
A.S. Sarimin
School of Environmental and Natural Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
M.A. Ghaffar
School of Environmental and Natural Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
C.A.R. Mohamed
Marine Ecosystem Research Centre, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 3 | Page No.: 231-238







ABSTRACT
A study on elemental composition in the otolith of giant mudskipper, Periophthalmodon schlosseri, was done from June to October 2003. Specimens were obtained from the mangrove areas of Kuala Selangor, Sepang and Melaka in the west coast of Peninsular Malaysia. A total of 70 sagitta otoliths were analyzed to detect variation of Sr, Ba and Mg, replacing the natural chemical composition of the otolith, which is the calcium carbonate (CaCO3). The average ratio of Sr:Ca was 0.11x10-4, Ba:Ca was 5.7x10-3 and Mg:Ca was 0.2x10-3. Strong correlation (R>0.8) between fish body size and otolith weight of mudskipper (p<0.01) also found during this study.
PDF Abstract XML References Citation
How to cite this article
A.S. Sarimin, M.A. Ghaffar and C.A.R. Mohamed, 2009. Variation of Ca, Sr, Ba and Mg in the Otolith of Mudskipper in West Coast of Peninsular Malaysia. Pakistan Journal of Biological Sciences, 12: 231-238.
DOI: 10.3923/pjbs.2009.231.238
URL: https://scialert.net/abstract/?doi=pjbs.2009.231.238
DOI: 10.3923/pjbs.2009.231.238
URL: https://scialert.net/abstract/?doi=pjbs.2009.231.238
INTRODUCTION
Otolith has been widely used as a tool in determining the age, fish growth, reproduction and fish migratory pattern (Secor and Rooker, 2000; Dwyer et al., 2003; Grandcourt et al., 2006; Walther and Thorrold, 2008; Leakey et al., 2009) in all types of fish, such as demersal and pelagic. Recent researches on otolith-water chemistry have been focused on interaction between ambient water and the otolith (Milton and Chenery, 2001). Giant mudskipper, Periophthalmodon schlosseri is different from the common fish because it spends more time on land than in water, searching, hunting, stalking and consuming prey in the mudflat (Mazlan et al., 2006). Furthermore, P. schlosseri can tolerate different types of ambient water (e.g., brackish, seawater and freshwater) and use the land adapted gill to breathe on land (Mc Inerny and Gerard, 1996). This fish holds air in its large buccopharyngeal-opercular cavity, where respiratory gas exchange takes place via gill and highly vascularized epithelium lining cavity (Aguilar et al., 2000). The entire life span of giant mudskipper is spent mainly on mudflat, where they dig burrows to seek refuge, lay eggs and escape from marine predators (Brillet, 1969; Sasekumar et al., 1994). Mudskippers generally deposit their eggs on the ceiling of the burrow and continue to develop until they are independent enough to leave the burrows (Kobayashi et al., 1971; Clayton 1993). The embryo aerates using the oxygen stored in the burrow by the parent (Ishimatsu et al., 1998).
Otolith functions as elemental recorder, which grows throughout the lifespan of fish and the materials incorporated onto the otolith matrix become permanent, even though the fish is distressed or starved (Campana and Neilson, 1985; Miller et al., 2006). Elsdon and Gillanders (2004) have shown that a significant relationship exist between ambient water chemistry and otolith chemistry. Most of the studies involving divalent elements, such as Mg, Ba, Mn and Sr, show strong relationship, while other elements, such as Li, Cu and Pb, do not show significant relationship (Milton and Chenery, 2001), suggesting that divalent element might substitute Ca in the calcified aragonitic structure of otolith. Ba and Sr have become important tools for determining fish migratory based on the assumption of the different content of Sr and Ba in seawater, brackish water and freshwater (Elsdon and Gillanders, 2003a, b). Compared to scales and spines, which are made from calcium phosphate, the discrete layer of otolith is made from calcium carbonate and is not directly in contact with ambient water (Gillanders, 2001). Studies on diadromous fish (e.g., sea bass) are more documented compared with those of mudskippers, which are amphidromous.
