
M.T. Sakr
Department of Agriculture Botany, Faculty of Agriculture, Mansoura University, 35516 Egypt
A.A. Arafa
Department of Agriculture Botany, Faculty of Agriculture, Mansoura University, 35516 Egypt
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 7 | Page No.: 582-588







ABSTRACT
In this study, two field experiments were carried out during the two growing seasons (2005-2006 and 2006-2007) to investigate the role of some applied antioxidants (spermine 10 mg L-1 and ascorbic acid 200 mg L-1) in counteracting the harmful effect of soil salinity stress (10.1 or 14.6 dS m-1) on canola plants. Growth characters, yield and its components as well as biochemical constituents were studied in the two growing seasons. The results showed that all growth characters including; plant height, leaves number and area/plant, shoot and root dry weight as well as yield and its components including; fruit number/plant, number of fruiting branches, seed number/fruit, seed yield/plant and seed oil content of canola plant were decreased with increasing soil salt level (A2) comparing with (A1). On the other hand, applied antioxidants spermine 10 mg L-1 and ascorbic acid 200 mg L-1) increased growth and yield of canola plant during the two growing seasons. However, the applied antioxidants were more effective under the first soil salt condition (A1) soil salt stress levels (A2) decreased each of photosynthetic pigments, K and P contents, while increased proline, soluble sugar, ascorbic acid, Na and Cl contents compared with (A1). On the other hand, applied antioxidants increased each of photosynthetic pigments, proline, soluble sugar, N, K and P contents, while decreased Na and Cl contents in canola plant under soil salt stress (A1 and A2) during the two growing seasons. It could be concluded that applied antioxidants could counteract the harmful effect of salt soil stress on growth, yield and biochemical constituents of canola plant.
PDF Abstract XML References Citation
How to cite this article
M.T. Sakr and A.A. Arafa, 2009. Effect of Some Antioxidants on Canola Plants Grown under Soil Salt Stress Condition. Pakistan Journal of Biological Sciences, 12: 582-588.
DOI: 10.3923/pjbs.2009.582.588
URL: https://scialert.net/abstract/?doi=pjbs.2009.582.588
DOI: 10.3923/pjbs.2009.582.588
URL: https://scialert.net/abstract/?doi=pjbs.2009.582.588
INTRODUCTION
The winter and spring type of Brassica napus L. and Brassica campestris L. as well as Indian mustard Brassica juncea provide approximately 16% for global vegetable oil requirements (Francois, 1994). Rapeseed oil is the term given to the oil obtained from the seeds of several species of Brassica genus.
Recently, the intensive study of specialists aims to study the responses of plants to salt stress and trying to overcome salt injury by applying antioxidants (Lopez and Satti, 1996). Ashraf and Sharif (1998) on Brassica carinata, Hashem et al. (1998) on Brassica napus and Wright et al. (1995) on canola and Indian mustard. Found that salinity stress reduced plant height, shoot and root dry weight, leaf number, leaf area, pod number/plant, seed number/fruit, 1000 seed weight, seed yield/plant as well as oil and protein content in the seeds.Regarding the effect of antioxidant on canola under salinity stress: Shalata and Neumann (2001) found that ascorbic acid acts directly to neutralize superoxide radicals, singlet oxygen or superoxide and as a secondary anti-oxidant during reductive recycling of the oxidized form of α-tocopherol.
It is now widely accepted that Reactive Oxygen Species (ROS) are responsible for various stress-induced damage to macromolecules and ultimately to cellular structure. Ascorbic acid is an important antioxidant, which reacts not only with H2O2 but also with O2, OH and lipid hydroperoxidases. On the other hand, ascorbic acid has been implicated in several types of biological activities in plants: (1) as an enzyme co-factor, (2) as an antioxidant and (3) as a donor/ acceptor in electron transport at the plasma membrane or in the chloroplasts, all of which are related to oxidative stress resistance (Conklin, 2001). As for the effect of polyamines: Zhao and Qin, 2004 established that polyamines had ameliorating effect on all morphological and physiological characters and prevented degradation of chlorophyll, however polyamines enhanced accumulation of all organic compounds under salinity stress study except phenols.
