
Carbajal-Fajardo Zuleica Shareet
Laboratorio de Ecologia, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los Barrios, No. 1, C. P. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de Mexico, Mexico
Franco-Lopez Jonathan
Laboratorio de Ecologia, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los Barrios, No. 1, C. P. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de Mexico, Mexico
Hector Barrera Escorcia
Laboratorio de Ecologia, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los Barrios, No. 1, C. P. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de Mexico, Mexico
Luis Gerardo Abarca Arenas
Instituto de Investigaciones Biologicas-Universidad Veracruzana, Dr. Luis Castelazo s/n, Col. Industrial animas, Xalapa, Ver, Mexico
Carlos Bedia Sanchez
Laboratorio de Ecologia, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los Barrios, No. 1, C. P. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de Mexico, Mexico
Angel Moran Silva
Laboratorio de Ecologia, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los Barrios, No. 1, C. P. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de Mexico, Mexico
Horacio Vazquez-Lopez
Laboratorio de Ecologia, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autonoma de Mexico. Av. de los Barrios, No. 1, C. P. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de Mexico, Mexico
Journal of Fisheries and Aquatic Science
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 75-89







ABSTRACT
The objective of this study was to analyze the seasonal behavior of the icthyofauna, considering the trophic relationships between the species that live in Camaronera Lagoon, their food composition, diversity and trophic amplitude, as of the captures obtained in Camaronera Lagoon inlet during February-June, 2000. We realized seasonal tables of the fish species and items of identified foods. The data were grouped in diurnal and nocturnal samplings in order to know the abundance, specific richness, evenness and McNaughton’s community dominance index. The amplitude of the niche was calculated as of Levin’s standardized index. Costello’s graphic method was used to evaluate the preference and feeding behavior of any given type. The families with more species were Cichlidae, Eleotridae and Gobiidae. The most representative species regarding biomass and abundance were Gambusia affinis, Petenia splendida, Cathorops melanopus, Diapterus auratus and Bathygobius soporator.
PDF Abstract XML References Citation
How to cite this article
Carbajal-Fajardo Zuleica Shareet, Franco-Lopez Jonathan, Hector Barrera Escorcia, Luis Gerardo Abarca Arenas, Carlos Bedia Sanchez, Angel Moran Silva and Horacio Vazquez-Lopez, 2009. Trophic Seasonal Behavior of the Icthyofauna of Camaronera Lagoon, Veracruz. Journal of Fisheries and Aquatic Science, 4: 75-89.
DOI: 10.3923/jfas.2009.75.89
URL: https://scialert.net/abstract/?doi=jfas.2009.75.89
DOI: 10.3923/jfas.2009.75.89
URL: https://scialert.net/abstract/?doi=jfas.2009.75.89
INTRODUCTION
The estuarine ecosystems are one of the most productive environment concerning aquatic ecosystems; their extension comprises 240x105 km2 in tropical and subtropical coasts. They are characterized by their great number of primary producers, their high diversity of microhabitats and multiple and complex interactions, as well as an intense exchange of organic matter and organisms inside and outside the ecosystem (Day et al., 1989; Twilley et al., 1992). Mexico has a great extension of littorals that, as a whole, represent an extension of 11 500 km, with a wide exclusive economic area of 3 millions of km2 (Arreguín, 2006). In this extension, Mexico possesses between 30 and 35% of estuaries and coastal lagoons in the Pacific, Caribbean Sea and the Gulf of Mexico (Yañez Arancibia, 1986) that represent 1.5 million ha of estuarine environments and real or potentially productive coastal lagoons surface (Arreguín, 2006).
These ecosystems constitute ecological zones of great importance due to the magnitude of the interaction processes that are carried out between the intertidal zones and their adjacent areas; this can be noticed in the variety of natural components that these places have. Among others, we can emphasize the inter-lagoon zones that constitute areas that are productive enough and have favorable conditions for the immigration or recruitment of fish larvae (Richards and Vásquez, 1996).