The aim of this study is to measure the concentration of Ca, Sr, Ba and Mg in the giant mudskipper sagitta otolith and to identify the influence of the menacing environment of the unique giant mudskipper to the otolith chemistry.
MATERIALS AND METHODS
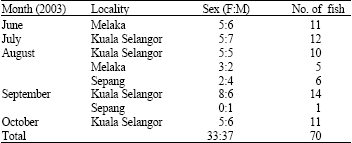
Three sampling locations of mudskipper habitat were selected namely Kuala Selangor, Sepang and Melaka from the west coast of Peninsular Malaysia (Fig. 1). This species is usually found and abundant in the mangroves and mudflat areas. In Melaka coast, the giant mudskippers were found on the mudflats but Kuala Selangor and Sepang areas were found in mangroves area. The salinity value at the sampling area ranged from 6-32 psu (Phuah, 2004; Tee, 2003). About 70 specimens of fish sample were collected from June to September 2003 in four occasions for Kuala Selangor, two occasions each for Sepang and Melaka (Table 1). Then the fish samples were dissected to extract their otolith (Secor et al., 1991).
All glassware and plastic materials used in the laboratory and during sampling were soaked in 5% of nitric acid (HNO3) overnight. Fish samples were randomly collected using fishing rods and nets. Specimens were weighed with electronic balance up to 0.01 g and the length was measured using a measuring scale board to the nearest 0.1 cm scale. The otolith length was measured with a digital electronic calliper with accuracy ± 0.01 mm while the otolith samples were weighted using the electronic balance with an accuracy of ± 0.01 mg.
| Table 1: | Samplings locations of giant mudskipper on year 2003 |
 | |
| Table 2: | The analyzed value of 3 replicates of CRM N0.22 and the recovery (%) based on certificate values |
 | |
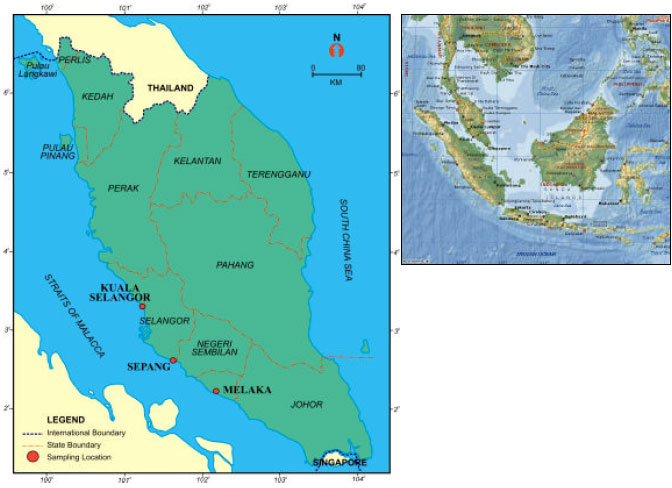 | |
| Fig. 1: | Samplings locations in Kuala Selangor, Sepang (Selangor) and Melaka in the West coast Peninsular of Malaysia |
About 10 mL of concentrated HNO3 was added into a glass beaker to digest the sagitta otolith. The samples were dried and redissolved with 20 mL of 0.5 M HCl. Concentrations of Ca, Sr, Ba and Mg were measured using the Perkin Elmer Optima 4300DV Inductive Coupled Plasma Omitted Emission Spectrophotometer (ICP-OES). Replicates of Certified Reference Material No. 22 (Otolith) were also prepared using the same procedure as otolith samples where the measurement results were in the range of certified values (Table 2).
