
S. Khammuang
Protein and Enzyme Technology Research Unit, Department of Chemistry, Faculty of Science, Mahasarakham University, T. Khamriang, A. Kantarawichai, Maha Sarakham, 44150 Thailand
R. Sarnthima
Protein and Enzyme Technology Research Unit, Department of Chemistry, Faculty of Science, Mahasarakham University, T. Khamriang, A. Kantarawichai, Maha Sarakham, 44150 Thailand
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 8 | Page No.: 616-623







ABSTRACT
This study reported the decolorization of hazardous xanthenes dye, Rhodamine B by the Laccase Mediator System (LMS). Seven redox mediators were investigated in the mediators-assisted lacase catalyze oxidation reactions. Among redox mediators tested, 2, 2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) was the best one which gave the highest Rhodamine B decolorization for more than 80% within 48 h while only 20% achieved with no mediator added. In the laccase-ABTS mediator system, the best molar ratio of dye/mediator was 1:10 and dye/enzyme ratio was 0.5 μmol U-1. The optimum conditions for Rhodamine B decolorization were at pH 4.0-5.0 and temperature 35-40°C. The laccase-ABTS system could be a promising biotechnology developed for treatment of textile waste waters containing Rhodamine B.
PDF Abstract XML References Citation
How to cite this article
S. Khammuang and R. Sarnthima, 2009. Mediator-Assisted Rhodamine B Decolorization by Tramates versicolor Laccase. Pakistan Journal of Biological Sciences, 12: 616-623.
DOI: 10.3923/pjbs.2009.616.623
URL: https://scialert.net/abstract/?doi=pjbs.2009.616.623
DOI: 10.3923/pjbs.2009.616.623
URL: https://scialert.net/abstract/?doi=pjbs.2009.616.623
INTRODUCTION
Synthetic dyes have been used worldwide in textile industries because of simply uses, low cost, resistance to light, heat and microorganisms as well as various color shades availability comparison to dyes from natural sources. Dye effluents releasing from those industries and daily uses contain complex mixtures of several toxic substances. These dye effluents are difficult to treat and nowadays environment control agency in most countries consider setting up law to control the dye amounts in waste effluent.
Dyes removal from wastewater can be performed by various methods such as co-precipitation, adsorption and chemical oxidation. However, these methods have some disadvantages for example, large amount chemicals uses or waste sludge formation; complicate operation as well as high cost process (Nigam et al., 2000). One interesting approach for dye decolorization is enzymatic treatment. This method is more environmental friendly and is a promising method appropriate for near future green industries.
Laccases (benzendiol: oxygen oxidoreductase, EC 1.10.3.2) are belong to polyphenol oxidase, catalyze the reduction of dioxygen to water while oxidize its substrates (Baldrian, 2006). This enzyme firstly found in Japanese lacquer tree, Rhus vernicifera and not long after that laccases from fungi has been reported, especially in white-rot fungi. With this source laccases are one of enzymes with high amount of literatures could be observed. Laccases are secreted as one of ligninolytic enzymes from white rot fungi; others are lignin peroxidases (LiP; EC 1.11.1.14) and manganese peroxidases (MnP; EC 1.11.1.13). From the irregular structure of lignin render these enzymes can catalyze the reactions with broad substrates, many dangerous substances including synthetic dyes. However, the reactions catalyze by LiP or MnP need hydrogen peroxide as co-substrate, this substance is not necessary for laccases catalysis. These unique properties made laccases to be promising enzymes for bioremediation. Alternatively, enzymatic oxidation of synthetic recalcitrant dyes using laccases has received great attention in recent years due to the efficient decolorization of various dye effluents (Kandelbauer et al., 2004; Murugesan et al., 2007).
However, many substrates of laccases are not oxidized due to the enzymes have relatively low redox potentials. Laccases efficiency could be enhanced when the low molecular weight compounds are included as redox mediators. This strategy has proven effectively in several reports by Murugesan et al. (2007), Chhabra et al. (2008) and Moldes and Vidal (2008).
Enzymatic catalyze the oxidation of synthetic dyes have several advantages over the conventional physico-chemical methods. This is because of mild conditions and environmental friendly methods have been used. Enzymatic decolorization of a xanthenes dye, Rhodamine B by LiP has been reported by Lan et al. (2006). There has no report described about this dye decolorization by laccase. Therefore, this research aims to investigate the decolorization of Rhodamine B by laccase and Laccase-Mediator System (LMS). In addition, the optimization of LMS condition including type and concentration of redox mediators, type and pH of buffers, dye/enzyme ratios as well as temperature effects, were also investigated.
