
Cahide Gokkusu
Department of Biochemist1Y,
Istanbul Medical Faculty,
Istanbul University,
34093, �apa, Istanbul, Turkey
Tulin Ayse Ozden
Institue of Child Health, �apa 34093, Istanbul, Turkey
Tannaz Mostafa
Department of Biochemist1Y,
Istanbul Medical Faculty,
Istanbul University,
34093, �apa, Istanbul, Turkey
Journal of Medical Sciences
Year: 2004 | Volume: 4 | Issue: 1 | Page No.: 70-76







ABSTRACT
To examine the influence of vitamin E on oxidative stress and membrane ATPases. Present study conducted on hypercholesterolemic rats. The animals were divided into four equal groups. The control group was fed with chow pellets for 4.5 months. The other groups received high cholesterol or vitamin E or high cholesterol plus vitamin E. The high cholesterol and high cholesterol plus vitamin E groups showed a significant increase of total cholesterol of plasma, erythrocytes and liver and also an increase in plasma phospholipid and triglyceride levels was observed. But erythrocytes and liver phospholipid levels in high choesterol and high cholesterol plus vitamin E groups remained unchanged. There were no significant differences in plasma, erythrocytes and liver total cholesterol levels between high cholesterol and high cholesterol plus vitamin E groups. The plasma triglyceride level was significantly decreased in the high cholesterol plus vitamin E group in comparison to the high cholesterol group. On the other hand, lipid peroxide levels in the high cholesterol group significantly elevated in comparison with the control group. On the contrary, the high cholesterol plus vitamin E group showed a significant decrease in the lipid peroxide levels of plasma, erythrocytes and liver compared to that of high cholesterol group. The glutathione content was found to be significantly lower than the control group. Following vitamin E supplementation, glutathione content was significantly higher in plasma, erythrocytes and liver. The Na+-K+-ATPase and Ca+2-ATPase activities of erythrocytes and liver in high cholesterol group significantly decreased when compared with controls. On the other hand, high cholesterol plus vitamin E injection to the rats suppressed the reduction of Na+-K+-ATPase and Ca+2-ATPase activities in these tissue membranes. In conclusion, long-term administration of vitamin E may be useful in preserving of membrane asymmetry by suppressing the oxidative stress.
PDF Abstract XML References Citation
How to cite this article
Cahide Gokkusu, Tulin Ayse Ozden and Tannaz Mostafa, 2004. The Relationship Between Oxidative Stress and Membrane ATPase Enzyme Activities in Hypercholesterolemic Rats Supplemented with Vitamin E. Journal of Medical Sciences, 4: 70-76.
DOI: 10.3923/jms.2004.70.76
URL: https://scialert.net/abstract/?doi=jms.2004.70.76
DOI: 10.3923/jms.2004.70.76
URL: https://scialert.net/abstract/?doi=jms.2004.70.76
INTRODUCTION
Membrane components of the cell play an important role in the regulation of cell function. It has been shown that the lipid composition of membranes can modify by dietary fats[1-5]. It is in agreement with the fact that the activity of many membrane bound enzymes and transport systems is dependent on the physical state of the membrane lipid microenvironment[6]. ATPases are membrane-bound enzymatic proteins and they are sensitive to the alterations in the membrane lipid composition[6]. It has also been observed that an increase in the cholesterol content of plasma membranes results in decreased activities of ATPases[4,7].
According to response-to-injury theory of the genesis of atherosclerosis, endothelial cell damage is the basic mechanism for initiation and maintenance of atherosclerosis[8]. Despite considerable epidemiological evidence that hypercholesterolemia affects the development of atherosclerosis, the mechanism by which this happens is not clear. There is strong evidence that hypercholesterolemia increases production of free radicals (FRs) and leads to endothelial cell injury, which sets the stage for atherosclerosis[8,9]. FRs are clearly involved in pathogenesis of various diseases such as atherosclerosis, inflammatory diseases and cancer[10,11]. FRs have been suggested to exert their cytotoxic effects by causing peroxidation of membrane phospholipids (PL), which can results in an elevation in membrane fluidity, which increases permeability and loss of membrane integrity[12,13].