RESULTS
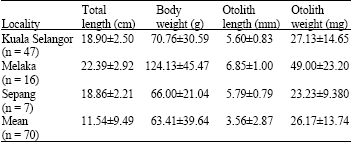
Relationship between giant mudskipper and otolith: The shape of the extracted sagitta otolith is round (Fig. 2). The heaviest and longest fish in Melaka acquired the heaviest and longest sagitta otolith, while the smallest and shortest fish in Sepang produced the lightest otolith (Table 3, 4). The difference in weight suggest that it is as a better indicator for relative growth of giant mudskipper in coherence with higher correlation value between fish length/weight with otolith weight (R≥0.89, p>0.01, Table 4). The correlation coefficient shows strong relationship between giant mudskipper and otolith (R>0.8, p>0.01) supporting the assumption that fish size (much more likely to represent the fish growth) influences the size of otolith (Table 4). This indicates that otolith development of giant mudskipper is dependent on cell growth. The relationship between fish length-weight revealed the highest correlation (R = 0.979, p>0.01), supporting that fish length various with length (Table 4). The strong relationship of the otolith and fish suggests that otolith might be a useful indicator of the size of a giant mudskipper (Table 4).
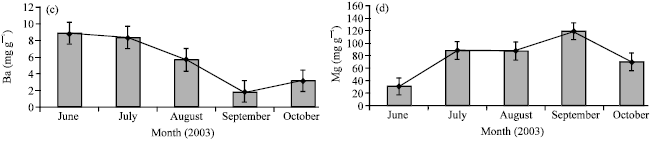
Elemental analysis of otolith: The concentration pattern of the elements varied from 64.30 to 1998.69 mg g-1 for Ca, 0.29 to 10.86 mg g-1 for Sr, 0 to 31.49 μg g-1 for Ba, 11.19 to 575.14 μg g-1 for Mg. There was no clear trend for average concentration of Ca and Sr obtained during this study (Fig. 3a-d). In comparison between the analyzed elements, Ba recorded concentration. In Fig. 3c and d, average concentrations of Ba and Mg showed inverse relationship, where the lowest and highest Ba concentration was recorded in June and September and in reverse with, respectively.
| Table 3: | Total length and body weight of giant mudskipper obtained from the research locations |
 | |
 | |
| Fig. 2: | A pair of giant mudskipper’s sagitta |
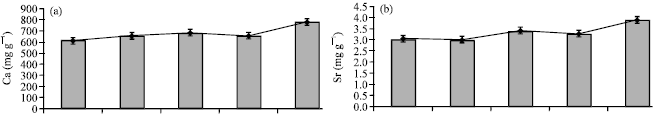 | |
| Fig. 3: | Average concentrations of (a) Ca, (b) Sr, (c) Ba and (d) Mg in the giant mudskipper otolith according to months |
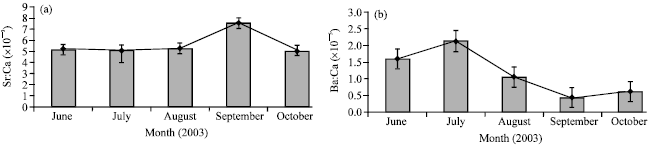 | |
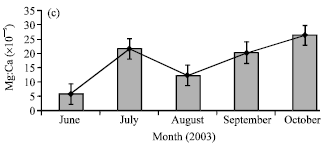
| Fig. 4: | Graph of (a) Sr:Ca, (b) Ba:Ca and (c) Mg:Ca ratios (Mean ± SE) in the giant mudskipper otolith according to months |
| Table 4: | Relationship between giant mudskipper and otolith, Pearson correlation |
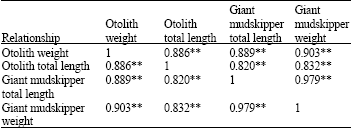 | |
| n = 70. **Correlation is significant at the 0.01 level (2-tailed), p<0.01 | |
From this results, the monthly distribution pattern of Sr:Ca ratio and Sr average concentration differed, in which the highest Sr concentration was observed in September, while Sr:Ca ratio was maximum in October (Fig. 3b, 4a). Ratios of Ba:Ca and Mg:Ca revealed a similar trends with Ba and Mg average concentration (Fig. 3c, d, 4b, c).
