
Fahri Yigit
Fethiye AS.M.K. Vocational School, University of Mugla, Fethiye, Mugla, Turkey
Murat Dikilitas
Department of Plant Protection, University of Harran, Sanliurfa, Turkey
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 159-163







ABSTRACT
Ability of biocontrol agents, fluorescent Pseudomonas, non-pathogen Fusarium strain and Trichoderma harzianum T-22 applied in combination and alone, to control of Fusarium oxysporum f. sp. lycopersici was studied in the greenhouse. Tomato roots were treated with biomass of flourescens Pseudomonas strain from rhizosphere, non-pathogen Fuasrium sp. and Trichoderma harzianum T-22 individually and in combination and planted in artificially infested soil with pathogen F. oxysporum f. sp. lycopersici. Although all biocontrol agents applied individually reduced disease incidence, treatments as combination, except for nonpathogen Fusarium strain + T. harzianum T-22, showed more protective effect. Combination of T. harzianum T-22 + fluorescent Pseudomonas isolate gave the best control (70.2%).
PDF Abstract XML References
How to cite this article
Fahri Yigit and Murat Dikilitas, 2007. Control of Fusarium Wilt of Tomato by Combination of Fluorescent Pseudomonas, Non-pathogen Fusarium and Trichoderma harzianum T-22 in Greenhouse Conditions. Plant Pathology Journal, 6: 159-163.
DOI: 10.3923/ppj.2007.159.163
URL: https://scialert.net/abstract/?doi=ppj.2007.159.163
DOI: 10.3923/ppj.2007.159.163
URL: https://scialert.net/abstract/?doi=ppj.2007.159.163
INTRODUCTION
Fusarium oxysporum f. sp. lycopersici is a highly destructive pathogen of both greenhouse and field grown tomatoes in warm vegetable production areas. The disease caused by this fungus is characterized by wilted plants, yellowed leaves and minimal or absent crop yield. Many strategies to control this disease on tomato have been investigated in the field. A promising strategy for the replacement of chemicals has been the implementation of biocontrol technology, used individually or as an integrated pest management component. The recent developments in the commercialization of biocontrol products has accelerated this approach (Lumbsden et al., 1996; Fravel et al., 2003). Biocontrol preparations of both fungi and bacteria have been applied to seeds, seedlings, and planting media in several ways to reduce tomato diseases in the field with various degrees of success. Two of the major biocontrol agents which reduce soilborne diseases of various crops include isolates of the bacterium fluorescent Pseudomonas spp. and the fungus Trichoderma spp. (Lumsden and Locke, 1989; Bowers and Parke, 1993; Howell and Stipanovic, 1995. Other antagonist recovered from Fusarium wilt-suppressive soils, especially nonpathogenic F. oxysporum, have been used to reduce Fusarium wilt diseases of several different crops (Minuto et al., 1995; Larkin et al., 1996; Nel et al., 2006). The use of combinations of multiple antagonist organisms also may provide improved disease control over the use of single organisms. Multiple organisms may enhance the level and consistency of control by providing multiple mechanisms of action, a more stable rhizosphere community, and effectiveness over a wider range of environmental conditions. In particular, combinations of fungi and bacteria may provide protection at different times or under different conditions, and occupy different or complementary niches. Such combinations may overcome inconsistencies in the performance of individual isolates. The objectives of this research were to evaluate the combined and alone effect of three biocontrol agents, T. harzianum T-22, nonpathogen Fuarium and flourescent Pseudomonas on Fusarium wilt of tomato in the greenhouse.
MATERIALS AND METHODS
Microbial cultures: The pathogenic fungi used in this study included isolate of Fusarium oxysporum f. sp. lycopersici Snyder and Hansen. Biocontrol agents studied were the fungus Trichodenna harzianum T-22 and the bacterium fluorescent Pseudomonas isolate. The pathogenic fungi were provided by Demir, S., Van Y.Y. University, Agriculture Faculty, Department of Plant Protection. The antagonistic bacterium and non-pathogen Fusarium were isolated from tomato rhizosphere during the before preliminary study. Flourescent Pseudomonas isolates were screened antagonistic activity against each fungi and selected isolate showing antagonistic activity against pathogen tested, not to T. harzianum. T. harzianum strain T-22 was isolated from commercial biopreparat (T-22 Planter Box) and proliferated on Potato Dextrose Agar medium.