MATERIALS AND METHODS
Laccase activity assay: The laccase (EC 1.10.3.2) from Trametes versicolor was purchased from Fluka (specific activity 0.53 U mg-1) and was used without further purification. The enzyme solution was prepared in 0.1 M sodium acetate buffer (pH 4.5). The enzyme assay was based on the oxidation of ABTS as previously described by Khammuang and Sarnthima (2007).
Protein determination: Soluble proteins in the enzyme solution were determined by the Bradford (1976) method using Bio-Rad Protein Assay Reagent (Bio-Rad) with bovine serum albumin as a protein standard.
In vitro Rhodamine B decolorization
Effect of mediator types: Reaction mixtures (2 mL) contained 0.02 U mL-1 laccase activity, 0.01 mM Rhodamine B (0.239 g L-1) and without or with 0.05 mM mediators (ABTS, vanillin, vanillic acid, 4-nitrophenol, veratyl alcohol, 1-naphthol and 2-naphthol) in 0.1 M sodium acetate buffer (pH 4.5). Reactions were incubated at 32°C for 48 h. For Rhodamine B decolorization, the decrease in absorbance at the maximum absorption wavelength (555 nm) was used to calculate decolorization percentage according to the following equation as previously described by Khammuang and Sarnthima (2007).
where, A0 is an absorbance at 555 nm immediately measured after adding the enzyme solution and At is an absorbance at 555 nm after each time intervals.
Effect of mediator concentration: The mediator ABTS was tested at various concentrations (0, 0.01, 0.05 and 0.10 mM) for decolorization of solution of Rhodamine B dye. Reaction mixture (5 mL) contained 0.1 M sodium acetate buffer (pH 4.5) and 0.01 mM Rhodamine B (0.239 g L-1) in the absence or presence of mediator. Reactions were initiated by the addition of the enzyme solution (0.02 U mL-1). The solutions were incubated at 32°C for 7 days. Aliquots were daily withdrawn for dye decolorization assessment.
Effect of pH: Reaction mixtures (2 mL) contained 0.01 mM Rhodamine B dye, 0.10 mM ABTS and the enzyme solution (0.02 U mL-1) in various pH buffers including 0.1 M sodium acetate buffer (pH 3.6, 4.0, 4.5, 5.0 and 5.5) or 0.1 M citrate-phosphate buffer (pH 3.0, 4.0, 5.0, 6.0, 7.0, 8.0). Reactions were incubated at 32°C for 24 h.
Effect of enzyme concentration: Reaction mixture (2 mL) contained 0.01 mM Rhodamine B, 0.10 mM ABTS in 0.1 M sodium acetate buffer (pH 4.5) with various enzyme concentrations (0.01, 0.02, 0.03, 0.04 and 0.05 U mL-1). Reactions were incubated at 32°C for 24 h.
Effect of dye concentration: Reaction mixtures (2 mL) were done at 32°C for 24 h in 0.1 M sodium acetate buffer (pH 4.5) containing the laccase activity 0.02 U mL-1 and various dye concentrations (0.01, 0.02, 0.03, 0.04 and 0.05 mM) in the presence of 0.10 mM ABTS.
Effect of temperature: Reaction mixture contained the laccase activity 0.02 U mL-1, 0.01 mM Rhodamine B and 0.10 mM ABTS in 0.1 M sodium acetate buffer (pH 4.5). Reaction mixtures (2 mL) were investigated at various temperatures (30, 35, 40, 45, 50, 60 and 70°C) for 24 h.
Absorption spectra: Reaction mixtures (6 mL) were investigated at various times (0, 18, 24, 48 and 72 h) at optimum temperature. Reaction mixture contained the laccase activity 0.02 U mL-1, 0.01 mM Rhodamine B and 0.10 mM ABTS in 0.1 M sodium acetate buffer (pH 4.5). Aliquots at various interval times were scanned for absorption spectra at wavelength from 400 to 800 nm by a UV-visible spectrophotometer (Perkin Elmer, Lamda 25, USA). Control was a reaction mixture without the Rhodamine B.