The evolution of oxidative processes within mammalian cells has necessitated the concomitant development of defensive mechanisms to protect vital cell components against oxidative stress. Oxidative stress, the disturbance of the delicate balance between oxidants and antioxidants, may result from increased production of free radicals and/or impaired antioxidant defense systems[14,15].
Vitamin E is an important lipid-soluble antioxidant placed in a special region of membranes. Recent evidences indicate that besides its antioxidant function, vitamin E may act as a modulator of PL metabolism in the mammalian tissues[16,17]. Deprivation of dietary vitamin E results in species-dependent and tissue-specific pathological lesions[18,19]. The studies carried on both experimental animals and human subjects have demonstrated that there is a negative correlation between cholesterol and vitamin E levels and vitamin E supplementation to the diet reduces the elevated serum cholesterol levels[16-20].
In the present study, we examined the effects of vitamin E supplementation on the lipid composition, the lipid peroxidation levels and glutathione (GSH) content in plasma, erythrocytes and liver of hypercholesterolemic rats. The activities of membrane ATPases in erythrocytes and liver cells were also determined in order to see whether vitamin E administration has any influence on cholesterol-induced changes in these tissues.
MATERIALS AND METHODS
Vitamin E was supplied from Roche Products Ltd. (Istanbul, Turkey). The standard diet was from Istanbul Diet Co. Ltd. (Istanbul, Turkey). All the other chemicals were from Sigma (St Louis, MO). Enzymatic kits were obtained from Randox (UK).
We used 48 male Sprauge Dawley rats weighing 180-200 g. Animals were supplied from DETAM (Istanbul, Turkey). The animals were initially fed the chow pellets for 15 days, after the adaptation, the rats were divided into four groups (12 rats each). The control group (Group I) fed basal diet. The basal diet was composed of 20% casein, 15% corn starch, 50% sucrose, 3.5% mineral mixture, 0.5% cellulose, 0.3% DL-methionine, 0.2% choline tartrate and 5% corn oil as previously described[15]. Rats in group II were received 280 U kg-1d-1 d, 1-alpha-tocopheryl acetate intraperitoneally (i.p) in addition to basal diet for 4.5 months. The hypercholesterolemic group (group III) fed basal diet plus 2% (w/w) cholesterol diet (2% cholesterol, 0.5% cholic acid and 2.5% propylthiouracil) as previously described[21,22]. Rats in group IV were received 280 U kg-1d-1 d, 1-alpha-tocopheryl acetate intraperitoneally (i.p) in addition to hypercholesterolemic diet for 4.5 months. Food consumption was determined every day for the experimental period. The animals were allowed water ad libitum. All the animals were sacrificed at the end of 4.5 months. Blood samples were collected into heparinized tubes by cardiac vein puncture. Plasma was separated from erythrocytes by centrifugation at +4°C. Livers were removed quickly, rinsed in ice-cold saline, dried on a filter paper and stored at -80°C until analyzed.
Measurement of plasma, erythrocyte and liver lipid compositions: Plasma, erythrocyte and liver lipids were extracted by the method of Folch et al.[23]. Enzymatic kits (Randox, UK) determined total cholesterol (TC) and triglyceride (TG) levels in these extracts. The total amounts of phospholipid (TPL) in the extracts were measured by the method of Gomori[24].
Measurement of TBARS levels: The thiobarbituric acid reacting substances (TBARS) levels in plasma, erythrocyte and liver were estimated by the method of Okhawa et al.[25]. Briefly, 0.2 ml 8.1% sodium dodecyl sulfate (SDS), 1.5 ml 0.8% thiobarbituric acid (TBA), 1.5 ml 20% glacial acetic acid and 0.6 ml distilled water were added and the mixture was kept in a water bath at 95oC for 60 min. the colored reaction product was extracted with 5 ml n-butanol / pyridine (15:1 v/v) and the absorbance at 532 nm was recorded. The breakdown product of tetra ethoxypropane was used as a standard. The results in plasma, erythrocyte and liver were expressed in nmol TBARS/ml, nmol TBARS/gHb and nmol TBARS/mg protein, respectively.
