
M.Mohammad- Shahi
Nutrition Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran
H. Rafiei
Department of Nutrition, Faculty of Para medicine, Jundishapur University of Medical Sciences, Ahvaz, Iran
M. Karandish
Diabetes Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran
K. Omidian
Department of Nutrition, Faculty of Para medicine, Jundishapur University of Medical Sciences, Ahvaz, Iran
F. Haidari
Nutrition Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran
Asian Journal of Biochemistry
Year: 2012 | Volume: 7 | Issue: 2 | Page No.: 98-105







ABSTRACT
It seems that soy isoflavones can improve obesity and reverse subsequent metabolic disorders. In this study we assessed the effect of restriction of calorie supplemented with genistein on diet-induced obese rats. Thirty male Wistar obese rats were divided randomly into 3 experimental groups (n = 10) as follows: group 1: low calorie diet supplemented with 50 mg kg-1 genistein, group 2: low calorie diet supplemented with dimethyl sulphoxide (as vehicle) and group 3: obese control rats with ad libitum access to standard food. The animals were carefully monitored and weighed daily. After 4 weeks, fasting blood samples were collected and analyzed for biochemical analysis. The results showed that the restriction of calorie intake caused to weight loss and subsequently led to a significant decrease in serum glucose (p<0.001), lipid profile (p<0.001) and hsCRP levels (p<0.001) but had no effect on resistin levels. Although, low calorie diet supplemented with genistein improved the lipid lowering effect of calorie restriction but this had no more effect on glucose, resistin and hsCRP levels. These results strongly suggest that low calorie diet supplemented with genistein is effective in weight management and has hypoglycemic, hypolipidemic and anti-inflammatory activities. However, it is not recommended before further investigations in animals and humans.
PDF Abstract XML References Citation
Received: November 02, 2011;
Accepted: December 19, 2011;
Published: March 08, 2012
How to cite this article
M.Mohammad- Shahi, H. Rafiei, M. Karandish, K. Omidian and F. Haidari, 2012. Effect of Calorie Restriction Supplemented with Genistein on Serum Levels of Glucose, Lipid Profile and Inflammatory Markers (Resistin and hsCRP) in Obese Rats. Asian Journal of Biochemistry, 7: 98-105.
DOI: 10.3923/ajb.2012.98.105
URL: https://scialert.net/abstract/?doi=ajb.2012.98.105
DOI: 10.3923/ajb.2012.98.105
URL: https://scialert.net/abstract/?doi=ajb.2012.98.105
INTRODUCTION
The prevalence of obesity worldwide has increased dramatically in the past decade (Lazar, 2005). Obesity is a risk factor for different health problems related to coronary heart disease, diabetes, hyperlipidemia and cancer; and its incidence is rapidly increasing (Zimmermann-Belsing and Feldt-Rasmussen, 2004; Ogden et al., 2006). Therefore, there is increased need to develop policies that will be effective for both the treatment and prevention of obesity.
Fat mass can be regulated by different factors, including estrogens which maintain, promote and control the distribution of fat and affect adipose tissue metabolism. The factors that regulate adipose tissue metabolism are of special interest because of rising in obesity and obesity-related problems (Kopelman, 2000). Increasing body of studies has focused on the role of nutritional factors in the etiology, prevention and treatment of obesity (Astrup et al., 2008). Among these, studies on the role of phytoestrogens in obesity have been focused as an important research area in the last 7 years (Cope et al., 2008; Orgaard and Jensen, 2008; Rayalam et al., 2008).
Phytoestrogens are plant-derived estrogens that can bind to both estrogen receptor-α (ERα) and estrogen receptor-β (ERβ) and mimic the actions of estrogens on target tissues (Naaz et al., 2008). The isoflavone genistein (4,5,7 trihydroxyisoflavone) is a phytoestrogen that is found in high concentrations in soy and soy products (Reinli and Block, 1996). It has attracted much attention among medical and public communities because of its possible role in prevention and treatment of many diseases including diabetes, menopausal symptoms, osteoporosis, arthritis, cardiovascular diseases, renal diseases and various types of cancers (Mohammad Shahi et al., 2011). Beneficial effects of genistein on glucose and lipid metabolism have also been reported by some studies and are attributed to its effect on Peroxisome Proliferator-Activated Receptor (PPAR) (Mezei et al., 2003; Kim et al., 2004).
Genistein and other soy isoflavones like daidzein are consumed by Oriental populations at concentrations up to 1 mg kg-1 per day and infants fed the soy formula consume even higher quantities of isoflavones (Setchell et al., 1997).