The composition of the fish communities in coastal lagoons and estuaries varies in terms of the icthyofauna that each place has, as well as in those that have some mechanisms to tolerate the hydrological conditions of the system. This way, the existence of a wide range of species with a high complexity has been recognized in lagoons and estuaries; this complexity has to be understood, in order to exploit it and to conserve it rationally (Flores-Coto and Álvarez, 1980). Most of the tropical coastal fish species with economic importance depend, partially or totally, of estuarine waters that are used by the juvenile stages to complete their development until the mature age. The juveniles take advantage of the high primary production of the estuaries, which are more productive that the fresh waters or adjacent marine waters. We can thereby recognize that a high percentage of the coastal fish spawns in the sea and its larvae migrate or are transported though the inlets to the lagoon systems searching food and refuge (Flores-Coto et al., 1986; Franco and Chávez, 1993; Roman et al., 2006).
This dynamics allows to the coastal area (including lagoon systems), to maintain a balance between the generation and nutrient recycling, which are of great ecological and socioeconomic importance (Post and Lundin, 1996). Additionally, from a functional point of view, it is recognized that one of the main ecological roles of the fishes, is to control the specific and numeric structure of consumers through competition and depredation, as well as to transport and contribute with the energetic flow between the ecosystems habitats and limits (Yañez-Arancibia and Nugent, 1977). These environments are under a growing risk due to inadequate handling of human activities, such as pollution, habitats’ destruction and overexploitation of the resources (Post and Lundin, 1996.)
For the above-mentioned, it is necessary to increase the knowledge of these environments and their biological components. This study was carried out with the objective of analyzing the trophic dynamics of the icthyofauna existing in the Camaronera Lagoon, through the analysis of nictimeral cycles in the inlet of this lagoon and patterns of spatial and temporal variation of the species of this group.
MATERIALS AND METHODS
The Camaronera lagoon is located in the Gulf of Mexico coastal plain, south of the Port of Veracruz (18° 50’ at 18° 52’ 30’’ N, 95° 54’ at 95° 58’ 30’’ W) (Contreras and Castañeda, 1993) (Fig. 1).
The organisms were captured using a rowing boat 50 m long, 2.5 m high and 2.5 cm mesh, in the Camaronera Lagoon inlet during February-June 2000, covering the dry and rainy seasons and another cold season dominated by northward winds (hurricanes season known in Mexico as nortes season). In each season, a cycle of 24 h was carried out with samplings every 4 h, beginning at 19:00 h and ending at 15:00 h. The fish were fixed in situ, injecting the abdominal cavity with formaldehyde 10% with the finality of stopping the digestive processes.
The organisms were identified according to Jordan and Evermann’s (1886-1900), Fischer (1978), Castro-Aguirre (1999) and Reséndez (1981). Each specimen was measured with a conventional ictiometer of 50 cm ( ± 0.05) and then weighted using a digital scale with a capacity of 2.6 kg ( ± 0.05).
The area swept by the fishing art had an average estimation of 1200 m2 for each throw. In this way, the obtained results of fish abundance and biomass of fish are expressed in terms of density (ind m-2) and biomass (g m-2).
The organisms were dissected in order to examine the stomach contents. The percentage of stomachs examined for each one of the species oscillated between 30 and 100%.
From the compiled data of stomach contents analysis of the captured icthyofauna, seasonal tables of the fish species and of the items of identified foods were elaborated.
In order to know the abundance, specific richness (S), Pielou’s evenness index:
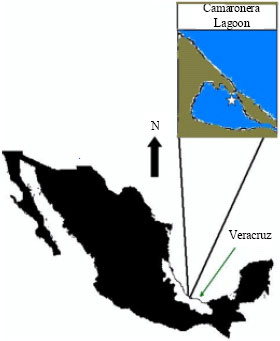 | |
| Fig. 1: | Study area |
were, H´ is the diversity by Shannon Wiener index and H´max is the maximum diversity or Log (S) and McNaughton’s community dominance index, the Community Analysis computer program (ANACOM) was used (De la Cruz, 1994).
