (Gallardo-Cabello et al., 2006).
The growth constants of von Bertalanffy (1938) equation, L∞, k y to were obtained by the method of Ford (1933), Walford (1946) and Gulland (1964) and were adjusted by the convergent iterative method with the Newton algorithm in the Solver program of Excel (Microsoft, 1992).
The function W = a Lb was used to obtain the weight-length relationship. The growth data for length and the weight-length relationship were used to obtain the weight at each age. The weight growth was obtained by substituting Lt and L∞ by Wt and W∞, in the von Bertalanffy (1938) equation. Taylor (1958 and 1960) equation was used to calculate the age limit or longevity (95% of the L∞).
The values of the condition factor were obtained from the a data in the weight-length relation for total and eviscerated weight.
The gonadosomatic (GSI) and the Hepatosomatic (HSI) indexes were determined by the formulas described by Rodríguez-Gutiérrez (1992):
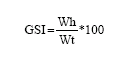 |
where:
| GSI | = | Gonadosomatic index |
| Wg | = | Weight of the gonad |
| Wt | = | Total weight of the organism |
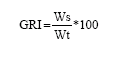 |
where:
| HSI | = | Hepatosomatic index |
| Wh | = | Weight of the liver |
| Wt | = | Total weight of the organism |
The Gastric Repletion Index (GRI) was calculated by the method of Sierra and Popota (1988):
where:
| GRI | = | Gastric repletion index |
| Ws | = | Stomach weight |
| Wt | = | Total weight of the organism |
RESULTS
Time of Growth Rings Formation
Figure 2 shows that the percentage of otoliths with fast growth bands is higher during autumn-winter, i.e., from September to February; while the percentage of otoliths with slow growth bands appear in spring-summer (March to August).
Length Growth
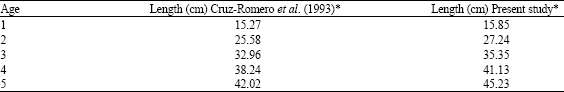
Analysis of length frequency: Table 1 presents the observed average lengths for each age group of C. caballus obtained by the length frequency analysis, by modal progression and by observation of otoliths sagittae and asterisci (Gallardo-Cabello et al., 2006); the values adjusted by the iterative method were also used. During the first year of life, C. caballus had the highest increment in length (15.85 cm) with the analysis of length frequency; thereafter the growth rate decreases: from age one to age two, 11.39 cm, 8.11 cm from age two to three, 5.77 from three to four, 4.11 from age four to five. Results of the analysis of the otoliths indicates that the growth decreases: 10.96 cm from age one to age two, 6.88 from age two to three and 5.60 from age three to four. Growth parameters of the standard length obtained from the otoliths analysis were: L∞ = 52.02 cm, k = 0.362 and to = -0.085.
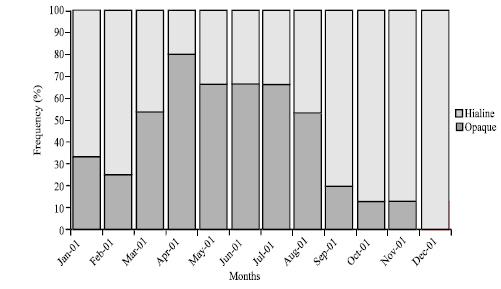 | |
| Fig. 2: | Monthly frequency of the slow (hyaline) and fast (opaque) growth borders in the C. caballus sagittae |
| Table 1: | Average of the standard length (cm) of each age group of C. caballus |
 | |
| Table 2: | Average length for each age of C. caballus in this paper and in Cruz-Romero et al. (1993) |
 | |
| * Length frequencies | |
Table 2 compares the values of the average length of each age with those calculated by Cruz-Romero et al. (1993). In both cases length determination was done with the analysis of length frequencies. The calculated data of this paper are higher in each age (0.5 to 3.5 cm).
Weight Growth
The value of the allometric index in the weight-length relation is b = 2.91 for total weight and b = 2.92 with eviscerated weight. Both values tend to be isometric (Fig. 3) (t = 1.965, p>0.05).