Although, several studies have shown that genistein is beneficial for hyperlipidemia and cardiovascular diseases and may help to improve obesity and diabetes (Banz et al., 2004; Jones et al., 2005) but the results remain controversial and need further investigation. The present study examined the effect of low calorie diet supplemented with genistein on body weight, serum glucose, lipid profile and inflammatory biomarkers including resistin and hsCRP (high sensitive C reactive protein). In this study, we hypothesized that the supplementation of low calorie diet with genistein can synergistically improve the serum glucose and lipid profiles as well as the increased concentrations of resistin and hsCRP in obese rats.
MATERIALS AND METHODS
Animals: Thirty six male Wistar rats (150-170 g), aged 7-8 weeks, were obtained from Physiology Research Center of Jundishapur University of Medical Sciences. The animals were housed under standard conditions (23±4°C, 57±4% humidity and a 12-h light/dark cycle) and had free access to ad libitum standard laboratory diet and water for 2 weeks for acclimatization. The study was conducted in consistent with ethical procedures and policies approved by the Animal Care and Use Committee of Jundishapur University of Medical Sciences, Ahvaz, Iran.
High fat diet- induced obesity: Obesity was induced in rats by feeding the rats with a High Fat Diet (HFD) for 6 weeks (Guo et al., 2009). The high fat diet consisted of 40% of calorie from fat, 20% from protein and 40% from carbohydrate (Table 1). Each gram of this diet contained 19.6 KJ (4.7 kcal). Body weight and weight of the ingested food were measured once within two days.
After feeding the high-fat diet for 6 weeks, a significant weight gain in HFD group was observed as compared to control group (254±22 g vs. 196±7 g; p = 0.000).
Study design: Thirty high-fat induced obese rats were divided randomly into three groups (n = 10) and treated as follows: Group 1: obese rats taking low calorie diet supplemented with 50 mg kg-1 genistein; Group 2: obese rats taking low calorie diet supplemented with dimethyl sulphoxide (DMSO) and Group 3: obese rats having free-access to standard laboratory diet (as control). Genistein (GE) and DMSO (DM) were given by oral gavage and the treatment continued for four weeks. Food intake and body weight were controlled daily during the intervention.
| Table 1: | Composition of the high-fat diet fed to rats |
 | |
| 1Mineral mixture for AIN-76A rodent diet; 2Vitamin mixture for AIN-76A rodent diet | |
During calorie restriction, the food intake of GE and DM groups were restricted to 40% of the energy available to their respective controls (22.63±5.00 and 23.45±6.01 kcal day-1, respectively, vs. 54.36±7.05 kcal day-1).
Biochemical analysis: At the end of the study, fasting blood samples were collected directly through the heart under light ether anesthesia. Sera were obtained by centrifuging the blood samples at 4000 rpm for 10 min at 4°C and stored in-70°C until assayed. Serum hsCRP and resistin concentrations were assayed by ELISA technique using commercial kits (BioVendor, Czech Republic) according to manufacturer's instructions. Fasting blood glucose, triglycerides (TG), Total Cholesterol (TC) and HDL-C levels were measured enzymatically using standard kits by autoanalyser SA1000. LDL-C level was calculated by Friedwald formula as follows:
LDL cholesterol = Total cholesterol-HDL cholesterol-(triglyceride/5) |
Statistical analysis: Statistical analyses were carried out by SPSS 17 program for windows. Data were expressed as Mean±SD. Statistical analysis was performed by one way ANOVA, Independent sample t-test and Paired sample t-test with 95% Confidence Interval (CI). Differences were considered to be statistically significant at p<0.05.
RESULTS
The initial and final body weights of experimental groups are summarized in Table 2. As it is shown in the table, the initial body weight of 3 intervention groups were equal and the difference between means was not statistically significant (p = 0.985). Restriction of the calorie resulted in a significant weight loss in GE and DM group compared to control group (p = 0.000).
The body weight changes during the intervention (4 weeks) in experimental groups are also presented in Fig. 1. At the end of the study, the weight of rats in GE group was lower than that of DM group but it is not statistically significant (p = 0.628).
The means of serum glucose and lipid profile are shown in Table 3. As it is indicated in the table, serum glucose levels of obese control rats is significantly higher than those of GE and DM groups (p = 0.001 and p = 0.002, respectively). But the difference between means of glucose concentration in GE and DM group was not statistically significant (p = 0.927).