The niche amplitude (BA) was calculated using Levin’s standardized index (Krebs, 1989), according to the equation:
Where:
| pj | = | The diet proportion comprising the prey j species |
| n | = | The total number of the prey’s species |
The obtained value represents the niche amplitude for the species; it varies from 0 to 1, where low values indicate that the feeding is dominated by few preys.
In order to evaluate the preference and nutritious behavior in any given type, the graphic method of Costello was employed, (Marshall and Elliott, 1997), where the occurrence percentage and weight percentage of each feeding type is applied. The occurrence percentage was graphed against the weight percentage, where the elements are recognized with regard to their position in the graph.
RESULTS
Environmental Parameters
During the dry season, the mean temperature was of 27.8 °C, with minimum and maximum temperatures of 25 and 31 °C, respectively; the salinity was of 16.1‰, with a variation between 14 and 18‰; the dissolved oxygen presented values between 8 and 12 ppm, registering a mean concentration of 9.83 ppm. In the rainy season, the mean temperature was of 29.5 °C, with a minimum temperature of 27 °C, reaching a maximum of 33 °C. The salinity increased up to a mean of 18.5‰, being the lowest in 16 ‰ and registering the highest value in the year, 21‰, the oxygen concentration was of 8.16 ppm, varying between 7 and 9 ppm in the captures (Table 1). Nortes season showed a mean temperature of 24.5 °C, with maximum temperature of 29 °C and minimum temperature of 22 °C. The mean salinity was 12‰, with minimum value of 11 and maximum of 21‰; the mean of dissolved oxygen was of 8.9 ppm, with values between 6.8 and 11 ppm.
Abundance and Biomass
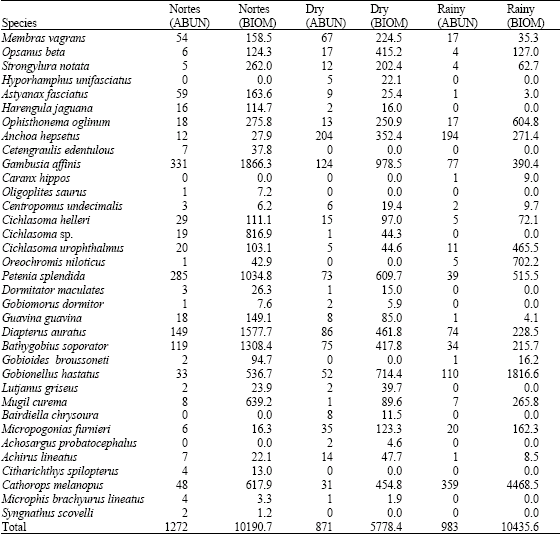
A total of 3126 individuals were captured with a biomass of 26404.7 g, belonging to 35 species of 22 families (Table 1). The dominant species according to McNaughton´s index for abundance and biomass in diurnal and nocturnal hours for each season correspond to the five species that provided between 65% and 84% of the total records (Table 2). The abundance and biomass records were greater in the rainy and nortes seasons, the abundances and biomasses being greater in diurnal hours than in nocturnal hours (Fig. 2).
The behavior of the abundance and biomass obtained in each season, shows that the records are higher in diurnal hours (07:00 to 15:00) than in nocturnal hours (19:00 to 03:00) (Fig. 3, 4).
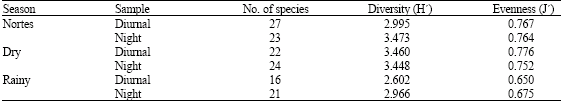
The records of specific richness, diversity and evenness calculated for each season in diurnal and nocturnal hours, help to recognize that in nortes and dry seasons, the diversity and evenness values are higher in nocturnal samplings compared with diurnal samplings. Regarding the rainy season, these values are smaller to those registered in the other seasons (Table 3).