Measurement of GSH levels in erythrocyte and liver: SH-groups were assayed by the method of Ellman[26]. Briefly, 2 ml of erythrocyte suspensions and liver homogenate were deproteinized by adding of 3 ml deproteinization solution (NaCl, metaphosphoric acid, EDTA and distilled water). After centrifugation, 0.5 ml of supernatant was added to 2 ml 0.3 M Na2HPO4 2H2O and 0.2 ml solution of dithiobisnitrobenzoate (0.4 mg ml-1 in 1% sodium citrate) and the absorbance at 412 nm was measured immediately after mixing.
Measurement of activities of membrane ATPases: Washed erythrocytes were lysed with hypotonic 7 mM Tris-0.1 mM Na2-EDTA buffer, pH 7.4 and the membranes precipitated by centrifugation at 15.000xg for 20min at 4°C[24,25]. 10% tissue homogenates were prepared in 0.25 M sucrose, 0.5 mM CaCl2, 5 mM Tris-HCl, pH 7.4. Homogenates were centrifuged two times at 450 rpm for 10 min. at +2°C then the supernatants were centrifuged at 1800 rpm for 12 min. at +2°C. The precipitants were suspended (0.25M sucrose + 5 mM Tris-HCl, pH 7.4) and centrifuged two times at 3800 rpm for 10 min. at +2°C. The plasma membranes were resuspended in 5 ml ice-cold bidistiled water and used for the measurement of ATPases' activities.
| ● | Total ATPase activity was measured by a coupled optical test at 700 nm at 37°C as described in[27,28] with slight modifications. The 2.5 ml of assay medium contained standard buffer (25 mmol Tris-HCl, 75 mmol NaCl, 5 mmol KCl, 1mmol MgCl2, pH 7.4), 5 mmol CaCl2 and enzyme suspension. The reaction was started by addition of 5mM Na2-ATP. Total ATPase activity was expressed as μmol Pi /mg protein/h. |
| ● | Na+-K+-Mg+2 ATPase activity was determined at 37°C using 2.5 ml of an assay medium containing standard buffer (25 mmol Tris-HCl, 75 mmol NaCl, 5 mmol KCl, 1mmol MgCl2, pH 7.4), 2.5 mM EGTA and enzyme suspension. The reaction was started by addition of 5mM Na2-ATP. The absorbance changes were recorded at 700 nm. Na+-K+-Mg+2 ATPase activity was expressed as μmol Pi /mg protein/h. |
| ● | Mg+2 ATPase activity was determined at 37°C using 2.5 ml of an assay medium containing standard buffer (25 mmol Tris-HCl, 75 mmol NaCl, 5 mmol KCl, 1mmol MgCl2, pH 7.4), 2.5 mM EGTA, 25 mM Oubain and enzyme suspension. The reaction was started by addition of 5mM Na2-ATP. The absorbance changes were recorded at 700 nm. Na+-K+-Mg+2 ATPase activity was expressed as μmol Pi /mg protein/h. |
| ● | Ca+2-ATPase activity was calculated by subtracting Na+-K+-Mg+2 ATPase activity from total ATPase activity. |
Na+-K+-ATPase activity was calculated by subtracting Mg+2-ATPase activity from Na+-K+-Mg+2-ATPase activity.
Measurement of protein concentrations: The protein concentration of the ghost and liver homogenate was determined by the Lowry method[29].
Statistical analyses of the data were performed with Student's t-test.
RESULTS
The body weight of rats fed the hypercholesterolemic diet showed a mean value of 171.2±42.2 while those animals fed the basal diet had a mean weight of 245.9±38.8 (results no shown).
The lipid levels of plasma, erythrocytes and liver homogenate at the end of the experiment are shown in Table 1. The HC and HC plus vitamin E groups showed a significant increase of total cholesterol (TC) of plasma, erythrocytes and liver and also an increase in plasma phospholipid (PL) and triglyceride (TG) levels was observed. But erythrocytes and liver PL levels in HC and HC plus vitamin E groups remained unchanged. There were no significant differences in plasma, erythrocytes and liver homogenate TC levels between HC diet and HC diet plus vitamin E groups. The plasma TG level was significantly decreased in the HC diet plus vitamin E group in comparison with the HC diet group.