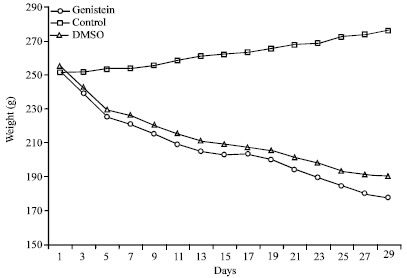 | |
| Fig. 1: | Effect of calorie restriction supplemented with genistein on body weight changes |
| Table 2: | Means of body weight in experimental groups at the beginning and the end of study |
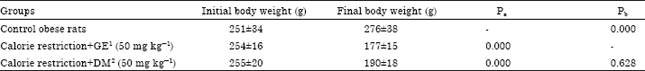 | |
| All values are expressed as Mean±SD (n = 10). Pa indicates p value vs. control group and Pb indicates p value vs. GE group (independent sample t-test, CI 95%). 1 Genisterin, 2Dimethyl sulphoxide | |
| Table 3: | Means of serum glucose and lipid profile levels in experimental groups at the end of study |
 | |
| All values are expressed as Mean±SD (n = 10). Independent-sample t-test was used for statistical significance assessment. †indicates p<0.05 and ††indicates p<0.001 vs. control group; *indicates p<0.05 and **indicates p<0.001 vs. GE group. 1Genisterin, 2Dimethyl sulphoxide | |
| Table 4: | Means of serum resistin and hsCRP levels in experimental groups at the end of study |
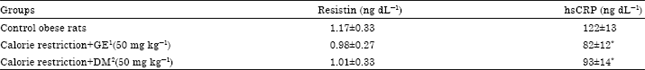 | |
| All values are expressed as Mean±SD (n = 10). Independent-sample t-test was used for statistical significance assessment. *indicates p<0.001 vs. control group. 1Genisterin, 2Dimethyl sulphoxide | |
Serum TG, TC and LDL-C were significantly higher in the obese control rats than those of GE and DM groups (p = 0.000 for all). Obese control rats had also significant lower levels of HDL-C than those of treated groups (p = 0.000). Following treatment of obese rats with genistein, the serum levels of TG, TC and LDL-C were decreased and HDL-C concentration was increased significantly (p<0.05).
In the Table 4, the effects of low calorie diet supplemented with genistein on serum levels of resistin and hsCRP are shown. As it is obvious in the table, there was no significant difference in resistin levels between control group and treated groups (GM and DM groups) (p = 0.519 and p = 0.654, respectively). However, low calorie diet supplemented with or without genistein significantly decreased the levels of hs-CRP compared to control group (p<0.001). We also found that genistein supplementation is not more effective on resistin and hsCRP levels rather than calorie restriction alone.
The present data also indicated a strong relation between hsCRP levels and body weight (r = 0.653, p = 0.000) but the relation between resistin levels and body weight was not statistically significant (p>0.05).
DISCUSSION
Increasing body of evidence suggest that soy and its isoflavones constituents can lead to the reduction in food intake, body weight and adipogenesis (Orgaard and Jensen, 2008; Kishida et al., 2008). However, in our study we found that the supplementation with 50 mg kg-1 genistein had no more effect on body weight rather than calorie restriction alone. The results regarding the effect of genistein on body weight and adipogenesis are controversial and need further investigation. Some studies showed that genistein like esterogen inhibit Lipoprotein Lipase (LPL) in adipose tissue (Naaz et al., 2008). Moreover, several evidences have established that isoflavones not only act via ERs but also exert their effects through other pathways, including those regulated by PPARs. Unlike the highly specific ERs, PPARs can bind to various number of ligands and directly affect lipid metabolism by increasing transcription of PPAR-regulated genes (Kishida et al., 2008). Genistein can also act as a tyrosine kinase inhibitor and therefore, contributes to adipocyte differentiation (Kishida et al., 2008). In our study, the effect of calorie restriction on the rate of weight loss was so strong that may be masked the effect of genistein on body weight. Also, it seems that the duration of the intervention and the dose of genistein are critical factors in this context.
In this study we also found that genistein can significantly decrease the levels of TG, TC, LDL-C and increase HDL-C. Kirk et al. (1998) demonstrated that the ability of the isoflavones genistein and daidzein to decrease serum cholesterol levels may be attributed to an increase in LDL-C receptor activity which is consistent with other results from a in vitro studies (Owen et al., 2004; Mullen et al., 2004).