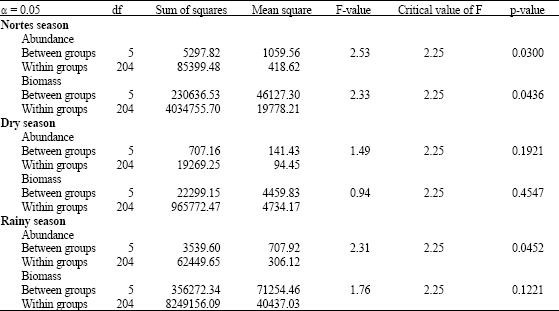
With the purpose of verifying if the obtained abundance and biomass records had significant differences regarding the seasons, a factor analysis of variance (α = 0.05) was carried out. This analysis shows that nortes and rainy seasons are the ones with significant differences in both parameters (Table 4) (ANOVA).
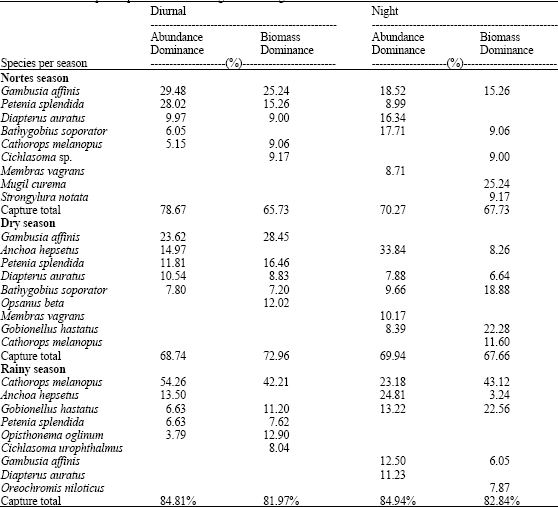
The estimation of abundance and biomass dominance according to McNaughton’s Index allowed the recognition that for diurnal samplings in nortes season there were nine species that represented between 65.73 and 78.67% of the records in both parameters of these 9 species. Gambusia affinis, Petenia splendida and Diapterus auratus, showed the greatest abundance records in diurnal hours in both parameters, while Gambusia affinis, Petenia splendida, Diapterus auratus and Bathygobius soporator provided with the greatest abundance records in nocturnal hours and Gambusia affinis, Mugil curema, Strongylura notata and Bathygobius soporator constituted the greatest records in biomass during nocturnal hours (Table 3).
During the dry season, nine species were dominant, providing 67.66-72.96% of the abundance records such as biomass. Regarding diurnal samplings, the species with the greatest abundance records were Gambusia affinis, Anchoa hepsetus, Petenia splendida and Diapterus auratus, being Gambusia affinis, Petenia splendida, Diapterus auratus and Opsanus beta the ones that provided the greatest biomass records in diurnal hours. The most abundant species for nocturnal samplings Anchoa hepsetus, Membras vagrans, Bathygobius soporator and Gobionellus hastatus, while regarding biomass, the species with the greatest records were Gobionellus hastatus, Bathygobius soporator, Cathorops melanopus and Anchoa hepsetus.
| Table 1: | Physicochemical seasonal parameters |
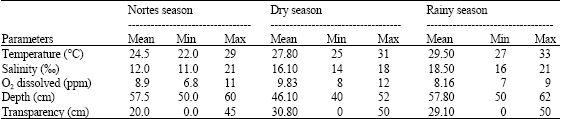 | |
| Table 2: | Seasonal Abundance (ABUN) (n/1200 m2) and Biomass (BIOM) (g/1200 m2) of the icthyofauna |
 | |
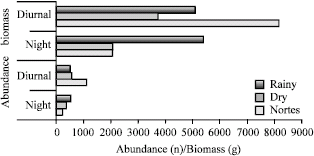 | |
| Fig. 2: | Abundance and biomass seasonal behavior |
In the rainy season, nine species were recognized as dominant; they represented 81.97-84.94% of the abundance and biomass records of this season. In diurnal captures, both Cathorops melanopus and Anchoa hepsetus were the most abundant species, whereas regarding the biomass, Cathorops melanopus, Gobionellus hastatus and Opisthonema oglinum offered the highest records. On the other hand, during nocturnal collections, Cathorops melanopus, Anchoa hepsetus Gobionellus, hastatus and Gambusia affinis were the most abundant and Cathorops melanopus, Gobionellus hastatus, Gambusia affinis and Oreochromis niloticus were the species that provided the greatest biomass (Table 5).
