
T.B. Arabani
Department of Marine Biology, Faculty of Marine Science and Technology, Islamic Azad University, North Branch, Tehran, Iran
S. Jamili
Iranian Fisheries Research Organization, P.O. Box 14155-6116, Tehran, Iran
F. Abbassi
Department of Biology, Faculty of Science, Al-Zahra University, Tehran, Iran
Research Journal of Environmental Sciences
Year: 2010 | Volume: 4 | Issue: 5 | Page No.: 473-477







ABSTRACT
The effect of heavy metals on Ca2+-ATPase from the sarcoplasmic reticulum in fish muscles has been studied, but there is not any evidence about interaction between heavy metals and Ca2+ in muscle tissue. The concentration of heavy metals (Zn, Cu, Pb) in two economically important fish species (Ctenopharyngodon idella, Hypophthalmichthys molitrix) collected from some ponds in Gilan, Iran were determined using flame atomic absorption spectrometry after digestion methods. Trace metals content in fish samples were 0.88-4.50 μg g-1 for copper, 1.83-5.92 μg g-1 for lead, 5.63-12.63 μg g-1 for zinc and 72.31-173.43 μg g-1 for calcium. There is no interaction between heavy metals and calcium concentration in muscles.
PDF Abstract XML References Citation
How to cite this article
T.B. Arabani, S. Jamili and F. Abbassi, 2010. Effect of Heavy Metals on Ca2+ Concentration in Muscle Tissue of Grass Carp and Silver Carp. Research Journal of Environmental Sciences, 4: 473-477.
DOI: 10.3923/rjes.2010.473.477
URL: https://scialert.net/abstract/?doi=rjes.2010.473.477
DOI: 10.3923/rjes.2010.473.477
URL: https://scialert.net/abstract/?doi=rjes.2010.473.477
INTRODUCTION
The concern on the effects of anthropogenic pollution of freshwater ecosystems is growing. Heavy metals from natural and anthropogenic sources are continually released into aquatic ecosystems and they are a serious threat because of their toxicity, long persistence, bioaccumulation and biomagnifications in the food chain. Fish samples are considered as one of the most indicative factors, in freshwater systems, for the estimation of trace metals pollution potential (Papagiannis et al., 2004).
Because of complicated kinetics of metals (absorption, distribution, metabolism, elimination, deposition and excretion) and their mutual interactions, it is almost impossible to predict metal effects on tissue concentrations based on their concentrations in the water. Also, the biological availability of an element and its ultimate effect on an organism may depend on a chemical form in which the element is encountered (Kosanovic et al., 2007).
In muscle contraction, Ca2+ is released from sarcoplasmic reticulum into muscle cells via Ca2+-release channel, Ca2+-ATPase then pumps back the released Ca2+ into the SR to cause relaxation. This pump runs as long as ATP and Ca2+ are present in the cytoplasm (Toyoshima, 2008). The Ca2+-ATPase of the Sarcoplasmic Reticulum (SR) is an intrinsic membrane enzyme playing an important role in the muscle contraction of skeletal muscle (Godiksen and Jessen, 2002). Contraction is regulated by Ca2+-sensitive molecular switches on the myosin or actin filaments, depending on the muscle and species. At low Ca2+ levels, actin-myosin interaction is inhibited and actin-activated myosin ATPase is low. At high Ca2+ concentrations, the inhibition of actin–myosin interaction is removed and myosin ATPase is activated by actin (Zhao and Craig, 2003). Voltage-gated Ca2+ channels represent a route of entry into the cell for a wide variety of divalent and trivalent metals (Zalpus and Koropatnick, 2000). A number of metals have been examined and found to have blocking effects and are likely to permeate Ca2+ channels, including Zn (Winegar et al., 1991).
Omnivorous, sediment-dwelling, fish species such as carp may therefore, accumulate heavy metals more readily than pelagic species as a result of exposure to the generally higher metal content of sediment compared with the water column (Alam et al., 2002). Grass carp (C. idella) has received much attention all around the world, primarily due to its potential use for biological control of aquatic vegetation (Vigh et al., 1996). Cd, Cu and Zn contents in edible muscles of pelagic fish species were lower than for benthic fish species. Similarly, the Cd, Cu and Zn contents in muscles of Hypophthalmichthys molitrix (pelagic fish) were lower than those in Cyprinus carpio (benthic fishes). The Pb contents in muscles showed no correlation with fish species (Chi et al., 2007).
In present investigation, the levels of heavy metals and calcium in fish muscles collected from some ponds in Gilan province, Iran, were determined by flame atomic absorption spectrometry after digestion. Then mean concentrations were compared.
MATERIALS AND METHODS
The fish samples (Ctenopharyngodon idella, Hypophthalmichthys molitrix) were collected from three different ponds in Gilan province, Iran in 2002. The collected samples were transferred to the laboratory and washed with distilled water, stored in plastic bags at -20°C until dissection.