DISCUSSION
Characteristic of Ca, Sr, Ba and Mg in the mudskipper otolith: Otolith was developed from the daily crystallization process of fluid within the endolymphatic canal of the fish inner ear in the form of concentric layers series around the nucleus (Farrell and Campana, 1996; Thorrold et al., 1997). This growth layer will stop when the growth of the fish stops which is directly related to otolith size/weight to fish size/weight (Romanek and Gauldie, 1996).
Mazlan and Rohaya (2008) also found high correlation (R>0.9) between daily rings of otolith with the size of giant mudskipper, similar to the results obtained from this research. The variability of the otolith shape decreases the accuracy of the length measured, compared to otolith weight (Strelcheck et al., 2003). In addition, otolith weight is a preferred indicator of relative growth difference in juvenile gag (Mycteroperca microlepis) compared to otolith length (Strelcheck et al., 2003). Mazlan and Rohaya (2008) estimated that the growth parameter for giant mudskipper were L8 = 29 cm and K = 1.4 year-1, where the body growth is influenced by the growth of otolith.
Generally, otolith is composed of 99% CaCO3 (Payan et al., 2004) and Ca is the highest element in otolith (Fig. 3a). The presence of Sr, Mg and Ba by ranking in the otolith suggest that other divalent transition element can substitute Ca because the atomic radii of Mg, Ba and Sr are 0.065, 0.113 and 0.135 Å, respectively, similar to the Ca atomic radius, 0.099 Å (Gauldie et al., 1995). The other elements such as Sr, Mg and Ba have similar atomic radii with Ca in the ambient water are taken in through the gill by active transport of Ca. The Ca transport system, located in chloride cells of gills (Campana, 1999; Chowdhury and Blust, 2002), takes up Sr greater than Ba because concentration of Sr in brackish water exceeds that of Ba (Hamer et al., 2006) (Fig. 3b). In contrast, the concentration of Mg in brackish water is higher than Sr but the uptake of Sr is greater than Mg (Hamer et al., 2006). It also means that the concentration levels of trace elements in otolith are not directly related to the level of trace elements in water column at study site.
Despite other calcified structures, such as corals and bones, the incorporation of elements onto otolith consists of three main interfaces: brachial uptake, cellular transport and crystallization. Elements, which are mostly divalent elements, must pass these barriers before they are crystallized onto otoliths matrix (Campana, 1999). In fact, the final precipitation of elements onto otolith is due to physical chemistry of biomineralisation (Nielson and Christoffersen, 1982), which is affected by physiological process (Ca+) and environmental variables (Hoff and Fuiman, 1995). The elements incorporate onto otolith which represents the influence of the environment to the entire life span of the giant mudskipper, which inhabits the mangrove throughout its lifetime (Ishimatsu et al., 1998).
Salinity recorded in study sites is between 6-32 psu (Phuah, 2004; Tee, 2003), which was lower than that of seawater. Water flow in the study areas was affected by seawater intrusion from the tidal wave in the mangrove systems and dilution was influenced by the freshwater input from the nearest river (Pritchard, 1955). Hence, the concentration level of elements is increased due to the mixing of freshwater and seawater happens. In the present study, Sr concentration is high compared to freshwater because of the high salinity (Fig. 3). Even though the monsoon season brought heavy rainfall from September to December, no significant difference is traced from Sr, Mg and Ba, trends from September to October (Fig. 3b, c). There were also no obvious trends indicating the differences from the pre-monsoon season suggesting that monsoon season is not affecting the water flow system in the study areas (Fig. 3). The monthly Mean Sea Level (MSL) recorded in Melaka and Selangor was insignificantly different (JUPEM, 2003).
Brackish water fishes are usually high uptake of Sr than Ba because concentrate level of Sr in brackish water column (de Vries et al., 2005). While, Mg and Ba contents show the contradictory pattern suggesting that Mg and Ba are replacing each other by getting the influence of the freshwater input in the study area (Fig. 3c, d).