Rhodamine B decolorization at a higher concentration of naphtha compounds: The system with 1-naphthol and 2-naphthol as redox mediators at higher concentrations were performed using 0.5 mM 1-naphthol or 2-naphthol in 0.1 M sodium acetate buffer (pH 4.5) containing 0.01 mM Rhodamine B, 0.02 U mL-1 laccase at 32°C for 24 h.
All reported data were averaged from duplicate experiments. All experiments were performed in the Protein and Enzyme Technology Research Unit, Faculty of Science, Mahasarakham University, during June 2007 to January 2009.
RESULTS
Effect of mediator types on decolorization of Rhodamine B: In this experiment, the decolorization of hazardous xanthenes dye, Rhodamine B was investigated in the presence of seven redox mediators including synthetic compounds (ABTS, 4-aminophenol, veratryl alcohol, 1-naphthol and 2-naphthol) and natural ones (vanillin, vanillic acid) (Fig. 1a-h) in comparison with the absence mediator condition.
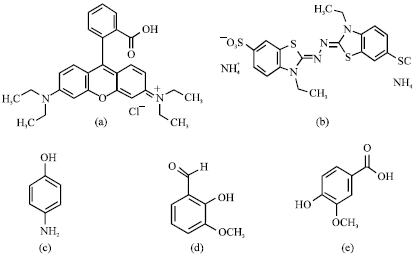 | |
| Fig. 1: | Chemical structures of Rhodamine B dye and 7 redox mediators. (a) Rhodamine B, (b) ABTS, (c) 4-aminophenol, (d)vanillin, (e) 4-hydroxy-3-methoxybenzoic acid (vanillic acid), (f) 3,4-dimethoxyphenyl methanol (veratryl alcohol), (g) 1-naphthol and (h) 2-naphthol |
The results showed that in the absence of redox mediator, Rhodamine B was decolorized only about 19% within 48 h as shown in Fig. 2. In the presence of certain redox mediators, decolorization efficiency of the dye was increased, but in some cases became worst. Vanillin and vanillic acid had no effect on decolorization of the dye (~17% decolorization), whereas 4-aminophenol and veratryl alcohol decreased decolorization ability of the laccase enzyme (<10% decolorization). Among mediators tested, one of synthetic mediator ABTS gave the highest percentage of Rhodamine B decolorization (80% decolorization).
Effect of mediator concentration: Due to synthetic mediators ABTS showed the most promising result, it was chosen for further study the effect of mediator concentration on the dye decolorization. The decolorization of Rhodamine B was strongly affected by the ABTS concentration and time-dependent increases in decolorization percentages (Fig. 3). Higher concentration of mediator provided higher decolorization percentages. At 24 h of incubation, in the absence of mediator the dye was decolorized only 14.7%, whereas in the presence of 0.01, 0.05 and 0.10 mM ABTS the dye was decolorized for 31.1, 54.7 and 84.3%, respectively.
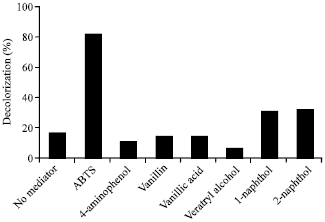 | |
| Fig. 2: | Effect of redox mediator types on decolorization percentage of Rhodamine B (0.01 mM) by laccase from Trametes versicolor (0.02 U mL-1) in 0.1 M sodium acetate buffer, pH 4.5 at 32°C for 48 h in the presence or absence of redox mediator (0.05 mM) |
The decolorization percentage was increased differently in latter days. In case where no mediator presence in the reaction, the decolorization percentage was slightly increased to 19.0% in day 2 and then tended to constant decolorization observed.
In case where 0.01 mM ABTS presence, the decolorization ability appeared to increase enormously before reached to its maximum value around 80% at day 5. In the presence of 0.05 mM ABTS, Rhodamine B was degraded above 80% within 2 days and then slightly increased and reached the plateau above 90% at day 4. At the highest ABTS concentration tested (0.10 mM), the reaction was reached the equilibrium shortest within 2 days and provided the highest decolorization percentage of Rhodamine B about 92% (Fig. 3).
Effect of pH: Type of buffers and pH values were also affected on decolorization of Rhodamine B by the laccase. As shown in Fig. 4, the reactions in sodium acetate buffer showed high decolorization percentages around pH 4.0-5.5 (above 80% decolorization) whereas, the reactions in citrate-phosphate buffer showed lower efficiency in the same pH range (~50-60% decolorization).
