
Ekachai Chukeatirote
Department of Biotechnology, School of Science, Mae Fah Luang University, Chiang Rai 57100 Thailand
Chularat Chainun
Department of Biotechnology, School of Science, Mae Fah Luang University, Chiang Rai 57100 Thailand
Apiradee Siengsubchart
Department of Biotechnology, School of Science, Mae Fah Luang University, Chiang Rai 57100 Thailand
Churairat Moukamnerd
Department of Biotechnology, School of Science, Mae Fah Luang University, Chiang Rai 57100 Thailand
Panuwan Chantawannakul
Department of Biology, Faculty of Science, Chiang Mai University,
Chiang Mai 50200 Thailand
Saisamorn Lumyong
Department of Biology, Faculty of Science, Chiang Mai University,
Chiang Mai 50200 Thailand
Nitaya Boontim
Institute for Science and Technology Research and Development, Chiang Mai University, Chiang Mai 50200 Thailand
Primprao Thakang
Department of Biotechnology, School of Science, Mae Fah Luang University, Chiang Rai 57100 Thailand
Research Journal of Microbiology
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 38-44







ABSTRACT
In this study, we aimed to investigate the microbiological and biochemical changes during the fermentation. It was showed that microbial population (bacteria and fungi) and pH increased with fermentation time. The bacterial groups isolated were further characterised and found to be Bacillus sp.; however, some Lactobacillus sp. and unidentified cocci were also detected. In addition, some chemical properties (i.e., pH and ammonia concentration) and enzymatic activities were determined. Proteases and amylases activities increased rapidly reaching a peak at 60 h (100% relative activity) and at 72 h (1,380 U), respectively. Phytase activity was also detected varying from 114.7-152.6 U. In contrast, lipases activity was minimal throughout the fermentation (0.78-1.31 U).
PDF Abstract XML References
How to cite this article
Ekachai Chukeatirote, Chularat Chainun, Apiradee Siengsubchart, Churairat Moukamnerd, Panuwan Chantawannakul, Saisamorn Lumyong, Nitaya Boontim and Primprao Thakang, 2006. Microbiological and Biochemical Changes in Thua nao Fermentation. Research Journal of Microbiology, 1: 38-44.
URL: https://scialert.net/abstract/?doi=jm.2006.38.44
URL: https://scialert.net/abstract/?doi=jm.2006.38.44
INTRODUCTION
Thai fermented soybeans (the so-called Thua nao named after its strong and pungent odour during fermentation) have been traditionally produced and consumed by people in the Northern part of Thailand and in the Shan State of Myanmar for centuries (Sundhagul et al., 1972; Chantawannakul, 1995). This protein-rich product is generally used as meat substitute and condiment in local food. To make Thua nao, whole soybeans are cleaned, washed and soaked in tap water overnight. Soaked beans are then cooked by boiling for 3-4 h (or until the soybeans are soft enough to crush by fingers) and excess water is drained off. Cooked soybeans are air-dried, allowed to cool down before putting in a bamboo basket lined and covered with banana leaves and left to ferment naturally for 2-3 days. These fermented soybean products are then cooked by steaming or roasting prior to consuming. Alternatively, the fermented products could be smashed, formed into a thin disc (8-10 cm diameter) and exposed to sunlight resulting in a dried form of Thua nao that can be stored for 3-4 months. Its use is versatile; fresh Thua nao, for example, could be seasoned with spices and consumed with sticky rice whereas the dried Thua nao are used in various kinds of local dishes.
Thua nao which is similar to Indian kinema and Japanese natto is a type of non-salted fermented soybean products. There are also other related products in which legume seeds are used rather than soybeans such as ugba and iru of Western Africa. Of these, only a few of them (i.e., natto, kinema and daddawa) have been characterised systematically and manufactured commercially. Presently, industrial production of natto has been developed and established; these include a use of pure bacterial starter culture and quality-controlled processing line (Hosoi and Kiuchi, 2003).
Several Bacillus species have been found predominantly in these fermented soybean products. For example, in Nigerian daddawa a group of Bacillus species including B. subtilis, B. pumilus and B. licheniformis could be isolated (Ogbadu and Okagbue, 1988; Omafuvbe et al., 2000). Similarly, the Bacillus species (i.e., B. subtilis, B. licheniformis and B. badius) was also dominant in kinema and cembalo (Sarkar et al., 1994; 2002). It should also be noted, however that other bacterial species (i.e., lactic acid bacteria or Enterococcus species) as well as some certain yeast strains may exist (Sarkar et al., 1994; Chukeatirote, 1993). The abundant presence of Bacillus species is possibly due to the boiling effect from initial preparation. The occurrence of non-Bacillus species is likely derived from contamination during and/or after the fermentation process. Furthermore, the beneficial effect of Bacillus fermentation on these fermented soybean products has been widely acknowledged including the presence of anti-mutagenic and anti-oxidative agents as well as some proteolytic enzymes (Fujita et al., 1993; Osawa and Matsumoto, 1997; Minamiyama et al., 2003; Park et al., 2003; Ren et al., 2005).