The objectives of this investigation were to counteract the adverse effect of salinity by exogenously spermine and ascorbic acid application.
MATERIALS AND METHODS
Two field experiments were carried out in the Farm of Field Crops Research Department, National Research Center, Egypt, during the two successive seasons of 2005/2006 and 2006/2007 to investigates the effects of different soil salt on canola growth and yield. In addition, studies on the possibility of using certain antioxidants as spermine and ascorbic for controlling the harmful effects of salinity on different aspects in this investigation were made.
Canola Brassica napus L. cv. Pactol seeds were obtained from the Ministry of Agriculture Oil Crop Research Center Giza, Egypt. Two areas differ in their soil salt were chosen as follows:
• | 1st area (A1) has 10.1 dS m-1 |
| • | 2nd area (A2) has 14.6 dS m-1 |
Antioxidants used: Canola seeds were presoaked for 6 h before sowing in any of applied antioxidants (spermine 10 mg L-1 or ascorbic acid 200 mg L-1) or tap water (control) then canola seeds were sown on 25th November in the two growing seasons (2005/2006 and 2006/2007). After 20 days from sowing, the plants were thinned to leave one plant at a distance 20 cm. The nitrogen fertilizer was added at three intervals after 20, 30 and 60 days after sowing as ammonium nitrate.
Plants were sprayed with the same applied antioxidants or tap water which used in seed presoaking. Automatic atomizers were used for spraying the antioxidants after adding tween 20 as a wetting agent 0.05%. The plants were sprayed at two stages, the first after 40 days from sowing vegetative stage and the second after 70 day from sowing flowering stage. Each treatment replicated 3 times and arranged in a complete randomized block designee.
Sampling dates: In the two growing seasons, five plants representing each treatment were taken randomly after 80 days from sowing. The growth characters (stem and main root length, leaves area and number of leaves per plant and dry weight of shoot and roots) were determined. At harvest (154 days from sowing) number of flowering branches/plant, seeds/siliqua, weight of 100 seed, seed yield/plant g and seed oil yield were recorded.
Biochemical determination: Biochemical constituents of canola plants such as photosynthetic pigments, proline, sugars, ascorbic acid, N, P, K, Na and Cl, contents were determined only in the second season.
Oil content was extracted and determined in the seed according to Folch et al. (1957). The seeds were finely ground in household electric grinder. An aliquot (10 g) from ground seed were extracted by 80 mL chloroform- methanol (2:1 v/v) for 3 h at laboratory temperature with occasional stirring. The mixture was filtered through Whatmann No. 1 filter paper, the seeds were retracted three times, then filtrated and the filtrates were combined together.
Photosynthetic pigments: Photosynthetic pigments were determined in the 4th upper leaf after 80 days from sowing according to the methods of MacKinney (1941). Weight of 0.05 g fresh material was extracted by 10 mL methanol 90% for 24 h. Then both chlorophyll and carotenoid were determined by spectrophotometer at the wave length 452.5, 650 and 665 nm.
Proline content was determined in canola shoot after (60 days from sowing) by the modified ninhydrin method of Troll and Lindsley (1955). Weight of 2 g from shoot fresh weight were placed into tube containing 10 mL of distilled water. The tubes were kept in boiling water bath for 30 min, then cooled at room temperature. An aliquot from corresponding water extract, added 2 mL of ninhydrin reagent and the mixture were maintained in boiling water bath for 20 min, the mixture was cooled in an ice-water bath. The product formed was extracted by toluene by vigorous shaking. Absorbance was measured at 520 nm by spectrophotometer.
Total soluble carbohydrates were extracted from 0.1 g shoot dry matter by ethanol 80% for overnight at laboratory temperature, then filtered through Whatman No. 1 filter paper. Total soluble sugars were determined by the anthron methods (Sadasivam and Manickam, 1996) through adding 3 mL anthron reagent to 0.1 mL filtrate, then heated for 10 min in a boiling water bath, cool rapidly and the developed green color was read at 630 nm by spectrophotometer.