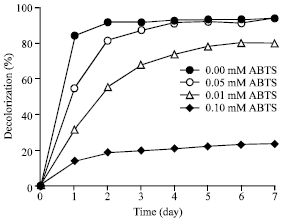 | |
| Fig. 3: | Decolorization percentage of Rhodamine B (0.01 mM) by laccase from Trametes versicolor (0.02 U mL-1) in 0.1 M sodium acetate buffer, pH 4.5 at 32°C in the absence or presence of various concentrations of ABTS mediator |
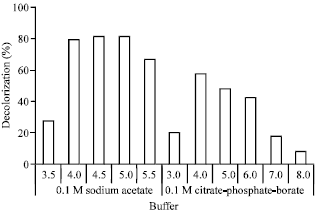 | |
| Fig. 4: | Effect of pH types and values on decolorization of Rhodamine B by laccase from Trametes versicolor (0.02 U mL-1). The reactions contained 0.10 mM ABTS mediator were experimented at 32°C for 24 h using 0.1 M sodium acetate buffer and 0.1 M citrate-phosphate-borate buffer |
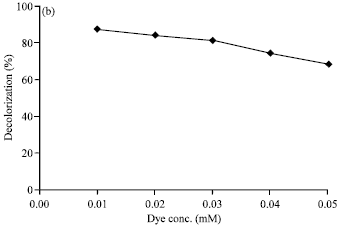
Effect of enzyme and dye concentration: Enzyme and dye concentrations were found to have a slightly effect on decolorization of Rhodamine B by the laccase from T. versicolor. The results revealed that higher laccase activity gave higher decolorization percentages and the higher concentration of the dye, the lower the decolorization ability. The best proportion of dye and enzyme which yield the highest percentage in the dye decolorization, were 0.01 mM Rhodamine B and 0.02 U mL-1 laccase activity (or 0.5 μmol U-1) in the presence of 0.10 mM ABTS mediator. The results were shown in Fig. 5a and b.
Effect of temperature: Thermal effect was also tested on decolorization of Rhodamine B by the laccase. As the results showed in Fig. 6, at 30°C, within 24 h dye was decolorized only about 50%.
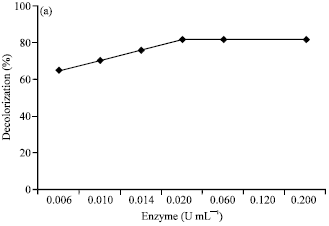 | |
| Fig. 5: | Effect of (a) enzyme concentration and (b) dye concentration on decolorization of Rhodamine B by laccase from Trametes versicolor. The reactions contained 0.01 mM Rhodamine B in 0.1 M sodium acetate buffer (pH 4.5) at 32°C for 24 h in the presence of 0.10 mM ABTS mediator |
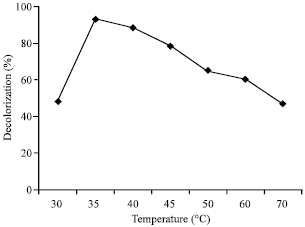 | |
| Fig. 6: | Effect of temperature on decolorization of Rhodamine B by laccase from Trametes versicolor. The reactions contained 0.01 mM Rhodamine B, 0.02 U mL-1 laccase in 0.1 M sodium acetate buffer, pH 4.5 at various temperatures for 24 h in the presence of 0.10 mM ABTS mediator |
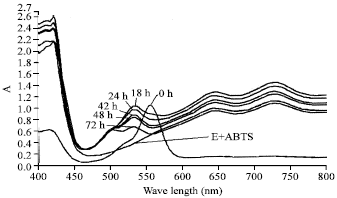 | |
| Fig. 7: | Absorption spectral changes of Rhodamine B (0.01 mM) treated with laccase from Trametes versicolor (0.02 U mL-1) in 0.1 M sodium acetate buffer, pH 4.5 at 32°C at various incubation times in the presence of ABTS mediator (0.1 mM). Data were from a UV-visible spectrophotometer measurements |
Higher temperature of reaction raised the decolorization yield. The most optimum temperature was at 35°C, above this temperature the decolorization percentages seemed to slightly decrease. At 70°C, the decolorization of dye was around 50% after 24 h of incubation.