On the other hand, TBARS level in the HC diet group significantly elevated (1.2 and 1.3 fold, respectively) in comparison to the control. On the contrary, the HC diet plus vitamin E group showed a significant decrease (2.6, 1.3 and 1.1 fold, respectively) in the TBARS level of plasma, erythrocytes and liver homogenate compared to that of HC diet group (Table 2). The GSH content was found to be significantly lower than the control group (56% and 80%, respectively). Following vitamin E supplementation, GSH content was significant higher in plasma, erythrocytes and liver homogenate.
| Table 1: | Effects of vitamin E on plasma, erythrocyte and liver lipid levels in hypercholesterolemic rats (Means±SD) |
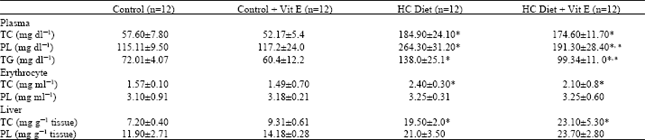 | |
| *p<0.001 when HC and HC plus vitamin E groups were compared with the control group ap<0.001when HC group was compared with HC plus vitamin E group | |
| Table 2: | Effects of vitamin E on plasma, erythrocyte and liver TBARS levels and GSH content in hypercholesterolemic rats (Means±SD) |
 | |
| *p<0.001 when HC and HC plus vitamin E groups were compared with the control group a p<0.001, b p<0.01 when HC group was compared with HC plus vitamin E group | |
| Table 3: | Effects of vitamin E on erythrocyte and liver plasma membrane ATPase activities (μmol Pi/ mg protein/ h) in hypercholesterolemic rats (Means±SD) |
 | |
| *p<0.001, **p<0.01 when HC and HC plus vitamin E groups were compared with the control group ap<0.001when HC group was compared with HC plus vitamin E group | |
HC administration to the rats for 4.5 months, significantly decreased erythrocytes and liver plasma membrane Na+-K+-ATPase and Ca+2-ATPase activities when compared with controls. On the other hand, HC plus vitamin E injection to the rats suppressed the reduction of Na+-K+-ATPase (p<0.001 and p<0.001, respectively) and Ca+2-ATPase activities (p<0.001 and p<0.001, respectively) in these tissue membranes (Table 3).
DISCUSSION
In this study, the antioxidant effects of vitamin E were examined in plasma; erythrocytes and liver in rat fed a high cholesterol diet. During the experimental procedure, significant reductions in body weight of animals were observed, which may be attributed to the anorectic effect of cholic acid in diet[30].
As previously reported, cholesterol-rich diet caused significant increases in the plasma, erythrocyte and liver TC, plasma TG and TBARS levels, but no significant differences was observed in PL levels, except plasma in this study. High-cholesterole diet also decreased the Na+-K+-ATPase and Ca+2-ATPase activities in the erythrocyte and liver as well as glutathione (GSH) levels in plasma, erythrocyte and liver of the rats in comparison with the controls. Following the high-cholesterole diet, the reduction in the activities of the ATPases may be due to enhanced oxidation of membrane lipids and proteins. Our results are in accordance with the pervious reports[4,6,40].
It is known that vitamin E is a lipophilic antioxidant and its primary role is to prevent lipid peroxidative damage in the tissues[1,15]. A stabilizing role of vitamin E for PLs in membranes has also been reported[4]. Although it is suggested that vitamin E supplementation to diet may prevent the elevation of serum lipids, there are some controversial reports about this subject[19,20]. Viswanathan et al.[31] have reported an increase in serum triglycerides, a decrease in LDL-C and no change in serum TC in rabbits that treated with vitamin E. Komaratat et al.[32] observed a decrease in total TC, LDL-C and VLDL-C and an increase in HDL-D with increasing doses of vitamin E in rabbits. In another study, Prasad et al.[8] reported that 0.04g kg-1 vitamin E administration to rabbits did not change the lipid profile of rabbits. These differences in results could be due to the differences in the doses of vitamin E. In the present study, there were no significant differences in plasma, erythrocytes and liver homogenate TC levels between HC diet and HC diet plus vitamin E groups. Our results are in accordance with the results of the other reports[8,15].