Ascorbic acid was extracted from 2 g shoot fresh material by 4% oxalic acid, then made up to a known volume (100 mL) and centrifuged at 2000 rpm for 5 min. then added 10 mL of 4% oxalic acid then titrate using 2,6-dichlorphenol-indophenol as described by Sadasivam and Manickam (1996).
Total nitrogen content: Sample of 0.2 g dry material were digested by sulphuric and perchloric acids using Micro-Kjeldahl method (Jackson, 1967). Distillation was carried out with 40% NaOH and ammonia was received in 4% boric acid solution.
Sodium, potassium and chloride content: weight of 0.2 g dry matter from canola shoot was extracted (Chaudhary et al., 1996) for 1 h in a boiling-tube of distilled water in a boiling water bath, the extract was filtered. Sodium and potassium content in the aqueous extracts were measured with Flame Photometer. Meanwhile, chloride was determined by titration with 0.001 N AgNO2 using potassium dichromate as indicator.
Phosphorous content: Phosphorous was determined calorimetrically at wavelength 725 nm using chlorostannous-reduced molybdo phosphoric blue color method, as described by Jackson (1967).
Statistical analysis: Data were subjected to statistical analysis of variance according to Gomez and Gomez (1984) and LSD value for comparison.
RESULTS
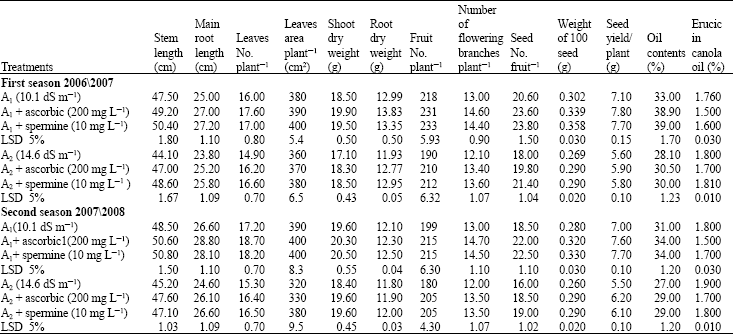
Growth and yield experiment: Data presented in Table 1 show that all growth characters including stem and root length, leaves number and leaves area per plant as well as shoot and root dry weight were significantly decreased with high salinity level (soil salt, A2) throughout the two growing seasons. The great reduction in these parameters was observed under high soil salt (A2; 14.6 dS m-1) compared with (A1; 10.1 dS m-1).
Data in the same table reveal that all growth characters increased with antioxidants (spermine 10 mg L-1 or ascorbic acid 200 mg L-1 ) used above untreated plants. Spermine was the most effective in this respect.
Both antioxidants (spermine 10 mg L-1 or ascorbic acid 200 mg L-1) counteracted the harmful effects of salinity stress (soil salt) on growth characters of canola plant.
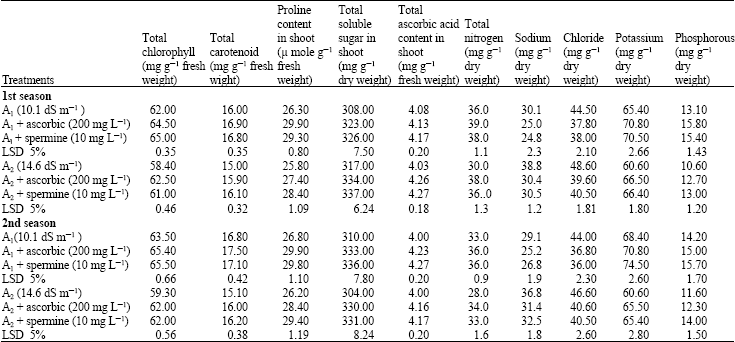
Biochemical constituents: Data in Table 2 show that high soil salt, A2 (14.6 dS m-1) decreased each of chlorophyll, carotenoid, nitrogen, potassium and phosphorus contents, while increased proline, soluble sugar, ascorbic acid, sodium and chloride contents compared with soil salt area, A1 (10.1 dS m-1). Applied antioxidants (spermine 10 mg L-1 and ascorbic acid 200 mg L-1) increased each of chlorophyll, carotenoid, proline, soluble sugar, ascorbic acid, nitrogen, potassium and phosphorus contents. While decreased sodium and chloride contents under the salt soil areas (A1 and A2) throughout the growing seasons.