Preparation of inocula: F. oxysporum f. sp. lycopersici and T. harzianum T-22 were cultured in moistened wheat bran-corn mill (l: l, w/w) medium in flats for three weeks at 25°C. After incubation, the each inoculum was air-dried for three days, milled in a blender and sieved through a 3.36 mm screen and stored at 4°C (Mao et al., 1997). Inoculum viability of each fungi was determined by serial dilution on a peptone pentachloronitrobenzene (PCNB) medium for pathogen and water agar for T. harzianum. There were approximately 1.6xl06 and 1.8x107 colony forming units (cfu) per gram of inoculum of F. oxysporum f. sp. lycopercisi and T. harzianum T-22, respectively. The fluorescent Pseudomonas isolate was grown on 9 cm diameter petri plates containing 10 mL King-B. Plates were incubated in the dark for one week at 25 + 2°C . when the antagonistic bacterium would be inoculated to plant roots, culture grown on the medium were scraped from the surface of the plates and adjusted 1x109 cfu mL-1 by serial dilution.
Greenhouse tests: For root dipping, each biomass, alone and in combination were prepared separately in different container containing an uncentrifuged bacterial suspension (1.8x107) and 100.0 g L-1 of both biocontrol fungi biomass except for pathogen Fusarium. Before the transplanting, roots of transplants were dipped into each biomass and then transplanted to greenhouse soil artificially infested with pathogen. Four control rows were planted with untreated tomato transplants. Greenhouse soil was artificially infested with pathogen fungi grown on moistened wheat bran-corn mill at rate of 100 g m-2 soil. Each treatment consisted of four replicate rows of ten plants/row. Disease was monitored for 6 to 8 weeks and assayed as the total percentage of plants showing any wilt symptoms due to the pathogen (yellowing and dropping of leaves, vascular discoloration, wilting). Stem sections of wilted plants were surface-disinfested in 0.5% sodium hypochlorite and plated on PCNB medium to confirm the presence of the wilt pathogen. Stem sections of asymptomatic plants were also plated at the conclusion of the experiment to evaluate potential pathogen infection. Experiment were conducted in Fethiye county of Mugla province in 2005-2006 growing season.
All greenhouse experiments were performed twice with four replicates per treatment and arranged in a randomized complete block design. Disease incidence (%) were analyzed using an analysis of variance (ANOVA) and grouped by DUNCAN test.
RESULTS AND DISCUSSION
Individual and combination of biocontrol organisms tested significantly reduced Fusarium wilt of tomato in greenhouse tests (Table 1). Reductions in disease incidence ranged from 30 to 70.2% relative to control. The most effective combinations were non-pathogen Fusarium with fluorescent Pseudomonas sp and T. harzianum T-22 with fluorescent Pseudomonas sp and reduced disease incidence by 70.2 and 60.8%, respectively. However, the level of disease control provided by non-pathogen Fusarium + T. harzianum T-22 was not as good as than provided by the individual biocontrol organisms tested. When compared to individual effect, non-pathogen Fusarium and fluorescent Pseudomonas sp. were more affective than T. harzianum T-22. These results showed that non-pathogen Fusarium isolates would be important in reducing Fusarium wilt disease. Larkin and Fravel (1998) reported that specific nonpathogenic isolates of Fusarium oxysporum and F. solani collected from a Fusarium-wilt suppressive soil were the most effective antagonists and provided significant and consistent disease control (50 to 80% reduction of diseases incidence) in several repeated tests. In addition, isolates of Gliocladium virens, T. hamatum, Pseudomonas fluorescens and Burkholderia cepacia, also significantly reduced Fusarium wilt compared to disease controls (30 to 65% reduction), but were not as consistently effective as the nonpathogenic Fusarium isolates.