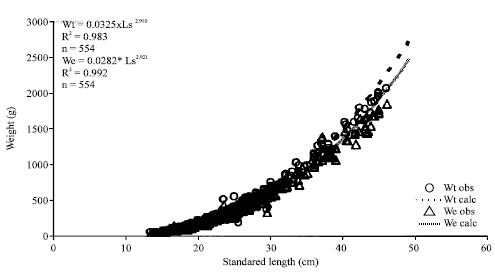 | |
| Fig. 3: | Relation between standard length (Ls) and total (Wt) and eviscerated weight (We) of C. caballus |
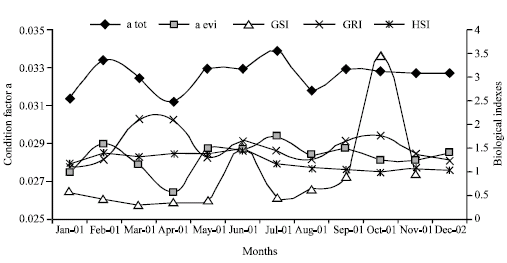 | |
| Fig. 4: | Monthly values of the condition factor (a tot = total weight and a evi = eviscerated weight), of the Gonadosomatic Index in females (GSI), the Gastric Repletion Index (GRI) and the Hepatosomatic Index (HSI) |
Theoretical Weight Growth
The values of theoretical weight growth in total and eviscerated specimens, for each age, obtained with length frequency data and otoliths are shown in Table 3. The values of the analysis of length frequency in weight are: Wt∞ = 3,841.65 g and We∞ = 3,486.65 g and Wt∞ = 3,205.60 g and We∞ = 2,905.04 g, for the results of the analysis of otoliths.
Longevity (Age A0.95)
This species reached 95% of L∞ in 8.2 years which corresponds to an organism of 55 cm of length and an approximate weight of 3 kilograms.
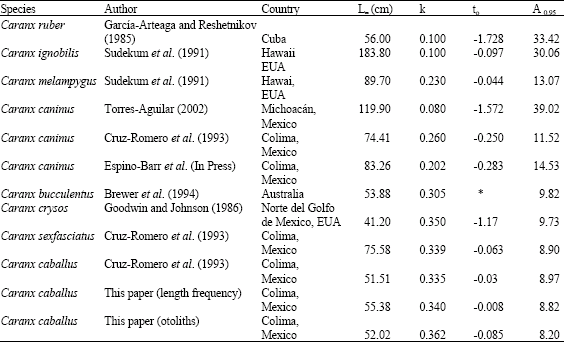
Comparison of Growth Parameters Values of C. caballus with Other Caranx Genus Species
von Bertalanffy’s equation parameters for several species of the Caranx genus are shown in Table 4. Longevity information was calculated from the data reported by Taylor (1958 and 1960).
| Table 3: | Total and eviscerated individual average weight (grams) for each age group of C. caballus |
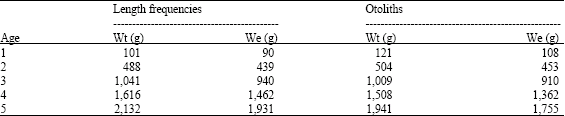 | |
| Table 4: | Values of the growth parameters for different species of the Caranx genus |
 | |
| * Not reported | |
Biological Considerations During the Growth of C. Caballus
Figure 4 shows the monthly values of indexes according to the condition factor (a), the Gonadosomatic (GSI), Hepatosomatic (HIS) and Gastric Repletion (GRI). The period of the highest spawning occurs during October and that of lesser spawning in June. The most active feeding period goes from September to February. The highest values of the RGI are during March-April and September-October.
DISCUSSION
Analysis of the otoliths showed a band of slow growth and another of fast growth are formed each year, which validates age estimate based on the number of rings in the otoliths of C. caballus.