It could be noticed that applied antioxidants could partially counteract the harmful effect of soil salinity stress on the biochemical constituents of canola plants under the experimental conditions.
Yield and its components: Data presented in Table 1 show that, yield and its components fruit number per plant; number of flowering branches per plant; seed number per fruit; seed index and seed yield per plant of canola plant were significantly decreased with increasing soil salt levels especially under the higher soil salinity level, A2 (14.6 dS m-1). On the other hand, exogenous application of both antioxidants (spermine 10 mg L-1 and ascorbic acid 200 mg L-1) significantly increased yield and its component above untreated canola plants under the two soil salt areas (A1 and A2) throughout the two growing seasons.
Applied spermine was the most effective than ascorbic in counteracting the harmful effects of salinity on yield and its components.
| Table 1: | Effect of spermine 10 mg L-1 or ascorbic acid 200 mg L-1 as well as their combinations with soil salt levels on growth and yield of canola plants during the two growing seasons, 2005/2006 and 2006/2007 |
 | |
| Table 2: | Effect of spermine 10 mg L-1 or ascorbic acid 200 mg L-1 as well as their combinations with soil salt levels on some biochemical constituents of canola plants during the two growing seasons, 2005/2006 and 2006/2007 |
 | |
Seed oil content: Data in Table 1 indicate that canola seed oil yield was significantly decreased with increasing soil salt stress (A2) compared with soil salt area (A1). On the other hand, application of both antioxidants increased seed oil content and enhanced the plants to tolerate the harmful effect of high salinity level in this respect.
Erucic content in canola oil seeds: The results indicated clearly that the high soil salt (A2) increased the erucic acid percentage up to the harmful effect to human comparing with soil salt area (A1). Whereas applied antioxidants led to a decrease in this respect.
DISCUSSION
Growth and yield: The inhibitory effect of soil salt on canola growth in the present investigation may be due to:
| • | Decreasing meristematic activity and/or cell enlargement (Khadr et al., 1994; Sakr et al., 2007) |
| • | Damaging growth cells so that they can not perform their functions |
| • | Limiting their supply of essential metabolites |
| • | Perturbing the functioning of vital components of photosynthesis (Yang and Britton, 1990) |
The reduction in seed yield is largely due to:
| • | Reduction in pollen viability has been related to decreased calcium mobilization from plant leaves treated with sodium chloride, which is important in pollen germination and pollen tube growth |
| • | Abscission of flowers or young fruit due to ethylene induction by salinity |
| • | Decreasing the leaf area and number per plant, resulting reduction in the supply of carbon assimilate due to decreasing the net photosynthetic rate and biomass accumulation (Sakr et al., 2007) |
Photosynthetic pigments contents: Photosynthetic pigments in canola leaves were significantly decreased with increasing soil salt levels. This reduction may be related to enhancing the activity of chlorophyll degrading enzyme chlorophyllase (Mishra and Sharma, 1994).
It could be concluded that, ion accumulation in leaves specially chloride and sodium, affect chlorophyll biosynthesis due to its effect on the activity of Fe containing enzymes, cytochrome oxidase (Munne-Bosch et al., 1999).
Proline accumulation: Proline accumulation is one of the most frequently reported modifications induced by water deficit and salt stress in plants and it is involved in stress resistance mechanisms (Sakr et al., 2007). Several functions are proposed for the accumulation of proline in tissues submitted to salt stress:
| • | Osmotic adjustment |
| • | C and N reserve for growth after stress relief |
| • | Stabilization of proteins and/or membranes |
| • | Being a scavenger of free radicals |
| • | Proline plays an important role in the protection of enzymes or membranes against salinity (Ozdemir et al., 2004) |
Na+, Cl– and K + contents: In a saline environment, plants take up excessive amount of Na+ and Cl– as in halophytes resulted in high Na+/Ca2+ and Na+/k+ ratios which may impair the selectivity of the root membrane. The accumulation of Na+ in plant root may be due to the high mobility of Na+ in the phloem (Ozdemir et al., 2004).