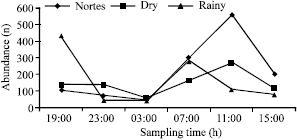 | |
| Fig. 3: | Abundance (n) seasonal behavior per sampling hour in Camaronera Lagoon |
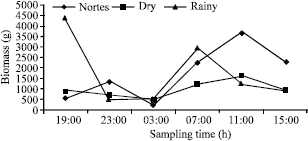 | |
| Fig. 4: | Biomass (g) seasonal behavior per sampling hour |
| Table 3: | Dominant species per season according to MacNaughton’s index |
 | |
| Table 4: | No. of species, diversity and eveness per season in Camaronera lagoon regarding abundance |
 | |
| Table 5: | Variance analysis of one factor (0.05) in order to prove significant differences in Abundance and Biomass per sampling hour and climatic season |
 | |
Feeding
A stomach contents analysis was carried out in 1333 organisms that represented 42% of the organisms captured in the three seasons. The main feeding types recognized by season and the schedule were as follows: nortes season, diurnal samplings registered fish of the Engraulidae and Gobiidae families, insects such as Hymenoptera, Diptera, Coleoptera and Homoptera, Ruppia maritima, anfipods and poliquets. For nocturnal captures, the registered feeding types were fish of the Engraulidae family, insects such as Hymenoptera, Hemipterous and Homoptera, detritus, Ruppia maritima, copepods and anfipods. During diurnal captures in the dry season, the registered feeding types were detritus, anfipods, poliquets, carideans and Ruppia maritima, while for nocturnal captures, the feeding types were detritus, Ruppia maritime, copepods, anfipods and algae. In the rainy season, the feeding types registered in diurnal captures were detritus, algae and remains of fish and in nocturnal captures, detritus and fish like Anchoa hepsetus were the most representative.
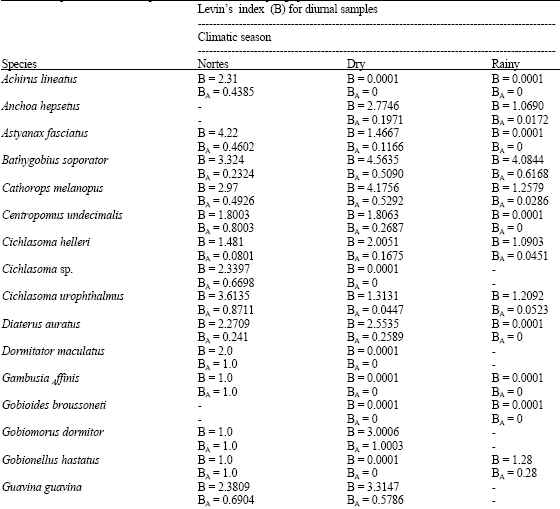
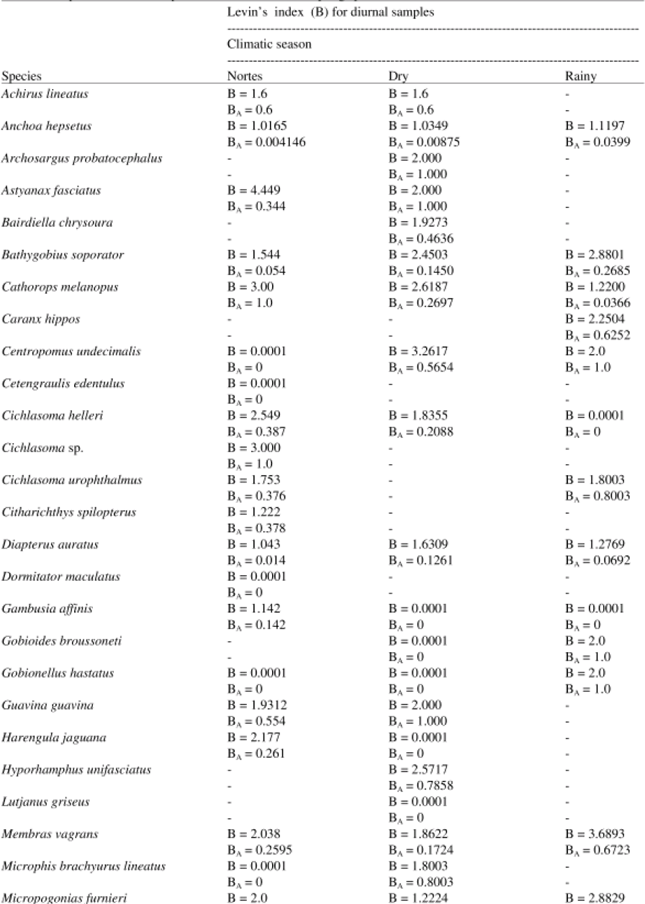