Results of the analysis of length frequency with the application of the ELEFAN method (Gayanilo et al., 1994) and the growth ring determination of the sagittae and asterisci otoliths are very similar, which validate the parameters (Joseph, 1962). In addition, the data of average length of each age found by Cruz-Romero et al. (1993) were also similar. This indicates that the population of C. caballus has maintained its growth characteristics in the last decade, suggesting that this stock has reached its equilibrium and its biomass, due to the growth and recruitment and it decreases due to natural and fishery mortality. This in turn seems to indicate that the artisanal fishery in the coast of Colima does not affect the population biomass, a fact which coincides with the study of Espino-Barr (2000).
With respect of weight-length relation, Cruz-Romero et al. (1993) found an allometric growth index of 2.78; however in this study the value was 2.91, which is an isometric growth index.
Table 4 shows that the different species have a tendency whereby the smaller the k index is (0.08 to 0.26), the higher the L∞ and the longevity, as occurs with Caranx caninus, Caranx ruber, Caranx ignobilis and Caranx melampygus. Caranx caballus belongs to the species of smaller size with a higher value of the k index and a lower L∞ and longevity, as in Caranx crysos and Caranx bucculentus.
During the periods of higher spawning, the fat reserve index (HIS) diminishes, owing to a very high corporal waste of the fish (Nikolsky, 1963). Likewise, during the spawning periods there is a tendency to increase the gastric repletion index, showing an active feeding period that favors the formation of sexual products, both in quantity and quality. The effect of this higher feeding was observed two months later in the increments of the condition factor index for total and eviscerated weight.
The periods of more active feeding coincide with those when fast growth rings are formed in the sagittae, during September to February, i.e., in the rainy season (Observatorio de Manzanillo, personal communication; Espino-Barr, 2000), when more nutrients flow into the river which rushes to the coastal areas and into the sea. This also occurs during hurricanes season (Observatorio de Manzanillo, personal communication; Espino-Barr, 2000). These phenomena remove the bottom of the narrow Colima continental shelf, sending the nutrients to the surface of the ocean and increasing the production of plankton and of organisms that feed from it and form the food web. During this period the weight of the organisms increase and a faster growth in length of the larvae and juveniles takes place and recruit in the fishing area.
On the other hand, during the months of March-August (spring-summer) food availability decreases, the condition factor index diminishes and the slow growth ring are formed in the sagittae.
REFERENCES
- Cruz-Romero, M., E. Espino-Barr and A. Garcia-Boa, 1988. Preliminary estimation of the maximum sustainable yield of the coastal fishery. (Estimacion preliminar del indice de rendimiento maximo sostenible para la captura de escama riberena). CRIP-Manzanillo, Col., INP/SEPESCA, Informe Tecnico, pp: 34.
- Ford, E., 1933. An account of the herring investigations conducted at Plymouth during the years from 1924 to 1933. J. Mar. Biol. Assoc. UK., 19: 305-384.
CrossRefDirect Link - Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the coast of Colima, Mexico. J. Fish. Aquat. Sci., 1: 157-170.
CrossRefDirect Link - Joseph, D.C., 1962. Growth characteristics of two Southern California Surffishes, the California corbina and spotfin croaker, Family Sciaenidae. The resources agency of California and game. Fish Bull., 119: 1-54.
Direct Link - Sierra, L.M. and O.A. Popota, 1988. Digestion velocity, daily feeding rhythm and feeding ratios of some fish in the Batabano Gula, Cuba. (Velocidad de digestion, ritmo diario de la alimentacion y raciones alimenticias de algunas especies de peces del Golfo de Batabano, Cuba.). (Rep. Invest. No. 6). Acad. Cien. Cuba, Instituto de Oceanologia.
- Sudekum, A.E., J.D. Parrish, R.L. Radtke and S. Ralston, 1991. Life history and ecology of large jacks in undisturbed, shallow,oceanic communities. Fish. Bull. US., 89: 493-513.
Direct Link - Taylor, C.C., 1960. Temperature, growth and mortality-the Pacific cockle. J. Conseil., 26: 117-124.
CrossRef - Von Bertalanffy, L., 1938. A quantitative theory of organic growth (Inquiries on growth laws. II). Hum. Biol., 10: 181-213.
Direct Link - Walford, L.A., 1946. A new graphic method of describing the growth of animals. Biol. Bull., 90: 141-147.
PubMedDirect Link