The most important result from our research was the improved biocontrol performance when nonpathogen Fusarium isolate was used in combination with a fluorescent Pseudomonas strain. Park et al. (1988) and Leeman et al. (1995, 1996) have also reported a similar enhanced response with combinations of Fusarium and Pseudomonas spp. in cucumber and radish. In addition, Fuchs et al. (1999) and Alabouvette et al. (1993) reported improved biocontrol performance when Fusarium isolate Fo47 was used in combination with a fluorescent Pseudomonas strain (C7). The nonpathogenic F. oxysporum strains show several modes of action contributing to their biocontrol capacity. They are able to compete for nutrients in the soil, affecting the rate of chlamydospore germination of the pathogen.
| Table 1: | Development of Fusarium wilt in tomato plants as affected by treatment with various combinations of biocontrol organisms and alone |
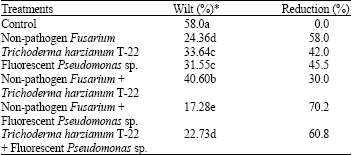 | |
| *Values followed by different letters within a column differ significantly, p<0.05 | |
They can also compete for infection sites on the root and can trigger plant defense reactions, inducing systemic resistance (Benhamou et al., 2002). The competitive ability of a nonpathogenic strain partly determines its capacity to establish in soil and in the plant rhizosphere and is probably involved in its capability to colonize the root surface. Nagao et al. (1990) demonstrated that different strains have different capacities to colonize heat treated soil. In addition, Saprophytic colonisation of soil depends not only on the fungal strain but also on biotic and abiotic soil characteristics. Colonisation of the root surface and root tissues probably depends not only on the fungal strain but also on the plant species and plant cultivar.
Benhamou and Garand (2001) determined enzymatic changes in the plant inoculated with nonpathogen Fusarium, often leading to induction of the physical barriers. Tamietti et al. (1993) found increased activity of several plant enzymes related to plant defense reactions in tomato plants transplanted in sterilized soil infested with a strain of nonpathogenic F. oxysporum. Fuchs et al. (1997) attributed the biocontrol activity of the nonpathogenic strain Fo47 to induced resistance in tomato, correlated with an increased activity of chitinase, 1-3 glucanase and 1-4 glucosidase. Duijff et al. (1998) showed that the nonpathogenic strain Fo47 although not very effective in inducing systemic resistance in tomato induced an increase of PR proteins. Recorbet et al. (1998) showed an overall increased activity of constitutive glycosidase isoforms in response to infection by F. o. f. sp. lycopersici that did not occur in roots colonized with nonpathogenic strains. Finally, when the main mode of action of a nonpathogenic strain is induction of systemic resistance, it is obvious that this phenomenon implies the physiological state of the plant and fluctuating environmental conditions may affect the ability of the plant to express its resistance to the pathogen, induced by the nonpathogenic F. oxysporum. The other mechanism involves competition for carbon provided by both F. oxysporum and fluorescent Pseudomonas and competition for iron provided by the bacteria. Fluorescent Pseudomonas strains are an important components of the rhizosphere and exclusive the pathogens from rhizosphere via several factors, such as toxins, antibiotics and siderophores (Thomashow and Weller, 1996). Lemanceau et al. (1992, 1993) suggested that the presence of the siderophore produced by P. fluorescens increases the intensity of the antagonism provided by Fo47, making the pathogen more sensitive to glucose competition by the antagonistic Fusarium. However, in our experiment, it was not search this response. Duijff et al. (1998) reported that the suppression of Fusarium wilt by P. fluorescens WCS417 was due to systemic induced resistance without any detection of the pathogenesis-related proteins tested. In contrast, the suppression achieved by nonpathogenic F. oxysporum Fo47 appeared to be mainly ascribed to microbial antagonism but also to a lesser extent to systemic induced resistance. Duijff et al. (1998) reported that suppression of Fusarium wilt by P. luorescent WCS417 was due to systemic induced resistance. In present sturdy, it was thought that the reason of highly suppressive effect of nonpathogen Fusarium and fluorescens Pseudomonas in combination could be due to synergist effects of all biocontrol mechanisms.