Giant mudskippers inhabit estuary in the tidal mudflat in the mangroves forest. Generally, the hydrology system in mangroves of Malaysia is influenced by rainfall, evaporation and tidal flushing (Chan, 1987). It had been proven that Ba content is higher in ambient water with fresh water intrusion, which is rain water and river (Elsdon and Gillander, 2003a, b, 2004). This freshwater intrusion factors most likely is influence by monsoon season and tides. However, this study revealed that these factors were not significantly affect the Ba content in otolith (Fig. 3c). During monsoon season, heavy rain fall will happen from September to December and less rain during pre-monsoon period (Cheang, 1987). This will decrease the salinity value of ambient water and enhancement the Ba levels. In this study, suggesting, the water contents of estuary will affect the concentration levels of Ba in the otolith (Fig. 3c). This also indicates, the sampling stations located in mangrove area is real estuarine (Pritchard, 1955), where a low salinity value at the top layer and high value at the bottom layer of water body.
| Table 5: | Sr:Ca, Ba:Ca and Mg:Ca ratios in sea water, brackish water, fresh water and giant mudskipper otolith |
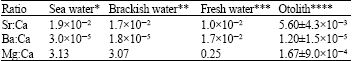 | |
| *Summerhayes and Thorpe (1996), **Hamer et al. (2006), ***SRM NIST 1643e, ****Giant mudskipper otolith ratios | |
In addition, larvae of giant mudskippers develop in deep burrows dug in soft sediment in mudflats (Brillet, 1969). Although burrow condition is hypoxic, the larvae can still survive because the burrows accumulate with air by the adult not only for the larvae to develop, but also to provide oxygen to adult during thermoregulate (Tytler and Vaughan, 1983) and avoiding from predators (Sasekumar et al., 1994). The giant mudskipper acquires low salinity water from the surface of the water body and did not swim deeper or stay in the high salinity of water body. This study suggests that high Ba and Sr in the otolith suggested that the water body inhabited by the giant mudskipper was brackish water and low Mg support the low sea water intrusion to the study area.
Ratios of elements to calcium: Calcium (Ca) is locally incorporated in otolith but others divalent elements such as Sr, Ba and Mg, have the ability to substitute with calcium ion in natural chemical composition depending on environmental factors (Gauldie et al., 1995). The ratio of elements to Ca is determined to detect the variation of substitute elements (i.e., Sr, Ba and Mg) in the otolith. The ratios of elements to Ca are lower than the values of seawater, brackish water and freshwater (Table 5). The incorporation of Ca on otolith matrix is a natural process and not related to the content of Ca in the ambient water (Farrell and Campana, 1996; Thorrold et al., 1997).
In this study, the ratios of Sr:Ca are used as indicators to investigate the changing of salinity in various marine environments (Campana and Tzeng, 2000) because high Sr:Ca ratio value is found in the saline waters (Table 5). On the other hand, Mg:Ca ratios were used as elemental fingerprints (Campana and Tzeng, 2000) but the Ba:Ca ratios will revealed the migration patterns of fish from brackish water to low salinity gradient value during breeding season (Milton and Chenery, 2001). But during investigation, the giant mudskippers were staying near to their burrows with a low freshwater input. The foreign divalent element (e.g., Sr, Ba and Mg) ratio to Ca indicates the acceptance of the otolith towards these elements based on the similarity of the atomic radii to Ca atomic radius. The Sr:Ca ratios were the highest ratio obtained during this study because of its availability in ambient water. While the abundant of Mg value in otolith is not only as signal the seawater input but also indicate the uptake process occur during fish growing in various marine environment.