Levin’s standardized Index (BA) of niche amplitude for diurnal samplings shows that the feeding in dry and rainy seasons (Table 6) is dominated by few prey-types. It also shows the existence of certain specialization between the diet of Achirus lineatus, Astyanax fasciatus, Centropomus undecimalis, Cichlasoma sp., Diapterus auratus, Dormitator maculatus, Gambusia affinis, Gobioides broussoneti, Gobionellus hastatus, Membras vagrans, Mugil curema, Oreochromis niloticus and Strongylura notata. In nocturnal samplings, there are few prey-types present in the nortes season and species such as Centropomus undecimalis, Cetengraulis edentulus, Dormitator maculatus, Gobionellus hastatus, Microphis brachyurus lineatus, Mugil curema, Opisthonema oglinum and Oreochromis niloticus, exhibit certain specialization in their diets. In both cases, they complement their diet with other preys, depending on the available resources in each climatic season (Table 7).
| Table 6: | Amplitude index of trophic niche in diurnal samplings per climatic season |
 | |
 | |
| Table 7: | Amplitude index of trophic niche in nocturnal samplings per climatic season |
  | |
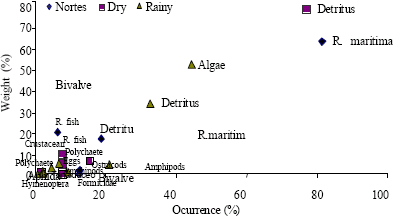 | |
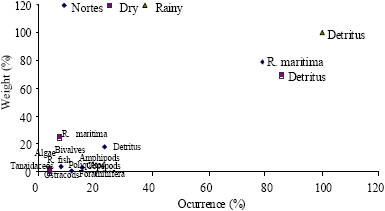
| Fig. 5: | Feeding types per season according to Costello’s graphic method for Cathorops melanopus |
 | |
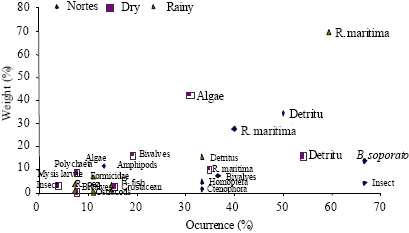
| Fig. 6: | Feeding types per season according to Costello’s graphic method for Diapterus auratus |
According to the obtained data of Costello’s graphic method, the species in all samples behaved as generalists, even though few feeding types dominate the diet of some species. Of the analyzed species, the ones that provided the greatest amount of feeding types were Cathorops melanopus (Fig. 5), Diapterus auratus (Fig. 6) and Petenia splendida (Fig. 7).