Samples (50 g) were weighed and then dried at 105°C for 24 h. Approximately 0.5 g of dried samples were digested in 4 mL nitric acid and after 24 h added 1 mL perchloric acid, followed by heating at a temperature of 60°C for 1 h, then temperature was raised to 120 for 1 h. Samples were then allowed to cool and final solutions were made up to 10 mL with double distilled water (Mason and Barak, 1990). In order to determine Ca2+ in muscle, samples put in muffle furnace set at a temperature of 105°C for 10 h, then ashes were weighted, heated at 505°C until the ash was free from all visible traces of carbon. The ash is treated with 20.0 cm3 (1-1) HCl. The solution is transferred quantitatively to a 50 cm3 volumetric flask. A total of 6.0 cm3 of 25000 ppm lanthanum is added and the resulting solution diluted to the mark with water (Udoh, 2000).
Following acid digestion, all samples were analyzed for elements by flame atomic absorption spectrometry (Shimadzu.AA68V). The operating parameters for working elements were set as recommended by the manufacturer (Table 1). Lead, zinc, copper and calcium were determined in air-acetylene flame. A blank solution is also prepared and analyzed. There are several methods for the determination of metallic elements in biological samples. Atomic absorption spectroscopy is arguably faster, more sensitive and reliable but the determination of calcium poses a problem because of the interference of phosphate ion.
| Table 1: | Instrumental analytical conditions of investigated elements conditions for absorption spectrometry |
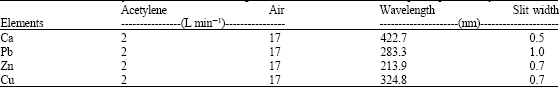 | |
This interference is usually overcome by the addition of lanthanum to the sample solution prior to analysis (Fritze et al., 1987). All digested samples were analyzed three times for each metal.
All reagents were of analytical reagent grade, unless otherwise stated. Double deionised water (Milli-Q Millipore 18.2 MX cm-1 resistivity) was used for all dilutions. HNO3, H2O2 and HCI were of Suprapur quality (E.Merck, Darmstadt, Germany). All the plastic and glassware were cleaned by soaking in dilute HNO3 (1/9, v/v) and were rinsed with distilled water prior to use. The element standard solutions used for calibration were produced by diluting a stock solution of 1000 mg L-1 of the given e lement, supplied by Sigma Chemical Co. (St. Louis, MO). The standard solutions were prepared from Pb (NO3)2, CuSo4, ZnCl2 and CaCO3.
RESULTS AND DISCUSSION
All metal concentrations were determined on a dry weight basis. The levels of zinc, copper, lead and calcium in muscle tissue of grass carp (Ctenopharyngodon idella) and silver carp (Hypophthalmichthys molitrix) are given in Table 2.
Statistical analysis of data was carried out using SPSS statistical package program. According to these data, the high metal accumulation levels in the species were found in C. idella for Cu, Zn and H. molitrix for Pb, respectively. The highest and lowest concentrations of calcium in H. molitrix were found to be 173.43 and 83.35 μg g-1, respectively. While mean concentration of calcium in C. idella ranged from 72.31 to 109.75 μg g-1. Average copper content of H. molitrix samples varied from 0.88 to 2.63 μg g-1. The maximum and minimum copper levels in C. idella were 1.13 and 4.50 μg g-1, respectively. Our copper value was higher than those reported earlier (Vigh et al., 1996). The highest and lowest levels of lead (5.92 and 1.83 μg g-1) were detected in H. moltrix. Our lead value was higher than those reported earlier (Chi et al., 2007). In addition, maximum lead level was higher than those reported from Iranian Fisheries Research Organization (Sadeghi, 1993). Mean zinc concentrations ranged from 5.63-12.63 μg g-1 measured in C. idella. Mea zinc content in samples of H. molitrix was between 6.88-12.13 μg g-1 which was lower than those reported earlier (Chi et al., 2007).
No correlation was reported, however, between metal concentration in the water and fish muscles. Muscle was a poor indicator of low level copper and zinc contamination. That is also true for most other metals, except for mercury which shows higher affinity to the muscles comparing to other metals (Dural et al., 2007).
According to standard deviation in Table 3, there is no significant difference between mean calcium concentrations in silver grass carp and Pb, Zn, Cu accumulation in muscle. In addition, mean difference between calcium and heavy metals in muscle tissue of silver carp were compared, there is no standard deviation (Table 4).