Trichoderma species are among the most-promising biocontrol fungi against many fungal plant pathogens. T. harzianum has multiple mechanisms of action, including mycoparasitism via production of chitinases, β-1-3 glucanases and β-1-4 glucanases, antibiotics, competition, solubilization of inorganic plant nutrients, induced resistance and inactivation of the pathogen's enzymes involved in the infection process (Elat et al., 1982; Sivan and Chet, 1989; Lorito et al., 1996; Bailey and Lumsden, 1998; Altomare et al., 1999; Elad and Kapat, 1999, Hanson and Howell, 2004, Harman, 2006). We suggested that nonpathogen Fusarium would be inhibited by these mechanisms mentioned above. Due to these possible antagonistic interactions, combination of nonpathogen Fusarium isolate and T. harzianum T-22 provided lower disease control than by individual nonpathogen Fusarium.
As a result, further research on the potential advantages of using combinations of these effective antagonists is needed. Although only a limited number of potential biocontrol isolates could be tested in this study, it was obtained satisfactory results. In many studies, many nonpathogen Fusarium strains showed antagonistic potential as biological control agents for the control of Fusarium wilt diseases.
REFERENCES
- Alabouvette, C., P. Lemanceau and C. Steinberg, 1993. Recent advances in the biological control of fusarium wilts. Pestic. Sci., 37: 365-373.
CrossRefDirect Link - Altomare, C., W.A. Norvell, T. Bjbrkman and G.E. Harman, 1999. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Applied Environ. Microbiol., 65: 2926-2933.
Direct Link - Benhamou, N. and C. Garand, 2001. Cytological analysis of defense-related mechanisms induced in pea root tissues in response to colonization by nonpathogenic Fusarium oxysporum Fo47. Phytopathology, 91: 730-740.
CrossRefDirect Link - Benhamou, N., C. Garand and A. Goulet, 2002. Ability of nonpathogenic Fusarium oxysporum strain Fo47 to induce resistance against Pythium ultimum infection in cucumber. Applied Environ. Microbiol., 68: 4044-4060.
Direct Link - Bowers, J.H. and J.L. Parke, 1993. Epidemiology of Pythium damping-off and Aphanomyces root rot of peas after seed treatment with bacterial agents for biological control. Phytoputhol., 83: 1466-1473.
Direct Link - Duijff, B.J., D. Pouhair, C. Olivain, C. Alabouvette and P. Lemanceau, 1998. Implication of systemic induced resistance in the suppression of Fusarium wilt of tomato by Pseudomonas fluorescens WCS417r and by non-pathogenic Fusarium oxysporum Fo47. Eur. J. Plant. Pathol., 104: 903-910.
Direct Link - Elad, Y., I. Chet and Y. Henis, 1982. Degradation of plant pathogenic fungi by Trichoderma harzianum. Can. J. Microbiol., 28: 719-725.
CrossRefDirect Link - Fravel, D., C. Olivain and C. Alabouvette, 2003. Fusarium oxysporum and its biocontrol. New Phytol., 157: 493-502.
CrossRefDirect Link - Fuchs, J.G., Y. Moenne-Loccoz and G. Defago, 1997. Nonpathogenic Fusarium oxysporum strain Fo47 induces resistance to Fusarium wilt in tomato. Plant Dis., 81: 492-496.
CrossRefDirect Link - Fuch, J.G., Y. Moenne-Loccoz and G. Defago, 1999. Ability of nonpathogenic Fusarium oxysporum strain Fo47 to protect tomato against Fusarium wilts. Biol. Control, 14: 105-110.
