
Ene E. Oku
Department of Zoology and Environmental Biology, University of Calabar, P.M.B. 1115, Calabar, Nigeria
Donald A. Ukeh
Department of Crop Science, University of Calabar, P.M.B. 1115, Calabar, Nigeria
Nsa E. Dada
Department of Zoology and Environmental Biology, University of Calabar, P.M.B. 1115, Calabar, Nigeria
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 3 | Page No.: 279-285







ABSTRACT
Most rainforest areas in West and Central Africa are endemic to various biting haematophagous insects that transmit pathogens to man and animals. Field surveys were conducted during the raining and dry seasons of 2006 to investigate the prevalence, seasonal distribution and intensity of biting flies in Rhoko forest, Iko Esai, Akamkpa local government area, in Cross River State, Nigeria. The rain forest was divided into four locations based on human activity and geographical sub-locations. Data obtained showed the mean prevalence of four genera of biting flies namely; Simulium spp. (77%), Chrysops spp. (16%), Glossina spp. (5%) and Tabanus spp. (2%), respectively. A greater number of the flies were caught in locations where human activity was greatest. The result also showed that the mean number of flies caught during the wet season was significantly (p<0.05) higher than the dry season while the peak period of fly activity was between 11.00 am and 3.00 pm. Present results suggest that visitors should be informed about the entomological data of Rhoko forest as a management strategy between the vectors and potential hosts.
PDF Abstract XML References Citation
Received: February 21, 2011;
Accepted: March 15, 2011;
Published: May 30, 2011
How to cite this article
Ene E. Oku, Donald A. Ukeh and Nsa E. Dada, 2011. Prevalence and Seasonal Distribution of Daytime Biting Diptera in Rhoko Forest in Akamkpa, Cross River State, Nigeria. International Journal of Zoological Research, 7: 279-285.
DOI: 10.3923/ijzr.2011.279.285
URL: https://scialert.net/abstract/?doi=ijzr.2011.279.285
DOI: 10.3923/ijzr.2011.279.285
URL: https://scialert.net/abstract/?doi=ijzr.2011.279.285
INTRODUCTION
Biting flies are external insects that feed on the blood of vertebrate hosts and feed actively in the daytime. Their irritating bites could transmit pathogenic organisms that cause devastating loss of human and animal life (Logan and Birkett, 2007). The impacts of invasion of biting insects also have a huge cost to the economy of many countries (Okoth et al., 2007; Mamoudou et al., 2008). The most important groups of biting flies are mosquitoes, Tsetse flies (Glossina spp.) (Diptera: Glossinidae), black flies (Simulium spp.)(Diptera: Simuliidae), sand flies (Chrysops spp.) (Diptera: Psycholidae) and tabanids (Tabanus spp.) (Diptera: Tabanidae). Mosquitoes are well known for their spread of malaria, tallow fever, dengue, encephalitis and elephantiasis. Leishmaniasis, caused by the protozoan Leishmania is transmitted by sand flies. Tabanids are the vectors of Loaiasis caused by the helminth, Loa Loa or the African eye worm because the adult worm can be seen moving through the sclera of the eye (Padgett and Jacobsen, 2008). They have also been implicated as vectors of human cases of tularaemia due to Francisella tularensis and anthrax caused by Bacillus anthracis (Azar and Nel, 2003). The bites of black flies cause pain and dermatitis at the site of the bite due to the toxicity of their saliva and intense itching may last several days and serious allergic reactions may occur. Death may also result from suffocation, as a result of plugged nasal and bronchial tubes, exsanguinations and allergic reaction. Simulium spp. are known to transmit Onchocerciasis (river blindness) to humans and Leucocytozoon parasites to turkeys and wild birds. They have also been reported as transmitters of encephalitis (Matthews et al., 2009). Bite wounds resulting from these flies may lead to secondary bacterial infections (Okon and Asor, 2006). Socially, biting flies are significant because they can discourage people from remaining in or visiting certain areas such as recreational facilities thereby limiting their maximum utilisation.