Absorption spectra: Absorption spectra obtained for the Rhodamine B dyes incubated with laccase from T. versicolor in 0.1 M sodium acetate buffer, pH 4.5 at time 0, 18, 24, 48 and 72 h of incubation were as shown in Fig. 7.
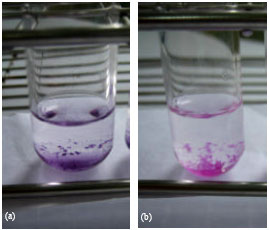 | |
| Fig. 8: | Co-precipitation properties of (a) 1-naphthol and (b) 2-naphthol in decolorization of Rhodamine B by laccase from Trametes versicolor. The reactions contained 0.01 mM Rhodamine B, 0.02 U mL-1 laccase in 0.1 M sodium acetate buffer (pH 4.5) at 32°C for 24 h in the presence of 0.5 mM 1-naphthol and 2-naphthol |
The results showed a spectrum of decolorization products shift to the shorter wavelength 540 and then 500 nm.
Rhodamine B decolorization at a higher concentration of naphtha compounds: The system with 1-naphthol and 2-naphthol as redox mediators at higher concentrations revealed interesting results. As shown in Fig. 8a and b, apart from decolorization of the dye, the precipitation products appeared and remained the clear solution. In the presence of 1-naphthol, purple precipitate occurred whereas pink precipitate in the case of 2-naphthol compound.
DISCUSSION
The basis of the laccase-mediator concept is the use of low molecular weight compounds those, once oxidized by the enzyme to stable radicals, act as redox mediators, oxidizing other compounds that in principle are not substrates of laccases. Mediator types had a strong effect on decolorization of Rhodamine B by laccase from T. versicolor. Among mediators tested, synthetic mediator ABTS gave the highest percentage of the dye decolorization. It is possible that because of ABTS which has been reported as one of laccase’s substrate with a low km, could bind with the enzyme better than other compounds then be converted to cation radical (ABTS+) and thus catalyzed the oxidation of Rhodamine dye (Bourbonnais and Paice, 1990).
Present result with ABTS, a synthetic substance, as the reasonable good mediator is in agreement with a number of reports. It enhanced decolorization of sella solid red and luganil green (Rodríguez et al., 2005), naphtol blue black (Svobodová et al., 2008) as well as a number of reactive and acidic dyes (Chhabra et al., 2008). There have been some reports about phenolic compound could act as enzyme redox mediators. Phenolic compound mediators, such as phenol compounds syringaldehyde, acetosyringone, whose mediating mechanism was same as -NOH- type mediators (such as 1-hydroxybenzotriazole, HBT) and except that intermediate was a phenoxy radical (instead of NO·− radicals) (Minussi et al., 2007). However, 4-aminophenol, veratryl alcohol, vanillin and vanillic acid were not found to be effective for decolorization of Rhodamine B by T. versicolor laccase. These phenolic compounds seemed not be effective mediators in agreement to the report of Camarero et al. (2005). The 4-aminophenol and 1-naphthol could not promote decolorization of direct dyes salt fractionated turnip (Brassica rapa) proteins which reported by Matto and Husain (2007). In present study, 4-aminophenol gave the adverse effect whereas 1-naphthol could increase decolorization percentages about 2 folds comparison to no mediator use. In the same report, vanillin could be a reasonable good mediator for direct dyes decolorization, however, it could not be effective in Rhodamine B decolorization by T. versicolor laccase in this report. In conclusion, present results went into the same ways with several reports that ABTS was one of the mediators of choice.
The results suggested that the effective ABTS as a redox mediator for the dye decolorization much depended on its concentration in the reaction and time-dependent manner. This finding is in agreement with the results reported by Chhabra et al. (2008) in the decolorization of reactive black 5 and acid violet 17 as well as in the report of Camarero et al. (2005).
Only about 10% increase in sella solid red decolorization when ABTS used as mediator in a very low concentration such 0.1 or 0.5 μM (Rodrνguez et al., 2005). In present result, the optimum decolorization was achieved at 0.10 mM ABTS mediator for Rhodamine B decolorization in the same concentration with the decolorization of reactive and acidic dyes reported by Chhabra et al. (2008). However, in their study with those dyes the dye/mediator molar ratio was 1:2 (whereas 1:10 in present study) which is more cost effective. Some reports used ABTS as mediator in dye decolorization at a higher concentration than that being use in present study. For example, in the report of Svobodová et al. (2008) used 0.2 mM ABTS to enhanced decolorization of naphtol blue black by the mycelium-associated laccase from Irpex lacteus. However, in the same report, in the presence of ABTS, the decolorization of RBBR was reduced. These imply that mediator-laccase system for dye decolorization ability is different among dye types and sources of laccases as well as mediator types.