| Table 2: | Maximum and minimum levels of heavy metals (μg/g dw) in C. idella and H. molitrix |
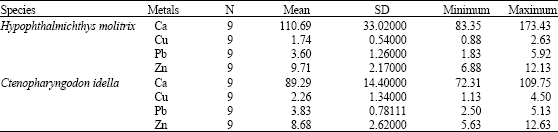 | |
| Table 3: | Comparison of heavy metals and calcium in muscle tissue of grass carp |
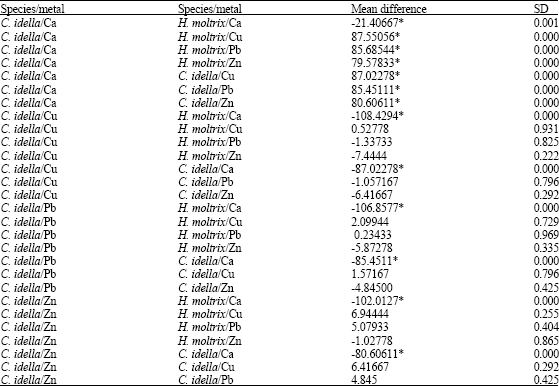 | |
| *Significant difference | |
| Table 4: | Comparison of heavy metals and calcium in muscle tissue of silver carp |
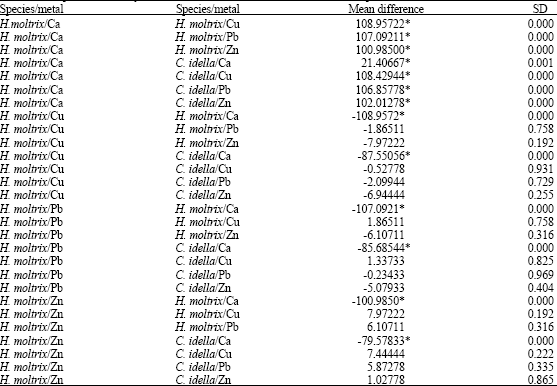 | |
| *Significant difference | |
The Ca-ATPase activity in the muscle decreased due to heavy metals exposure. This may result from the breakdown of the active transport mechanism depending upon the altered membrane permeability and also due to the disturbed Ca homeostasis (Atli and Canli, 2007). There is not any evidence which demonstrate direct interaction between heavy metals and Ca2+ in muscles.
REFERENCES
- Alam, M.G.M., A. Tanaka, G. Allinson, L.J.B. Laurenson, F. Stagnitti and E.T. Snow, 2002. A comparison of trace element concentrations in cultured and wild carp (Cyprinus carpio) of Lake Kasumigaura, Japan. Ecotoxicol. Environ. Saf., 53: 348-354.
CrossRef - Atli, G. and M. Canli, 2007. Enzymatic responses to metal exposures in a freshwater fish Oreochromis niloticus. Comp. Biochem. Physiol. C: Toxicol. Pharmacol., 145: 282-287.
CrossRef - Dural, M., M.Z.L. Goksu and A.A. Ozak, 2007. Investigation of heavy metal levels in economically important fish species captured from the Tuzla Lagoon. Food Chem., 102: 415-421.
CrossRefDirect Link - Godiksen, H. and F. Jessen, 2002. Temperature and Ca2+-dependence of the sarcoplasmic reticulum Ca2+-ATPase in haddock, salmon, rainbow trout and zebra cichlid. Camp. Biochem. Physiol., 133: 35-44.
CrossRef - Kosanovic, M., M.Y. Hasan, D. Subramanian, A.F. Al-Ahbabi and A.O. Al-Kathiri et al., 2007. Influence of urbanization of the western coast of the United Arab Emirates on trace metal content in muscle and liver of wild red-spot emperor (Lethrinus lentjan). Food Chem. Toxicol., 45: 2261-2266.
CrossRef - Mason, G. and N.A.C. Barak, 1990. A catchment survey using the eel (Anguilla anguilla). Chemosphere, 21: 695-699.
CrossRef - Toyoshima, C., 2008. Structural aspects of ion pumping by Ca2+-ATPase of sarcoplasmic reticulum. Arch. Biochem. Biophys., 476: 3-11.
CrossRef - Udoh, A.P., 2000. Atomic absorption spectrometric determination of calcium and other metallic elements in some animal protein sources. Talanta, 52: 749-754.
CrossRef - Vigh, P., Z. Mastalal and K.V. Balogh, 1996. Comparison of heavy metal concentration of grass carp (Ctenopharyngodon idella Cuv. et Val.) in a shallow eutrophic lake and a fish pond (possible effects of food contamination). Chemosphere, 32: 691-701.
CrossRef - Winegar, B.D., R. Kelly and J.B. Lansman, 1991. Block of current through single calcium channels by Fe, Co, and Ni; Location of the transition metal binding site in the pore. J. Gen. Physiol., 97: 351-367.
Direct Link - Zhao, F. and R. Craig, 2003. Ca2+ causes release of myosin heads from the thick filament surface on the milliseconds time scale. J. Molec. Biol., 327: 145-158.
CrossRef - Chi, Q.Q., G.W. Zhu and A. Langdon, 2007. Bioaccumulation of heavy metals in fishes from Taihu Lake, China. J. Environ. Sci., 19: 1500-1504.
CrossRefDirect Link - Papagiannis, I., I. Kagalou, J. Leonardos, D. Petridis and V. Kalfakakou, 2004. Copper and zinc in four freshwater fish species from lake pamvotis (Greece). Environ. Int., 30: 357-362.
CrossRefDirect Link