Many agricultural estates and forest reserved areas characterized by complex arthropod diversity and food webs are located in southern Nigeria as a result of the favourable climatic conditions (Banjo et al., 2006; Ukeh, 2007). Rhoko forest located in the Niger Delta region (one of the world’s largest wetlands) of southern Nigeria is a protected area which covers one of the thickest tropical rain forests in West Africa with diverse plant and animal species, most of which are of ecological and medicinal importance (Ubom, 2010), have not yet been discovered or are in danger of extinction. Researchers, students and tourists from all over the world visit Rhoko forest to carry out their studies or for recreational purposes. Unconfirmed reports from previous visits to this area have shown that biting flies exists in the forest. It has been reported that vector diversity, abundance and distribution could influence transmission risk to host organisms because insect vectors select their preferred hosts from a pool of available susceptible hosts (Okoth et al., 2007; Udoidung et al., 2007). Therefore, the objectives of this study were to identify the various groups of daytime biting flies prevalent in Rhoko forest, study their distribution within the forest, their daily peak periods and variations in their seasonal abundance.
MATERIALS AND METHODS
Study area and field sampling: The study was conducted in Cercopan field site of Rhoko forest located in Iko Esai village, Akamkpa Local Government Area of Cross River State which is situated between latitude 5°00′and 5°15′ North and longitude 8°04′ and 8°25′ East in southern Nigeria (Ukeh, 2008). The forest is a protected area surrounding the Oban Hills Division of Cross River National Park where no human activity other than research and tourism is allowed to take place. The core area of the forest is approximately 400 hectares in size supporting an incredible variety of plant and animal life. The climate is governed by two seasons-the wet season running from April to November and a short dry season from December to March. The altitude of the study area is about 100-200 m above sea level while topography varies over the area. Annual rainfall of about 160 and 220 mm, relative humidity range of 80-90% and temperature range of 23.4-39.69°C have been reported for this area (Udoidung et al., 2007).
Four locations were selected based on the nature of habitats and human activities and sampling points were selected randomly within these locations. The locations were A (Education centre), location B (where visitors and staff lodge is surrounded by thick forest and small fast flowing streams), location C (Rhoko river which is a major drainage, fast flowing, unpolluted river surrounded by thick forest land) and location D (southern region, a typical tropical rain forest where all human activities except research and tourism are prohibited) (Fig. 1). Sampling was carried out in January, February, April and May 2006 to cover the dry and wet seasons.
Sample collection, preservation and identification: Fly catching was done thrice a day in all locations. At each location, samples were collected at intervals of 4 h as follows: morning (7.00 am-11.00 am), afternoon (11.00 am-3.00 pm) and evening (3.00 pm-7.00 pm). Flies hovering were collected using a sweep net while those settling to feed were trapped by inverting sample bottles over them or hand picked when they were fully engorged.
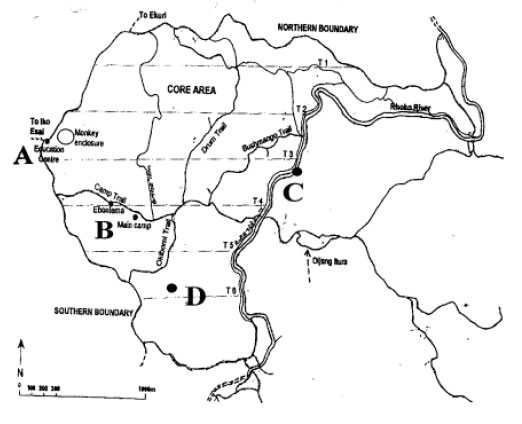 | |
| Fig. 1: | Map of study area (Rhoko forest in Nigeria) showing the different study sites |
Flies caught were counted and recorded for each species and time period and placed in labelled plastic sample bottles containing 70% alcohol. The flies remained in 70% alcohol for two days after which they were pinned on corks and placed in an oven maintained at 40°C for three days. After three days the corks were placed in insect boxes containing camphor for identification. Flies were identified to the genus level by microscopic examination based on standard taxonomic keys.