Several reports revealed an acidic condition of ABTS oxidation by laccases. As a redox mediator in these acidic conditions help Rhodamine B decolorization. Laccase from Trametes species normally are acid-stable. Laccase from T. hirsuta showed decolorization rate higher at pH 4.0 than at pH 5.0 for indigo carmine, methyl orange and poly R-478 except for bromophenol blue which was nearly total degraded in 2 h at both pH values (Rodríguez et al., 2004). However, apart from optimum stability or oxidation activity issues of enzyme and its substrate, pH condition (or buffer species) might have some effects on oxidation or stability of redox mediators and its radicals.
The initial dye concentration we used in present experiments was four times lower than those used by Lan et al. (2006). In their experiments used 40 μM while we used four times lower (10 μM). However, they reported Rhodamine B decolorization by LiP 0.04 U mL-1 (1.0 μmol U-1). In present study we used dye/enzyme ratio 0.5 μmol U-1 which was two times lower, but no use of H2O2 co-substrate.
Most dye decolorization studies using T. versicolor have been performed at 30°C (Rodríguez et al., 2005) or at room temperature (Rodríguez et al., 2004). In present study, decolorization ability was better at the temperature higher than 30°C. The higher temperature the increase in decolorization percentage until reached its maximum at 35°C. The dye decolorization percentages were decreased at temperature higher than 35°C. This might be because of a thermal denaturation of the enzyme. However, optimum temperature of certain substrates might not relate with dye decolorization.
The decolorization process might be similar to microwave-assisted photocatalytic system by an N-de-ethylation and/or a cleavage of conjugated chromospheres structure as proposed by He et al. (2009). This should be further investigating in order to more understand about the enzymatic mechanism of this laccase.
There are some reports that decolorization might be involved with enzymatic catalysis. For example, the decolorization of six xanthenes dyes (conc. 0.10 mM) by a white rot fungus, Coriolus versicolor has reported in liquid culture by Itoh and Yatome (2004). In their report, no decolorization of Rhodamine class (Rhodamine B, Rhodamine 123 hydrate and Rhodamine 6G) was observed by this study.
One directly reported enzymatic decolorization of Rhodamine B dye was the study of Lan et al. (2006) by LiP coupled with glucose oxidase which reported that three dyes including xylene cyanol, fuchsine and Rhodamine B had a high efficient decolorization. Even though, the decolorization of dyes by LiP does not need the mediator system, the enzyme needs H2O2. This might be a little more complicate to deal with the appropriate conditions (such as optimum in pH and temperature) for both enzymes in catalysis as well as stabilization. To work with a single enzyme in the treatment system might be a more practical approach.
The dye settled down as precipitates instead of decolorization when both naphtha compounds were presence. This interesting result might have a potential use of 1-naphthol and 2-naphthol as co-precipitate substances of other hazardous dyes in enzymatic system. These naphtha compounds which are normally used for dyes, medicinal organics and synthetic perfumes production could be polymerized into new polymeric materials by the oxidative reaction of laccases. During 1-naphthol oxidation a violet colored insoluble product was formed. The product of 2-naphthol oxidation was a white insoluble precipitate. This aspect of applications is still required for further investigation.
In conclusion, we report here an effective ABTS-laccase system for Rhodamine B decolorization. Optimal molar ratios of dye/mediator was 1:10, ratio of dye/enzyme was 0.5 μmol U-1. Temperature and pH optima were at 35-40°C and pH 4.0-5.0. The system with 1-naphthol and 2-naphthol at higher concentrations revealed a settled down of dye instead of a decolorization. More research is needed for potential use of these compounds as co-precipitate substances of other hazardous dyes in enzymatic system.
ACKNOWLEDGMENTS
This study was granted by Mahasarakham University (Fiscal year 2008). The author would like to thank Pimjai Suwannawong for her laboratory assistance. PERCH-CIC and Department of Chemistry, Faculty of Science, Mahasarakham University was also acknowledged.