 | |
| Fig. 7: | Feeding types per season according to Costello’s graphic method for Petenia splendida |
DISCUSSION
The organization and structure of the community have been the more analyzed aspects currently, their study has focused in the analysis of its structure as well as in the species patterns that compose it (Schoener and Spiller, 1987). A factor that difficult this study is the form in which the analysis is considered, being of temporal or spatial character. In the present study, when approaching their analysis from the temporary point of view, the obtained results show us that the greatest records both in abundance and in biomass were obtained during diurnal hours (Fig. 3, 4), not based in the number of species (Table 4). This behavior coincides with what Adams (1976c), Bayer (1981), Gibson et al. (1993) and Methven et al. (2001), pointed out that for the reported lagoon and estuarine systems, the captures are bigger during nocturnal hours than during diurnal hours. If only the behavior of abundance and biomass by capture was analyzed, these are bigger during diurnal hours, compared with nocturnal hours, but if we integrate the specific composition and diversity, we can observe that nocturnal records are bigger.
An aspect that contributes to mask these results is the abundance records of the dominant species during sampling hours. This represent between 65 and 78% of the abundance and biomass records in the nortes season, 67 and 72% of the same parameters in the dry season and 81-84% of the abundance and biomass records in the rainy season (Table 3). Regarding this, Chávez et al. (1991) and Chavez and Franco (1993) reported most of these species for this zone as species with a great affinity due to freshwater conditions, becoming present in mesohaline conditions such as those registered along the study. The variance analysis that was carried out p = 0.030 in nortes season and p = 0.045 in rainy season allow to recognize a significant difference in the abundances for captures hour in both seasons.
This behavior could be related with climatic and meteorological events that dominate both seasons. During the nortes season, a bigger incursion of species is favored, registering 31, while in the rainy season, the high levels of fluvial discharge favor the presence of species with freshwater affinity, decreasing the specific richness to 22 species. During the dry season, the conditions of more thermal stability and reduction of the precipitation levels increase the specific richness, compared with the rainy season, since 28 species are present.
Another aspect to consider is the availability of the resources used by the species to develop their vital processes. In this lagoon, the presence of patches of Ruppia maritima offers them diverse components of physical and feeding character to cover their requirements. The physical components such as space and coverage favor most species with a protection area against depredators such as other piscivores fish. This behavior has been documented by Helfman (1978, 1993) and Gibson et al. (1993, 1996). Some other authors argue that high turbidity levels can reduce the rates of depredation of some species, but they can also favor others that do not depend on visual recognition (Benfiel and Minello, 1996). The implications of a decrease in the populations of major depredators in shallow waters imply the maintenance of abundances in other species, including those of small depredators that would have the potential of inflicting a substantial mortality in prey species (Sheaves, 2001). In this study, this is the case with Petenia splendida, which corresponded to a dominant species, but with organisms smaller than 10 cm of longitude pattern.
In some cases, the authors consider that the current conditions in this type of systems can reflect the adaptations of diverse species of fish, so that they may take advantage of the high rates of productivity that are characteristic in this type of environments. At the same time, they can adjust their biological cycle to the patterns of environmental variability (Flores et al., 1990; Arceo et al., 2004). Thus, the community that lives inside and near the inlets may take advantage of the stable environmental conditions, as well as of the organic matter that flows in the system, in order to cover its feeding requirements. These are represented in a higher proportion by consumer species of second and third orders (Flores et al., 1990). This behavior has been reported as well by other authors (Jenkins and Wheatley, 1998; Guidetti, 2000) for coastal environments and littorals as the one mentioned in this study. Here we recognize that the importance that each species has in the system is directly related with the functional role that it carries out in a certain moment of its live cycle.
As a complementary way, the levels of secondary production in littoral environments help to maintain a greater number of fish species than the ones that live in bordering zones (Platell and Potter, 2001; Travers and Potter, 2002). They affirm that a biggest organic contribution can favor macroinvertebrated growth that constitutes as preys of the fish species. The variety of feeding types registered in this study, shows slight differences between the analyzed seasons, where the anfipods, poliquets, carideans and copepods, Ruppia maritima and detritus are the main feeding items that maintain most of the registered species along the study. In this seasonal analysis, it is observed that some species show minimum variations in their feeding habits along the year, while others change their preferences in a drastic way, changing even the trophic level; this is the case with Petenia splendida.