Data analysis: Data were transformed using square root transformation (![]() x+0.5) for homogeneity before analysed as a factorial experiment based on the randomized Complete Block Design to show the effect of time and seasonal variation on all flies at different locations and the effect of time on the flies at different locations in wet and dry seasons, respectively. A chi square test was also used to show if there was a significant difference in the seasonal abundance of each group of flies using GenStat 12th Edition Statistical Software.
x+0.5) for homogeneity before analysed as a factorial experiment based on the randomized Complete Block Design to show the effect of time and seasonal variation on all flies at different locations and the effect of time on the flies at different locations in wet and dry seasons, respectively. A chi square test was also used to show if there was a significant difference in the seasonal abundance of each group of flies using GenStat 12th Edition Statistical Software.
RESULTS AND DISCUSSION
Data obtained showed that four genera of flies were prevalent during the study namely Simulium spp. Glossina spp., Tabanus spp. and Chrysops, with Simulium spp. being the most abundant while Tabanus spp. was the least abundant (Fig. 2). Variation in the prevalence of the fly species was observed at the various sites sampled as well, with location D having the least prevalence of the flies. Simulium species were most abundant in location C, followed by B, A and D in that order (Fig. 3). There was also a significant difference (p<0.01) in the effect of season on the abundance of flies and this difference was observed for each species. The wet season recorded the highest number of all available species of biting flies in Rhoko forest (Fig. 4).
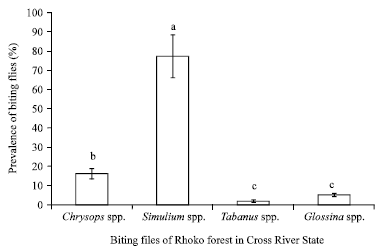 | |
| Fig. 2: | Mean % (±SE) prevalence of biting flies of Rhoko forest. Error bars = Standard Errors (SE) of the means. Bars with different letters are significantly (p<0.05) different from each other |
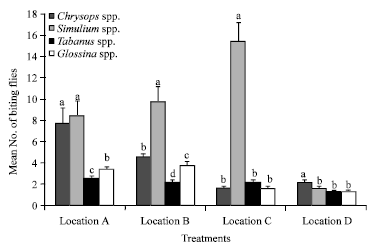 | |
| Fig. 3: | Mean±SE of biting flies at different locations of Rhoko forest. Error bars = Standard Errors (SE) of the means. Bars with different letters are significantly (p<0.05) different from each other |
During the wet season, Simulium spp. was the most prevalent biting insect recorded in the forest, followed by Chrysops spp., Glossina spp. and Tabanus spp. But in the dry season, although Simulium spp. remained significantly most prevalent, there was no significant difference (p>0.05) between Glossina and Tabanus species. Similarly, the time or period of sampling had significant effect on the number of flies caught. Relatively, fly abundance was highest in the afternoon than in the morning and evening (Fig. 5). It could be worthy to note specifically that Simulium species was significantly (p<0.05) more abundant followed by Chrysops, Glossina and Tabanus spp., respectively in the morning and evening. However while Simulium spp. remained prevalent followed by Chrysops spp. in the evening, there was no significant difference between the mean numbers of Glossina and Tabanus spp.
Results from this field study show the presence and intensity of biting flies in Rhoko forest, an area frequented for research and tourism in Akamkpa local government area of Cross River State, Nigeria.