REFERENCES
- Bourbonnais, R. and M.G. Paice, 1990. Oxidation of non-phenolic substrates. An expanded role for laccase in lignin biodegradation. FEBS Lett., 267: 99-102.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Camarero, S., D. Ibarra, M.J. Martinez and A.T. Martinez, 2005. Lignin-derived compounds as efficient laccase mediators for decolourization of different types of recalcitrant dyes. Applied Environ. Microbiol., 71: 1775-1784.
CrossRefDirect Link - Chhabra, M., S. Mishra and T.R. Sreekrishnan, 2008. Mediator-assisted decolorization and detoxification of textile dyes/dye mixture by Cyathus bulleri Laccase. Applied Biochem. Biotechnol., 151: 587-598.
CrossRef - He, Z., C. Sun, S. Yang, Y. Ding, H. He and Z. Wang, 2009. Photocatalytic degradation of rhodamine B by Bi2WO6with electron accepting agent under microwave irradiation: Mechanism and pathway. J. Hazard Mater., 162: 1477-1486.
CrossRef - Itoh, K. and C. Yatome, 2004. Decolorization and degradation of xanthenes dyes by a white rot fungus, Coriolus versicolor. J. Environ. Sci. Health A. Toxicol. Hazard Subst. Environ. Eng., 39: 2383-2389.
CrossRefDirect Link - Kandelbauer, A., A. Erlacher, A. Cavaco-Paulo and G.M. Guebitz, 2004. Laccase-catalyzed decolorization of the synthetic azo-dye diamond black PV 200 and of some structurally related derivatives. Biocatal. Biotrans., 22: 331-339.
CrossRef - Khammuang, S. and R. Sarnthima, 2007. Laccase from spent mushroom compost of Lentinus polychrous Lev. and its potential for remazol brilliant blue R decolourisation. Biotechnology, 6: 408-413.
CrossRefDirect Link - Lan, J., X. Huang, M. Hu, Y. Li, Y. Qu, P. Gao and D. Wu, 2006. High efficient degradation of dyes with lignin peroxidase coupled with glucose oxidase. J. Biotechnol., 123: 483-490.
CrossRef - Matto, M. and Q. Husain, 2007. Decolorization of direct dyes by salt fractionated turnip proteins enhanced in the presence of hydrogen peroxide and redox mediators. Chemosphere, 69: 338-345.
CrossRefDirect Link - Minussi, R.C., G.M. Pastore and N. Duran, 2007. Laccase induction in fungi and laccase/N-OH mediator systems applied in paper mill effluent. Bioresour. Technol., 98: 158-164.
CrossRef - Moldes, D. and T. Vidal, 2008. Laccase�HBT bleaching of eucalyptus kraft pulp: Influence of the operating conditions. Bioresour. Technol., 99: 8565-8570.
CrossRef - Murugesan, K., I.H. Nam, Y.M. Kim and Y.S. Chang, 2007. Decolorization of reactive dyes by a thermostable laccase produced by Ganoderma lucidum in solid state culture. Enzyme Microb. Tech., 40: 1662-1672.
Direct Link - Nigam, P., G. Armour, I.M. Banat, D. Singh, R Marchant, A.P. McHale and G. McMullan, 2000. Physical removal of textile dyes from effluents and solid-state fermentation of dye-adsorbed agricultural residues. Bioresour. Technol., 72: 219-226.
CrossRefDirect Link - Rodriguez, C.S., E. Rosales, M. Gundin and M.A. Sanroman, 2004. Exploitation of a waste from the brewing industry for laccase production by two Trametes species. J. Food Eng., 64: 423-428.
CrossRefDirect Link - Rodriguez, C.S., M.A. Sanroman and G.M. Gubitz, 2005. Influence of redox mediators and metal ions on synthetic acid dye decolourization by crude laccase from Trametes hirsute. Chemosphere, 58: 417-422.
Direct Link - Svobodova, K., A. Majcherczyk, Č. Novotn� and U. Kues, 2008. Implication of mycelium-associated laccase from Irpex lacteus in the decolorization of synthetic dyes. Bioresour. Technol., 99: 463-471.
CrossRef - Baldrian, P., 2006. Fungal laccases-occurrence and properties. FEMS Microbiol. Rev., 30: 215-242.
CrossRefDirect Link