Chloride is a more sensitive indicator of salt damage than sodium, since the plant stores it. Ozdemir et al. (2004) proposed that the relatively greater uptake of Cl– than Na+ in salt-stressed plants could be responsible for growth reduction by depressing the uptake of other anions.
The reduction of internal potassium concentration could be related to:
| • | Increased potassium efflux into the growth medium (Cramer et al., 1989), due to a disrupt in membrane integrity caused by Na+ and inhibit transport of these ion into root and up to the shoot |
| • | The antagonism between K+ and Na+ cations, which increased considerable as salinity increased (Sariam and Srivastava, 2002) |
Phosphorus, nitrogen and calcium contents: The decrease in P concentration associated with salinity conditions may be ascribed to a decrease in the translocation of P upward through the stem because of the increase in the osmotic pressure of the root medium (Sakr et al., 2007).
As for nitrogen, an increase in chloride uptake and accumulation is accompanied by a decrease in shoot nitrate concentrations of plants due to the compilation between chloride and nitrate, which decreases nitrate content (Sakr et al., 2007).
Regarding calcium, calcium concentration declined with increasing salinity due to the high sodium levels in the external media which reduced the activity of calcium in the solution and/or decrease the amount of calcium available for uptake by the plant and/or the antagonism of sodium and calcium at the site of uptake in roots and/or to the inhibition of uptake processes (Sakr et al., 2007).
Sugar contents: The accumulation of non-reducing sugar was the result of an enhanced efficiency in the use of carbon coupled to a reduction in cellular metabolism that could favor the accumulation of respiratory substrate to support the osmotic adjustment required to survive in saline media (Munns and Termaat, 1986).
The marked increase in soluble sugar in antioxidants treated plants in most cases over these of the control explained by ascorbic acid and spermine might alleviate the imposed salinity stress, either via osmotic adjustment (Stewart and Lee, 1974) or by conferring desiccation resistance to plant cell (Thakur and Rai, 1985).
Role of antioxidants on growth and yield under salinity stress: The promotive effect of antioxidants on stem and root length may be the result of increasing cell division in the apical meristem or increased cell division and cell enlargement (Arteca, 1996) due to water uptake caused by a decrease in the osmotic potential by increasing soluble sugars (Table 1, 2). In addition, both osmoregulators increased phosphorous content (Table 2) which is essential for cell division and development of meristem tissues (Foyer and Spancer, 1986).
The promotive effect of spermine and ascorbic acid on leaf number may be attributed to the high hygroscopic nature of antioxidants or decreasing sodium and chloride concentration. Results show that the antioxidants increased total soluble sugar (Table 2) which serve as a substrate for increasing initiation of leaf primordial and decreased the plastochron duration. The application of osmoregulators led to accumulation of non-toxic solutes i.e., proline, soluble sugar, phosphorous and potassium (Table 2) in whole plants. Such accumulations provide the turgor necessary for cell expansion resulting in increasing leaf area (Munns and Termaat, 1986).
The effect of antioxidant son on dry matter may be due to:
| • | An increase in leaf area (Table 1) |
| • | Stabilizing the enzymes involved in amino acid metabolism (Laurce and Stewart, 1990) |
| • | Increased potassium content in plant material (Table 2), which leads to an increase in leaf area, enhanced the production of photosynthates and their subsequent translocation resulting in dry matter accumulation |
| • | Enhanced phosphorous available for metabolism which increased photosynthetic rates (Table 2), resulting in increasing dry matter accumulation (Foyer and Spancer, 1986) |
Regarding canola seed yield; the increase in seed yield due to applied antioxidants may be the result of increasing potassium content as shown in Table 2 which enhancied photosynthetic pigments and capacity, leading to enhanced dry matter accumulation (Table 1), which in turn, resulted in an increase in seed yield due to an increase of assimilated food needed for plant production.