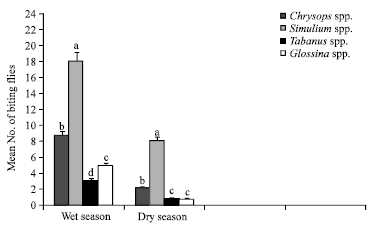 | |
| Fig. 4: | Differential abundance of biting flies (±SE)in wet and dry seasons. Error bars = Standard Errors (SE) of the means. Bars with different letters are significantly (p<0.05) different from each other |
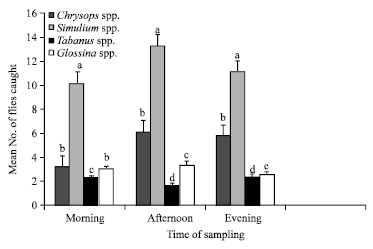 | |
| Fig. 5: | Differential abundance of biting flies (±SE) at different time intervals. Error bars = Standard Errors (SE) of the means. Bars with different letters are significantly (p<0.05) different from each other |
The study revealed a high prevalence of Simulium species (77%) and low incidence of Glossina species (5%) and Tabanus species (2%), respectively. There was a seasonal variation in the abundance of flies with more flies caught during the raining (wet) season than dry season. Similar results have been reported on the seasonal distribution and abundance of biting flies in Adamaoua plateau of Cameroon which has identical vegetation (Mamoudou et al., 2008). Rhoko forest extends into the Cameroon forest reserve in the south west province of Cameroon. Therefore, land covers such as evergreen forest trees and shrubs which are characteristics of the raining season have been reported to favour the prevalence of insect vectors such as Glossina species (Cecchi et al., 2008). The incidence of these flies increased drastically at the onset of rain storms (Cranshaw et al., 2005). The raining season is generally accompanied by flooding and phytoplankton richness and these constitutes food required by Simulium larvae for its development (Effenberger et al., 2008). This may be responsible for the high incidence of Simulium species in Rhoko forest which has several small, unpolluted rivers throughout the study period. Present findings are in agreement with Adandédjan et al. (2011), who attributed the prevalence and distribution of benthic insect fauna in the West African lagoon of Porto-Novo to hydrological and human activities as determining factors. The very low incidence of Tabanus and Glossina species in the dry season suggest that water could be essential for their development and prevalence. Furthermore, low prevalence of Glossina species could be attributed to the low risk of transmission factor which is primarily linked to the intensity of the encounters between animals (such as cattle) and vectors. In Nigeria, two-thirds of the nation’s livestock resources are practiced under semi-intensive and extensive systems predominantly practiced in the guinea and Sudan savannah regions of north (Samdi et al., 2010; Fajinmi et al., 2011). The peak activity of Chrysops species was in the afternoon but Simulium species continued throughout the day. This finding is in agreement with Wanji et al. (2003), who reported that Chrysops species are more prevalent during the wet season than dry season with peak activity in the morning and in the afternoon.
Data also showed that virtually all the four genera of biting flies were very prevalent in locations A and B, areas with more human occupation and activities. The flies generally seemed to be more prevalent in locations where human and animal habitation was most common since the insects feed on vertebrate blood (Udoidung et al., 2007). This could be the result of the release of human or animal semiochemicals (infochemicals) such as kairomones or attractants from the human body. It has been documented that during host location, haematophagous insects that bite vertebrates to obtain blood such as black flies, tsetse flies, horseflies and sand flies use olfactory and visual cues to locate their hosts (Logan and Birkett, 2007; Logan et al., 2008). These insects have a highly developed olfactory system and mainly use their antennae and in some cases, maxillary palps to detect infochemicals. The known olfactory stimuli from long range orientation may include ammonia, carbondioxide and 1-octen-3-ol (Bhasin et al., 2000). However, physical cues such as heat, moisture and vision play a prominent role to such insects during orientation and landing at close range to their host (Eiras and Jepson, 1994).
Rhoko forest is a reserved area which is frequented by tourists and researchers all over the world but the forest harbours daytime biting flies that have been implicated as vectors of various diseases such as loiasis, trypanosomiasis and onchocerciasis. The state and federal governments should take note of the health implications of these vector flies since they make outdoor activities uncomfortable in the forest. Data presented in this baseline study should be adequately utilized by the relevant authorities for the promotion of sustainable conservation and tourism potential of Rhoko forest in particular and other protected areas in general.
REFERENCES
- Adandedjan, D., P. Laleye, A. Ouattara and G. Gourene, 2011. Distribution of benthic insect fauna in a West African Lagoon: The Porto-Novo Lagoon in Benin. Asian J. Biol. Sci., 4: 116-127.
CrossRefDirect Link - Azar, D. and A. Nel, 2003. Fossil psychodoid flies and their relation to parasitic diseases. Mem. Inst. Oswaldo Cruz, 98: 35-37.
PubMed - Banjo, A.D., O.A. Lawal and S.A. Aina, 2006. Insect diversity of two medicinal Labiatae in Southwestern Nigeria. J. Entomol., 3: 298-304.