CrossRefDirect Link - Hanson, L.E. and C.R. Howell, 2004. Elicitors of plant defense responses from biocontrol strains of Trichoderma virens. Phytopathol., 94: 171-176.
Direct Link - Harman, G.E., 2006. Overview of mechanisms and uses of Trichoderma spp. Phytopathology, 96: 190-194.
CrossRefDirect Link - Larkin, R.P. and D.R. Fravel, 1998. Efficacy of various fungal and bacterial biocontrol organisms for control of Fusarium wilt of tomato. Plant Dis., 82: 1022-1028.
CrossRefDirect Link - Larkin, R.P., D.L. Hopkins and F.N. Martin, 1996. Suppression of Fusarium wilt of watermelon by nonpathogenic Fusarium oxysporum and other microorganisms recovered from a disease-suppressive soil. Phytopathology, 86: 812-819.
Direct Link - Leeman, M., F.M. Den Ouden, J.A. van Pelt, M.J. Hendrickx, R. Scheffer, P.A.H.M. Bakker and B. Schippers, 1995. Biocontrol of fusarium wilt of radish in commercial greenhouse trials by seed treatment with Pseudomonas fluorescens WCS374. Phytopathology, 85: 1301-1305.
Direct Link - Lemanceau, P., P.A. Bakker, W.J. De Kogel, C. Alabouvette and B. Schippers, 1992. Effect of pseudobactin 358 production by Pseudomonas putida WCS358 on suppression of fusarium wilt of carnations by nonpathogenic Fusarium oxysporum Fo47. Applied Environ. Microbiol., 58: 2978-2982.
PubMedDirect Link - Lemanceau, P., P.A.H.M. Bakker, W.J. De Kogel, C. Alabouvette and B. Schippers, 1993. Antagonistic effect on nonpathogenic Fusarium oxysporum strain Fo47 and pseudobactin 358 upon pathogenic Fusarium oxysporum f. sp. dianthi. Applied. Environ. Microbiol., 59: 74-82.
Direct Link - Lorito, M., S.L. Woo, M. Dambrosio, G.E. Harman, C.K. Haves C.P. Kubicek and F. Scala, 1996. Synergistic interactions between cell wall degrading enzymes and membrane affecting compounds. Mol. Plant-Microbe Interact., 9: 206-213.
Direct Link - Lumsden, R.D. and J.C. Locke, 1989. Biological control of damping-off caused by Pythium ultimum and Rhizoctonia solani with Gliocladium virens in soilless mix. Phytopathology, 79: 361-366.
Direct Link - Lumsden, R.D., J.F. Walter and C. Baker, 1996. Development of Gliocladium virens for damping-off disease control. Can. J. Plant Pathol., l8: 463-468.
Direct Link - Mao, W., J.A. Lewis, P.K. Hebbar and R.D. Lumsden, 1997. Seed treatment with a fungal or a bacterial antagonist for reducing corn damping-off caused by species of Pythium and Fusarium. Plant Dis., 81: 450-454.
CrossRefDirect Link - Nel, B., C. Steinberg, N. Labuschagne and A. Vilioen, 2006. The potential of nonpathogenic Fusarium oxysporum and other biological control organisms for suppressing fusarium wilt of banana. Plant Pathol., 55: 217-223.
Direct Link - Recorbet, G., G. Bestel-Corre, E. Dumas-Gaudot, S. Gianinazzi and C. Alabouvette, 1998. Differential accumulation of b-1,3-glucanase and chitinase isoforms in tomato roots in response to colonization by either pathogenic or non-pathogenic strains of Fusarium oxysporum. Microbiol. Res., 153: 257-263.
Direct Link - Sivan, A. and I. Chet, 1989. The possible role of competition between Trichoderma harzianum and Fusarium oxysporum on rhizosphere colonization. Phytopatholgy, 79: 198-203.
Direct Link - Tamietti, G., L. Ferraris, A. Matta and I.A. Gentile, 1993. Physiological responses of tomato plants grown in fusarium suppressive soil. J. Phytopathol., 138: 66-76.
Direct Link