In contrast, current status of Thua nao production still remains at traditional small-scale household basis with little data regarding microbial population and fermentation process. Previous work confirmed that Bacillus species were also predominant in Thua nao products (Chantawannakul et al., 2002). In this study, the information on the microbial community and biochemical changes that occurred during the Thua nao fermentation is reported.
MATERIALS AND METHODS
Thua nao was prepared conventionally by a local housewife group in Chiang Rai between October and November 2004. Briefly, the overall procedure was as follows: 1 kg of soybean was washed, soaked overnight, cooked for 3 h and allowed to ferment in a bamboo basket lined with banana leaves for 72 h at ambient temperature. Thua nao samples were then collected every twelve hours for further analysis.
Thua nao samples were characterised for their chemical properties as follows: pH, ammonia concentration, amount of reducing sugars and total proteins. Typically, 10.0 g of Thua nao samples were mixed with 90 mL of 0.85% (w/v) NaCl and subsequently centrifuged to collect the supernatant. The pH was determined using the digital pH meter (Sartorius). Ammonia concentration was measured by titration with 0.1 M HCl (AOAC, 1990) after steam distillation with Kjeltec 2100 (Foss Tecator). Amount of reducing sugar was estimated from a standard curve of glucose solution using the DNS method (Miller, 1959). Total protein content was calculated based on a standard graph of the BSA solution using the Lowry method (Lowry et al., 1951).
A sample (10.0 g) was mixed with 90 mL of 0.85% (w/v) sterile physiological saline and decimal dilution series was performed. Aliquots (0.1 mL) of appropriate dilutions (usually between 10-4-10-8) were plated in triplicate on nutrient agar for bacteria enumeration and potato dextrose agar supplying with 50 μg mL-1 of ampicillin and 100 μg mL-1 of chloramphenicol for fungi enumeration. The plates were then incubated at 37°C for 48 h for bacteria and at 30°C for 5 days for fungi. Microbial colonies were then counted and expressed as colony forming units per gram of the sample. Representative colonies from the plate counts were then purified by repeated streaking and subsequently characterised by the following standard methods: cell shape, Gram staining, presence of spore, catalase test, oxygen requirement, growth in the presence of NaCl and different temperatures, VP test, starch hydrolysis test and citrate test (MacFaddin, 2000).
The extracellular enzymes in the fermenting soybean (10 g) were extracted with 90 mL of 0.85% (w/v) NaCl. The soybean debris was then removed by centrifugation (14000 rpm, 10 min, 4°C) and the supernatant was collected and kept on ice until use.
To determine α-amylase activity (Calzyme, 2000), 0.5 mL of the extracting enzymes were initially incubated at 25°C for 5min and the reaction was begun by addition of 0.5 mL of 1% (w/v) starch solution; the reaction was then proceeded at 25°C for 3 min. The amylase activity was then assayed by determining amount of the reducing sugars using the DNS method. One unit of amylase activity was defined as amount of the enzyme which produced one micromole of glucose under experimental conditions.
For protease activity, the assay was essentially performed as described by Brock et al. (1982). Twenty microlitres of the crude enzymes were incubated at 37°C with a mixture of 230 μL of 2% azocasein and 150 μL of 0.2 M N-tris[hydroxymethyl]methyl-2-aminoethanesulfonic acid (TES) buffer pH 7.0 for 2 h. To stop the reaction, 1.2 mL of 10% trichloroacetic acid (TCA) was added. All samples were then incubated at room temperature for 15 min and subsequently centrifuged at 10,000 rpm at 4°C for 5 min. The supernatant (~1.0-1.2 mL) was collected and transferred to a new tube containing an equivalent volume of 1 M NaOH. The solution was finally mixed well prior to measuring the absorbance at 440 nm. The protease activity was then expressed as relative activity of the absorbance values obtained.
Lipase activity was also determined using p-nitrophenyl laurate (p-NPL) as a substrate (Sugihara et al., 1991). For this, 20 μL of the crude enzymes were added into 800 μL of the reaction buffer (50 mM Tris-HCl (pH 8.0), 0.1% arabic gum and 0.2% deoxycholate). The reaction was then initiated by addition of 100 μL of 8 mM p-NPL and incubated at 55°C for 3 min. To terminate the reaction, 0.5 mL of 3 M HCl was added. After centrifugation, 333 μL of the supernatant was mixed with 1 mL of 2 M NaOH and the OD420 was measured. One unit of the enzyme activity was described as the enzyme amount which produced 1 μmol of p-nitrophenol per millitre per minute.