CrossRefDirect Link - Cecchi, G., R.C. Mattioli, J. Slingenbergh and S. De La Rocque, 2008. Land cover and tsetse fly distributions in sub-Saharan Africa. Med. Vet. Entomol., 22: 364-373.
CrossRef - Effenberger, M., J. Engel, S. Diehl and C.D. Matthaei, 2008. Disturbance history influences the distribution of stream invertebrates by altering microhabitat parameters: A field experiment. Freshwater Biol., 53: 996-1011.
CrossRef - Eiras, A.E. and P.C. Jepson, 1994. Responses of female Aedes aegypti (Diptera: Culicidae) to host odours and convection currents using an olfactometer bioassay. Bull. Entomol. Res., 84: 207-211.
CrossRef - Fajinmi, A.O., O.O. Faleke, A.A. Magaji, A.I. Daneji and M. Gweba, 2011. Presence of trypanosome species and determination of anaemia in trade cattle at Sokoto abattoir, Nigeria. Res. J. Parasitol., 6: 31-42.
CrossRef - Logan, J.G., M.A. Birkett, S.J. Clark, S. Powers and N.J. Seal et al., 2008. Identification of human-derived volatile chemicals that interfere with attraction of Aedes aegypti mosquitoes. J. Chem. Ecol., 34: 308-322.
CrossRef - Logan, J.G. and M.A. Birkett, 2007. Semiochemicals for biting fly control: Their identification and exploitation. Pest Manage. Sci., 63: 647-657.
PubMed - Mamoudou, A., A. Zoli, H. Hamadama, Bourdanne and S. Abah et al., 2008. Seasonal distribution and abundance of tsetse flies (Glossina spp.) in the faro and deo division of the adamaoua plateau in Cameroon. Med. Vet. Entomol., 22: 32-36.
PubMed - Matthews, G.A., H.M. Dobson, P.B. Nkot, T.L. Wiles and M. Birchmore, 2009. Preliminary examination of integrated vector management in a tropical rainforest area of Cameroon. Trans. Royal Soc. Trop. Med. Hygiene, 103: 1098-1104.
CrossRef - Okoth, S.O., E.D. Kokwaro, J.M. Kiragu and G.A. Murilla, 2007. Diurnal activity of allopatric populations of Glossina pallidipes and hosts and their implications on risk of transmission of sleeping sickness in Kenya. J. Entomol., 4: 20-32.
CrossRefDirect Link - Padgett, J.J. and K.H. Jacobsen, 2008. Loiasis: African eye worm. Trans. Royal Soc. Trop. Hyg., 102: 983-989.
PubMed - Samdi, S.M., A.O. Fajinmi, J.O. Kalejaye, B. Wayo and M.K. Haruna et al., 2010. Periodic variation in trypanosoma infection rates in trade small ruminants at slaughter in Kaduna Central Abattoir. Res. J. Vet. Sci., 3: 189-193.
CrossRef - Ubom, R.M., 2010. Ethnobotany and biodiversity conservation in the Niger Delta, Nigeria. Int. J. Botany, 6: 310-322.
CrossRefDirect Link - Udoidung, N.I.G., I.E. Braide, K.N. Opara and H.A. Adie, 2007. Perstans filariasis in rural communities of the lower cross river basin nigeria: Parasitological observations. Int. J. Zool. Res., 3: 207-212.
CrossRefDirect Link - Ukeh, D.A., 2007. Distribution of Latoia viridissima and Oryctes monoceros in cross river oil palm estates, Nigeria. Int. J. Agric. Res., 2: 883-887.
CrossRefDirect Link - Ukeh, D.A., 2008. Bioactivities of essential oils of Aframomum melegueta and Zingiber officinale both (Zingiberaceae) against Rhyzopertha dominica (Fabricius). J. Entomol., 5: 193-199.
CrossRefDirect Link - Wanji, S., N. Tendongfor, M. Esum, S.N. Atanga and P. Enyong, 2003. Heterogeneity in the prevalence and intensity of loiasis in five contrasting bioecological zones in Cameroon. Trans. Royal Soc. Trop. Med. Hyg., 97: 182-187.
PubMed