Regarding photosynthetic pigments: The stimulating effect of antioxidants on photosynthetic pigments may be due to stabilizing active site of enzymes (Munne-Bosch et al., 1999). On the other hand, antioxidants increased some ions as Mg2+ that needed for chlorophyll synthesis (Shaddad, 1990). In addition, increased potassium content (Table 2), which increase leaf photosynthetic efficiency possibly by increasing the number of chloroplast per cell, number of cell per leaf and consequently leaf area (Possingham, 1980).
These results mean that, antioxidants application could be used to minimize the harmful effect of salinity on leaf pigments, which consequently may enable the plant to withstand salinity conditions.
REFERENCES
- Hashem, A., M.N.A. Majumdar, A. Hamid and M.M. Hossain, 1998. Drought stress effects on seed yield, yield attributes, growth, cell membrane stability and gas exchange of synthesized Brassica napus. J. Agron. Crop Sci., 180: 129-136.
CrossRef - Chaudhary, M.T., S.J. Wainwright and M.J. Merrett, 1996. Comparative NaCL tolerance of Lucerne plants regenerated from salt-selected suspension cultures. Plant Sci., 114: 221-232.
Direct Link - Conklin, P., 2001. Recent advances in the role and biosynthesis of ascorbic acid in plants. Plant Cell Environ., 24: 383-394.
PubMed - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Foyer, C. and C. Spencer, 1986. The relationship between phosphate status and photosynthesis in leaves. Effects on intracellular orthophosphate distribution, photosynthesis and assimilate partitioning. Planta, 167: 369-375.
CrossRefDirect Link - Francois, L.E., 1994. Growth, seed yield and oil content of canola grown under saline conditions. Agron. J., 86: 233-237.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Khadr, I., F. Nyireda, F. Shanahan, C. Nielsen and R. Andria, 1994. Ethephon alters corn growth under drought stress. Agron. J., 86: 283-288.
Direct Link - Lopez, M.V. and S.M.E. Satti, 1996. Calcium and potassium-enhanced growth and yield of tomato under sodium chloride stress. Plant Sci., 114: 19-27.
CrossRefDirect Link - Mackinney, G., 1941. Absorption of light by chlorophyll solutions. J. Biol. Chem., 104: 315-322.
Direct Link - Munns, R. and A. Termaat, 1986. Whole-plant responses to salinity Aust. J. Plant Physiol., 13: 143-160.
CrossRefDirect Link - Ozdemir, F., B. Melike, T. Demiral and I. Turkan, 2004. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul., 42: 203-211.
CrossRefDirect Link - Possingham, J.V., 1980. Plastid replication and development in the Life cycle of higher plants. Ann. Rev. Plant Physiol., 31: 113-129.
CrossRefDirect Link - Sairam, R.K. and G.C. Srivastava, 2002. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci., 162: 897-904.
CrossRefDirect Link - Shaddad, M.A., 1990. The effect of proline application on the physiology of Raphanus sativus plants grown under salinity stress. Biol. Plant, 32: 104-112.
CrossRefDirect Link - Shalata, A. and P.M. Neumann, 2001. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. J. Exp. Bot., 52: 2207-2211.
CrossRefDirect Link - Stewart, C.K. and J.A. Lee, 1974. The role of proline accumulation in halophytes. Planta, 120: 279-289.
CrossRefDirect Link - Thakur, P.S. and V.K. Rai, 1985. Exogenously supplied amino acids and water deficits in Zea mays cultivars. Biol. Plant, 27: 458-461.
CrossRefDirect Link - Troll, W. and J. Lindsley, 1955. A photometric method for the determination of proline. J. Biol. Chem., 215: 655-660.
Direct Link - Zhao, F.G. and P. Qin, 2004. Protective effect of exogenous polyamines on root tonoplast function against salt stress in barley seedlings. Plant Growth Regul., 1: 97-103.
Direct Link