For phytase activity (Fiske and Subbarow, 1925), a reaction mixture consisting of 100 μL of enzyme solution and 400 μL of 2 mM sodium phytate was prepared and subsequently incubated at 50°C for 30 min. To stop the reaction, 500 μL of 15% TCA was added. Four millitres of colouring reagent (1:1:1:2 ratio of 6 N H2SO4 : 2.5% ammonium molybdate : 10% ascorbic acid : H2O) were then added and the samples were kept at 50°C for 30 min prior to measuring the absorbance at 820 nm. One unit of phytase activity was defined as amount of enzyme used to liberate 1 μmol of phosphorus per minute under the assay condition.
RESULTS AND DISCUSSION
In this study, five batches of Thua nao were prepared traditionally by the local housewife group in Chiang Rai. To investigate the microbial succession during fermentation, Thua nao samples were collected every 12 h, subjected to ten-fold serial dilutions and 0.1 mL of the diluted samples were then transferred to the media plates using spread plate technique. As shown in Fig. 1, the total viable count of bacteria and fungi tended to increase with fermentation time. It was found that their population increased rapidly in the first 36 h of fermentation and the rates did not change much after that. Overall, the bacterial population exceeded that of fungi and reached its peak (2.88 x 1013 CFU g-1) at 72 h whereas the highest total count of fungi population was 3.58 x 1012 CFU g-1 at 48 h.
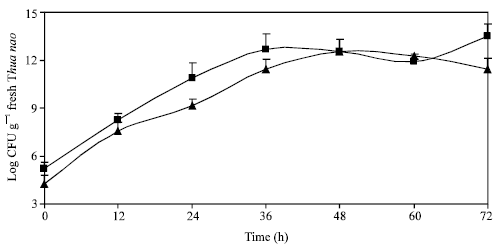 | |
| Fig. 1: | Changes of microflora during Thua nao fermentation: total count of bacteria (■) and fungi (▲). Mean of triplicates; bars indicate range |
| Table 1: | Occurrence of some bacterial species in Thua nao fermentation |
 | |
| +: Present -: Absent | |
Existence of fungi might cause a serious concern about food safety. Fortunately, Thua nao products were typically cooked again by steaming or roasting before consuming and thus should prevent this harm. It would also be of great importance to further study if the fungi present were mycotoxin-producing strain or not.
In addition, bacterial colonies obtained during the fermentation were randomly selected from different periods of time. They were restreaked and subjected to further analyses for Genus and species identification. These analyses included cell morphology, catalase test, Voges Proskaur (VP) assay, citrate utilisation, oxygen requirement, growth in different concentration of NaCl and starch hydrolysis. By these approaches, bacteria were identified as B. subtilis, B. pumilus and Lactobacillus sp. (Sneath, 1986). The occurrence of these bacterial isolates is shown in Table 1. It was found that Bacillus species were predominant especially B. subtilis which predominated throughout the 72 h fermentation. Present results are in accordance with those of Sarkar et al. (1994) and Omafuvbe et al. (2000) which reported the predominance of Bacillus species in kinema and daddawa. Lactobacillus spp. was present at the onset (12-24 h) and positively contributed to the lactic acid production in Thua nao products (Chukeatirote, 1993; Boontim et al., 2003). Micrococcus luteus, Staphylococcus epidermidis and Escherichia coli were absent however there existed of Gram positive cocci and Gram negative rod which remained as yet to be identified. This result is contrary to that of Omafuvbe et al. (2000) which reported the presence of S. epidermidis and M. luteus. However, the existence of both bacteria was limited to at 0 h and thus may not involve in the fermentation.
| Table 2: | Changes in physical, chemical and enzyme activities during Thua nao fermentation |
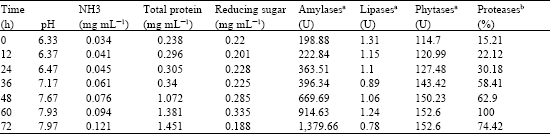 | |
| aThe enzyme unit was used to express the activities of amylases, lipases and phytases; one unit of enzyme activity was described as the enzyme amount which produced 1μmol of products per millitre per minute, bFor proteases, the relative activity (100%) was used instead. | |
pH, NH3 and total protein increased as the fermentation proceeded (Table 2). The increase of pH was due to proteolysis and the release of NH3 following the utilisation of amino acids by the fermenting microbes. High amount of NH3 was produced and caused strong ammoniacal smell resulting in the distinct feature of Thua nao products. The level of sugar was nearly constant from 0-36 h period with abrupt change between 60-72 h. The initial increase of reducing sugars was possibly due to the rise of amylase enzymes and its decrease at the end period was presumably caused by the increasing microbial population. The rise of total protein was also expected as an increasing microbial population would secrete extracellular enzymes to degrade macronutrients for food and energy. This occurrence was in agreement with the increase of these amylases, phytases and proteases but not lipases enzyme activities determined (Table 2).