This research also revealed that the dominances life time on giant mudskippers were on mudflat by comparing with the Sr:Ca, Ba:Ca and Mg:Ca ratios in seawater, brackish water and freshwater system. Then during investigation the giant mudskippers are moving in the ranged of 4200 square feet from their burrow for food searching (Murdy, 1986) as shows by the low ratio value of Ba:Ca from June to October 2003 (Fig. 4b). The contradictory pattern of Mg:Ca ratio and Ba:Ca ratio found in the otolith is similar ratios in seawater, brackish water and freshwater (Table 3) but that does not mean that Ca in seawater affected otolith. Ca incorporated on in otolith by the physiological endolymph uptake of fish (Simkiss, 1974; Farrell and Campana, 1996).
CONCLUSION
The concentration levels of Ca, Sr, Ba and Mg in otolith are related to the ambient water inhabited by giant mudskipper will revealed as Sr>Mg>Ba. The suitability and preferences of elements not only depends on the atomic radius closest to Ca atomic radius, but also the abundances of divalent elements (Mg and Ba) in the ambient water causing high uptake of elements onto otolith. The ratio of Sr:Ca is higher than Ba:Ca ratio, indicating that the lifespan of giant mudskipper is spent near the burrow area, they do not migrate to less saline (<6 psu) water body, upstream nor downstream.
ACKNOWLEDGMENTS
The authors would like to thanks Associate Dr. Takaomi Arai (Ocean Research Institute, University of Tokyo) and all staffs of Universiti Kebangsaan Malaysia and Land Atmosphere Ocean Interaction (LAOI) members for their help during sampling in the field and experimental analysis.
REFERENCES
- Aguilar, N.M., I. Atsushi, O. Kogi and K.H. Khoo, 2000. Aerial ventilory responses of the mudskipper Periophthalmodon schlosseri, to altered aerial and aquatic respiratory gas concentrations. Comput. Biochem. Physiol. A Mol. Integr. Physiol., 127: 285-292.
PubMed - Campana, S.E., 1999. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Series, 188: 263-297.
CrossRef - Campana, S.E. and J.D. Neilson, 1985. Microstructure of fish otoliths. Can. J. Fish. Aquat. Sci., 42: 1014-1032.
CrossRefDirect Link - Chowdhury, M.J. and R. Blust, 2002. Bioavaliabilty of waterborne strontium to the common carp, Cyprinus carpio, in complexing environments. Aquat. Toxicol., 58: 215-227.
Direct Link - Dwyer K.S., J.W. Stephen and S.E. Campana, 2003. Age determination, validation and growth of grand bank yellowtail flounder (Limanda ferruginea). ICES J. Mar. Sci., 60: 1123-1138.
CrossRefDirect Link - Elsdon, T.S. and B.M. Gillanders, 2003. Reconstructing migratory patterns of fish based on environmental influences on otolith chemistry. Rev. Fish Biol. Fish., 13: 217-235.
Direct Link - Elsdon, T.S. and B.M. Gillanders, 2003. Relationship between water and otolith elemental concentrations in juvenile black bream, Acanthopagrus butcheri. Mar. Ecol. Prog. Ser., 260: 263-272.
Direct Link - Elsdon, T.S. and B.M. Gillanders, 2004. Fish otolith chemistry influenced by exposure to multiple environmental variables. J. Exp. Biol. Ecol., 313: 269-284.
Direct Link - Farrell, J. and S.E. Campana, 1996. Regulation of calcium and strontium deposition on the otoliths of juvenile tilapia, Oreochromis niloticus. Comput. Biochem. Phys., 115: 103-109.
CrossRefDirect Link - Gillanders, B.M., 2001. Trace metal in four structures of fish and their use for estimates of stock structure. Fish. Bull., 99: 410-419.
Direct Link - Grandcourt, E.M., T.Z. Al Abdessalaam and F. Francis, 2006. Age, growth, mortality and reproduction of the blackspot snapper, Lutjanus fulviflamma (Forssk�l, 1775), in the Southern Arab Gulf. Fish. Res., 78: 203-210.