On the other hand, evidence suggests that the hypercholesterolemia could increase levels of oxygen free radicals in various ways[12,33,34]. Hypercholesterolemia-induced atherosclerosis and endothelial impairment by free radical formation were markedly suppressed after treatment with vitamin E. Our previous studies have shown that TBARS levels of plasma and tissues homogenates are increased in hypercholesterolemia atherosclerosis[15]. Prasad et al.[8] have reported a decrease in the blood TBRS levels in rabbits that supplemented with vitamin E. Szczeklik et al.[35] observed no change in plasma TBARS concentrations in vitamin E supplemented rabbits on a high-cholesterol diet. Our data indicate that plasma, erythrocyte and liver TBARS levels as markers of oxidative stress decreased in HC diet plus vitamin E group. The prevented effect on the TBARS levels of vitamin E in hypercholesterolemic rats may be trapping the chain-propagating peroxyl radicals. Our findings are in accordance with the results of the other reports[8,15,36].
Plasma thiols are physiological free radical scavengers and may serve an antioxidant function by several mechanisms. They may preemptively scavenge oxidants that initiate peroxidation, thus sparing vitamin E and/ or lipids from attack[37]. As markers of free radical scavenging, GSH may be a primary agent involved in redox regulation of protein thiols[38]. In our study, increases in plasma, erythrocytes and liver GSH content were observed in rats treated with 280 U kg-1 bw daily doses of vitamin E for 4.5 months. Similar results were obtained in human and animals studies[15,35].
On the other hand ATPases are membrane bound enzymatic proteins and serve in the maintenance of the ionic gradients between aqueous intra-and extracellular phases[3,4]. ATPases need phospholipids for their activities and increased levels of membrane CHO cause substantial modifications in the ordering of the phospholipids[39,40]. Several studies showed that activity of ATPases is inhibited in the presence of long chain fatty acids and cholesterol controls the membrane lipid order, which may be a factor in the activation of membrane transport systems[41,42,43]. In this study, vitamin E supplementation to diet significantly increased the tissue ATPase activities in comparison to hypercholesterolemic rats. Our data demonstrate that vitamin E protects ion pump ATPases from hypercholesterolemia mediated inhibition, presumably by limiting the degree of oxidation and levels of oxidation by products, capable of cross-linking membrane proteins.
In conclusion, long-term administration of vitamin E may play an important role in preserving of membrane asymmetry by suppressing the oxidative stress and may be useful for the prevention and/or early treatment of hypercholesterolemia.
ACKNOWLEDGMENTS
Istanbul University Research Fund supported this study.
REFERENCES
- Kehrer, J.P., 1993. Free radicals as mediators of tissue injury and disease. Crit. Rev. Toxicol., 23: 21-48.
CrossRefPubMedDirect Link - Gokkusu, C. and T. Mostafazadeh, 2003. Changes of oxidative stress in various tissues by long-term administration of vitamin E in hypercholesterolemic rats. Clinica Chimica Acta, 328: 155-161.
CrossRefDirect Link - Chow, C.K., 1991. Vitamin E and oxidative stress. Free Radic. Biol. Med., 11: 215-232.
CrossRefDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 82: 70-77.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Komaratat, P., N. Chupukcharoen and P. Wilairat, 1985. Effect of vitamin E on cholesterol plasma lipoprotein distribution and metabolism in rabbit. Int. J. Vitamin Nutr. Res., 55: 167-171.
Direct Link - Halliwell, B., 1994. Free radicals, antioxidants and human disease: Curiosity, cause, or consequence? Lancet, 344: 721-724.
CrossRefPubMedDirect Link - Sies, H., 1999. Glutathione and its role in cellular functions. Free Radic. Biol. Med., 27: 916-921.
CrossRefPubMedDirect Link