Lipase activity was also detected with inconsistent result ranging from 0.78-1.31 U. The low lipase activity was unexpected considered that soybean is an oil-rich seed. However, similar observations have been described with other oil-rich seed fermentation (Omafuvbe et al, 2000; Odibo et al., 1990). These results including ours suggest that activity of lipase may not be significant in the soybean fermentation. In addition, the Bacillus species which are ubiquitous in such fermentations are not known to be good lipase producers (Aderibigbe and Odunfa, 1990). Activity of phytases which are important for phytate degradation in soybean seeds was also detected varying from 114.7-152.6 U. The presence of phytase enzymes derived from the fermenting microbes are beneficial by transforming various kinds of fermented soybeans readily edible. Protease activity has previously been reported to be abundant in the fermentation of protein-rich foods (Sarkar et al., 1993). In this study, the protease activity was determined using the azocasein method (Brock et al., 1982) and their activity was then expressed as relative activity. It was shown that the activity of proteases increased and reached its peak (100%) at 60 h. The gradual increase of proteases also helped generate amount of NH3 from proteolysis reaction.
This study has shown that production of Thua nao is caused by microbial fermentation. The responsible bacterial species belong to the Genus Bacillus whereas the fungi present are being studied for their relevance in the fermentation. Several enzyme activities were observed and important for macronutrients hydrolysis resulting in increased soluble products.
ACKNOWLEDGMENTS
The authors would like to thank the Microbial Pathogen Unit of the Chiang Rai Province Hospital for detailed analysis of some microbial strains. This work is financially supported by the MFU (RMFU07/2547).
REFERENCES
- Brock, F.M., C.W. Forsberg and J.G. Buchanan-Smith, 1982. Proteolytic activity of rumen microorganisms and effects of proteinase inhibitors. Applied Environ. Microbiol., 44: 561-569.
Direct Link - Chantawannakul, P., A. Oncharoen, K. Klanbut, E. Chukeatirote and S. Lumyong, 2002. Characterisation of proteases of Bacillus subtilis strain 38 isolated from traditionally fermented soybean in Northern Thailand. ScienceAsia, 28: 241-245.
Direct Link - Fiske, C.H. and Y. Subbarow, 1925. The colorimetric determination of phosphorus. J. Biol. Chem., 66: 375-400.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Odibo, F.J.C., E. Nwabunnia and D.Z. Osuigwe, 1990. Biochemical changes during fermentation of Telfairia seeds for ogiri production. World J. Microbiol. Biotechnol., 6: 425-427.
CrossRefDirect Link - Ogbadu, L.J. and R.N. Okagbue, 1988. Bacterial fermentation of soybean for daddawa production. J. Applied Bacteriol., 65: 353-356.
CrossRef - Omafuvbe, B.O., O.O. Shonukan and S.H. Abiose, 2000. Microbiological and biochemical changes in the traditional fermentation of soybean for soy-daddawa-Nigerian food condiment. Food Microbiol., 17: 469-474.
CrossRefDirect Link - Sarkar, P.K., P.E. Cook and J.D. Owens, 1993. Bacillus fermentation of soybeans. World J. Microbiol. Biotechnol., 9: 295-299.
CrossRefDirect Link - Sarkar, P.K., J.P. Tamang, P.E. Cook and J.D. Owens, 1994. Kinema-a traditional soybean fermented food: Proximate composition and microflora. Food Microbiol., 11: 47-55.
CrossRef - Sarkar, P.K., B. Hasenack and M.J.R. Nout, 2002. Diversity and functionality of Bacillus and related genera isolated from spontaneously fermented soybeans (Indian Kinema) and locust beans (African Soumbala). Int. J. Food. Microbiol., 77: 175-186.
Direct Link - Ren, H., H. Lir, H. Endo, Y. Takagi and T. Hayashi, 2005. Anti-mutagenic and anti-oxidative activities found in Chinese traditional soybean fermented products furu. Food Chem., 95: 71-76.
Direct Link - MacFaddin, J.F., 2000. Biochemical Tests for Identification of Medical Bacteria. 3rd Edn., Lippincott Williams & Wilkins, Pennsylvania, United States, ISBN: 0-683-05318-3, Pages: 912.
Direct Link - Park, K., K. Jung, S. Rhee and Y.H. Choi, 2003. Antimutagenic effects of doenjang (Korean fermented soypaste) and its active compounds. Mutat. Res., 523-524: 43-53.
Direct Link