CrossRefDirect Link - Hamer, P.A., G.P. Jenkins and P. Coutin, 2006. Barium variation in Pagrus auratus (Sparidae) otoliths: A potential indicator of migration between an embayment and ocean waters in South-Eastern Australia. Estuarine Coastal Shelf Sci., 68: 686-702.
Direct Link - Hoff, G.R. and L.A. Fuiman, 1995. Environmentally induced variation in elemental composition of red drum (Sciaenops ocellatus) otoliths. Bull. Mar. Sci., 56: 576-591.
Direct Link - Ishimatsu, A., Y. Hishida, T. Takita, T. Kanda, S. Oikawa, T. Takeda and K.H. Khoo, 1998. Mudskipper store air in their burrows. Nature, 391: 237-238.
Direct Link - Mazlan, A.G., Y. Faridah, M.N. Shukor and A. Aziz, 2006. Foraging behavior and food selestion of giant Mudskipper (Periophthalmdon schlosseri) at Kuala Gula, Matang Mangrove Reserve, Perak, Malaysia. Coastal Mar. Sci., 30: 263-267.
Direct Link - Mazlan, A.G. and M. Rohaya, 2008. Size, growth and reproductive biology of the giant mudskipper, Periophthalmodon schlosseri (Pallas, 1770), in Malaysian waters. J. Applied Ichthyol., 24: 290-296.
CrossRefDirect Link - Miller, B.M., A.M. Clough, J.H. Batson and R.W. Vachet, 2006. Transition metal binding to cod otolith proteins. J. Exp. Mar. Biol. Ecol., 329: 135-143.
Direct Link - Milton, D.A. and S.R. Chenery, 2001. Sources and uptakes of trace metals in otoliths of juvenile barramundi (Lates calcarifer). J. Exp. Mar. Biol. Ecol., 264: 47-65.
Direct Link - Payan, P., H. De Pontual, G. Boeuf and N. Mayer-Gostan, 2004. Endolymph chemistry and otolith growth in fish. Comptes Rendus Palevol, 3: 535-547.
CrossRefDirect Link - Pritochard, D.W., 1955. Estuarine circulation patterns. Proc. Am. Soc. Civil Eng., 81: 1-11.
Direct Link - Romanek, C.S. and R.W. Gauldie, 1996. . A predictive model of otolith growth based on the chemistry of fish endolymph. Comp. Biochem. Phys., 114: 71-79.
Direct Link - Secor, D.H. and J.R. Rooker, 2000. Is otolith strontium a useful scalar of life cycles in estuarine fishes? Fish. Res., 46: 359-371.
Direct Link - Thorrold, S.R., C.M. Jones and S.E. Campana, 1997. Response of otolith microchemistry to environmental variations experienced by larval and juvenile Atlantic croacker (Micropogonias undulates). Limnol. Oceanogr., 42: 102-111.
Direct Link - Tytler, P. and T. Vaughan, 1983. Thermal ecology of the mudskippers, Periophthalmus koelreuteri (Pallas) and Boleophthalmus boddarti (Pallas) of Kuwait bay. J. Fish Biol., 23: 327-337.
CrossRefDirect Link - Leakey, C.D.B., M.J. Attrill and M.F. Fitzsimons, 2009. Multi-element otolith chemistry of juvenile sole (Solea solea), whiting (Merlangius merlangus) and European seabass (Dicentrarchus labrax) in the Thames Estuary and adjacent coastal regions. J. Sea Res.
Direct Link - De Vriesa, M.C., B.M. Gillanders and T.S. Elsdon, 2005. Facilitation of barium uptake into fish otoliths: Influence of strontium concentration and salinity. Geochimica et Cosmochimica Acta, 69: 4061-4072.
CrossRef - Strelcheck, A.J., G.R. Fitzhugh, F.C. Coleman and C.C. Koening, 2003. Otolith-fish size relationship in juvenile gag (Mycteroperca microlepis) of the eastern Gulf of Mexico: A comparison of growth rates between laboratory and field populations. Fish. Res., 60: 255-265.
CrossRef