In the present study we also found that low calorie diet treated groups (GE and DM groups) had lower serum glucose levels in comparison to control group that had ad libitum access to food. It seems that restriction of food intake and related decrease in body weight is responsible for low levels of glucose in low calorie diet treated rats versus obese control rats. But, genistein supplementation had no significant effect on serum glucose levels in GE treated group in comparison to DM group. Our finding is in concert with Amani et al. (2005). They found that soy isoflavones can ameliorate lipid profile but have no effect on blood glucose levels in hypercholestrolemic rabbits. But, Cederroth et al. (2008) showed that treatment of male CD-1 mice with soy phytoesterogens improves insulin sensitivity, at least in part, by activating Adenosine Mono Phosphate Kinase (AMPK) in various tissues, including skeletal muscles and white adipose tissue. Another mechanisem by which genistein can improve glucose levels was defined by Lee (2006). They showed that soy protein and genistein can increase glucokinase and decrease glucose-6-phosphatase in streptozocin-induced diabetic rats (Lee,2006). However, it seems that in our study the rigid restriction of calorie had so strong effect on serum glucose levels that masked the effect of genistein on glucose. Additionally in the Cederroth et al. (2008) study, mice were fed with high phytoestrogen diet from conception and maybe short duration of intervention in our study resulted in null effect of genistein on blood glucose.
In the present study, low calorie diet supplemented with genistein had no effect on serum resistin levels in obese rats. There are a few studies about the effects of isoflavones on resistin levels. Chen et al. (2006) investigated the effect of soy isoflavones on gene expression of resistin in insulin resistant rats fed a high-fat diet. They found that treatment with 450 mg kg-1 isoflavones can decrease resistin levels. One possible reason that genistein has no effect on resistin levels in our study is that the dose of genistein in our study is lower than Chen et al. (2006) study. Another possible explanation is that the duration of our treatment with genistein is short (only 4 weeks) and maybe longer treatment could affect on resistin levels. However, the exact mechanism by which isoflavones decrease resistin levels is not yet determined and further investigations is needed to clarify this.
The results of the present study exhibit lower levels of hsCRP in obese rats taking low calorie diet than those in obese control rats. There has been also a positive correlation between serum hsCRP concentration and glucose levels and body weight in our study which could simply explain that this reduction could be due to the weight loss of low calorie diet- treated groups. This finding is in agreement with other studies. Selvin et al. (2007) in a Meta analysis study also reported that weight loss results in reduction of serum hsCRP levels.
In present study we found that genistein supplementation is not more effective on hsCRP levels rather than calorie restriction alone. This result is consistent with other studies (Yildiz et al., 2005; Jenkins et al., 2002). One possible explanation is that hsCRP level is closely related to energy intake and rigid restriction of calorie in our study may be inhibit the effect of genistein on hsCRP level but it seems that this area of research requires further investigations.
CONCLUSION
This study concludes that the restriction of calorie intake is effective in weight management and controlling of glucose and lipid profiles in obese rats and possesses anti-inflammatory effect. Although, low calorie diet supplemented with genistein improves the lipid lowering effect of calorie restriction but this has no more effect on glucose, resistin and hsCRP levels. In this study we used only one dose of genistein, so maybe higher doses of it have favorable effect on these parameters. Soy isoflavones, thus, should be considered as an excellent candidate for future investigation in obesity.
ACKNOWLEDGMENT
This study is part of MSc thesis for Hossein Rafiei. Special thanks to Ahvaz Jundishapur University of Medical Sciences for the financial support.
REFERENCES
- Astrup, A., J. Dyerberg, M. Selleck and S. Stender, 2008. Nutrition transition and its relationship to the development of obesity and related chronic diseases. Obese Rev., 9: 48-52.
PubMed - Banz, W.J., J. Davis, R. Peterson and M.J. Iqbal, 2004. Gene expression and adiposity are modified by soy protein in male Zucker diabetic fatty rats. Obese Res., 12: 1907-1913.
PubMed - Cederroth, C.R., M. Vinciguerra, A. Gjinovci, F. Kuhne and M. Klein et al., 2008. Dietary phytoestrogens activate AMP-activated protein kinase with improvement in lipid and glucose metabolism. Diabetes, 57: 1176-1185.
CrossRefDirect Link - Chen, S.W., H.M. Zhang, L.S. Zhang and X.F. Feng, 2006. Effects of soy isoflavone on gene expression of resistin in insulin-resistance rats. Sichuan Da. Xue. Xue. Bao. Yi. Xue. Ban., 37: 717-720.
PubMed - Cope, M.B., J.W. Jr. Erdman and D.B. Allison, 2008. The potential role of soyfoods in weight and adiposity reduction: An evidence-based review. Obese Rev., 9: 219-235.
PubMed - Guo, Y., G. Wu, X. Su, H. Yang and J. Zhang, 2009. Antiobesity action of a daidzein derivative on male obese mice induced by a high-fat diet. Nutr. Res., 29: 656-663.
PubMed - Jenkins, D., C.W. Kendall, P.W. Conelly, C.J. Jackson, T. Parker, D. Faulkner and E. Vidgen, 2002. Effects of high-and low- isoflavone (phytoestrogen) soy foods on inflammatory biomarkers and pro inflammatory cytokines in middle-aged men and women. Metabolism, 51: 919-924.
PubMed - Jones, K.L., J. Harty, M.J. Roeder, T.A. Winters and W.J. Banz, 2005. In vitro effects of soy phytoestrogens on rat L6 skeletal muscle cells. J. Med. Food, 8: 327-331.
PubMed - Kim, S., H.J. Shin, S.Y. Kim, J.H. Kim, Y.S. Lee, D.H. Kim and M.O. Lee, 2004. Genistein enhances expression of genes involved in fatty acid catabolism through activation of PPARalpha. Mol. Cell. Endocrinol., 220: 51-58.
PubMed - Kirk, E.A., P. Sutherland, S.A. Wang, A. Chait and R.C. LeBoeuf, 1998. Dietary isoflavones reduce plasma cholesterol and atherosclerosis in C57BL/6 mice but not LDL receptor-deficient mice. J. Nutr., 128: 954-959.
PubMed - Kishida, T., T. Mizushige, Y. Ohtsu, S. Ishikawa and M. Nagamoto et al., 2008. Dietary soy isoflavone-aglycone lowers food intake in female rats with and without ovariectomy. Obesity, 16: 290-297.
CrossRefPubMedDirect Link - Lazar, M.A., 2005. How obesity causes diabetes: Not a tall tale. Science, 307: 373-375.
CrossRefPubMedDirect Link - Lee, J.S., 2006. Effects of soy protein and genistein on blood glucose, antioxidant enzyme activities and lipid profile in streptozotocin-induced diabetic rats. Life Sci., 79: 1578-1584.
CrossRefPubMedDirect Link - Mezei, O., W.J. Banz, R.W. Steger, M.R. Peluso, T.A. Winters and N. Shay, 2003. Soy isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr., 133: 1238-1343.
PubMed - Mohammad Shahi, M., M.R. Rashidi, S.A. Mahboob, F. Haidari, B. Rashidi and J. Hanaee, 2011. Protective effect of soy protein on collagen-induced arthritis in rat. Rheumatol. Int.
CrossRef - Mullen, E., R.M. Brown, T.F. Osborne and N.F. Shay, 2004. Soy isoflavones affect sterol regulatory element binding proteins (SREBPs) and SREBP-regulated genes in HepG2 cells. J. Nutr., 134: 2942-2947.
PubMed - Naaz, A., S. Yellayi, M.A. Zakroczymski, D. Bunick and D.R. Doerge et al., 2003. The soy isoflavone genistein decreases adipose deposition in mice. Endocrinology, 144: 3315-3320.
CrossRefPubMedDirect Link - Ogden, C.L., M.D. Carroll, L.R. Curtin, M.A. McDowell, C.J. Tabak and K.M. Flegal, 2006. Prevalence of overweight and obesity in the United States, 1999-2004. J. Am. Med. Assoc., 295: 1549-1555.
PubMedDirect Link - Orgaard, A. and L. Jensen, 2008. The effects of soy isoflavones on obesity. Exp. Biol. Med., 233: 1066-1080.
CrossRef - Rayalam, S., M.A. Della-Fera and C.A. Baile, 2008. Phytochemicals and regulation of the adipocyte life cycle. J. Nutr. Biochem., 19: 717-726.
CrossRefPubMedDirect Link - Reinli, K. and G. Block, 1996. Phytoestrogen content of foods-a compendium of literature values. Nutr. Cancer, 26: 123-148.
PubMed - Selvin, E., N.P. Paynter and T.P. Erlinger, 2007. The effect of weight loss on C-reactive protein: A systematic review. Arch. Int. Med., 167: 31-39.
PubMed - Setchell, K.D., L. Zimmer-Nechemias, J. Cai and J.E. Heubi, 1997. Exposure of infants to phyto-oestrogens from soy-based infant formula. Lancet, 350: 23-27.
PubMed - Yildiz, M.F., S. Kumru, A. Godekmerdan and S. Kutlu, 2005. Effects of raloxifene, hormone therapy and soy isoflavone on serum high-sensitive C-reactive protein in postmenopausal women. Int. J. Gynecol. Obstet., 90: 128-133.
PubMed - Zimmermann-Belsing, T. and U. Feldt-Rasmussen, 2004. Obesity: The new worldwide epidemic threat to general health and our complete lack of effective treatment. Endocrinology, 145: 1501-1502.
PubMed