The origins of these variations could be linked with several causes: competition, resource availability, trophic overlap, abundance and availability of disposable organisms for each climatic season and in this lagoon particularly, the contribution of organic matter from the fluvial systems that discharge in this lagoon system. The above-mentioned allows depredators species to enlarge their trophic spectrum because of the seasonal enrichment and the increase in productivity in the lagoon. This favors the growth of populations of prey species, thus enhancing the opportunity for the depredator species to enlarge their trophic spectrum, as Hori (2008) has mentioned it, in an analysis about rocky intertidal zones.
When comparing the obtained results of Levin’s Index in the different sampling seasons, it is observed that most of the species show low values of diet amplitude with values from 0-0.5; this indicates that their diet is composed of few feeding types. This is what happens with Astyanax fasciatus, Anchoa hepsetus, Bathygobius soporator, Cichlasoma helleri, Harengula jaguana, Membras vagrans among others; this index is greater in depredator species that show values that oscillate from 0.5-0.9, which confirms their opportunistic character when they consume a diet with a great amount of feeding items. In this group, we can mention the Centropomus undecimalis, Opsanus beta, Guavina guavina, Petenia splendida and Strongylura notata. Other species like Cathorops melanopus, Cichlasoma urophthalmus and Micropogonias furnieri show variations in this index, both during sampling hours and seasons. These variations suggest that these species consume any feeding type that is within their reach.
This behavior is reinforced when we analyze the feeding preferences with Costello’s graphic method, 1990. In this method we can see that, at least for the dominant species in all samplings, their diet is adjusted to the generalistic type, but with a marked preference for several feeding types such as detritus, grasses (Ruppia maritima) and algae (Fig. 5-7).
Both nictimeral and seasonal trophic behavior of the fish community in Camaronera lagoon, helped to distinguish the importance of feeding sources, such as detritus, crustaceans and Ruppia maritima in the system, to maintain levels of biological productivity. At the same time, the role played by the preys as important links that enrich the interactions in the current trophic nets was identified.
The seasonal analysis of the icthyofauna in Camaronera lagoon through diurnal and nocturnal samplings, helped to recognize that the better represented families were Cichlidae, Eleotridae and Gobiidae, accentuating Gambusia affinis, Petenia splendida, Cathorops melanopus, Diapterus auratus and Bathygobius soporator, as the most representative species regarding biomass and abundance.
The abundance behavior along the day shows significant variations in the nortes and rainy seasons, because of the climatic and meteorological conditions that prevail in both times of the year, favoring the incursion of species in the nortes season and limiting the entrance of marine species during the rainy season.
The trophic composition of the different species shows slight differences between the analyzed seasons, accentuating anfipods, poliquets, crustaceans (carideans and copepods), Ruppia maritima and detritus as the main feeding items that maintain most species registered along the study.
The estimate of the diet amplitude helped to recognize that some species, such as Astyanax fasciatus, Anchoa hepsetus, Bathygobius soporator, Cichlasoma helleri, Harengula jaguana and Membras vagrans, tend to show a specialist behavior in their feeding, while Centropomus undecimalis, Opsanus beta, Guavina guavina, Petenia splendida and Strongylura notata show a generalistic behavior. This behavior was confirmed when the information obtained in Costello’s Graphic Method was integrated; this information help to recognize that most of the species registered in the nortes, dry and rainy seasons show a generalistic strategy, even though few feeding types dominate the diet of some species.
The importance of the different elements in the maintenance of productive processes in the lagoon helps to recognize that a decrease in any of them could affect the community structure. This would affect trophic relationships and populational dynamics of the species, with a commercial and ecological impact for those who use the lagoon as a food source, for reproduction or as a protection zone in some period of their life cycle.
ACKNOWLEDGMENTS
The authors want to express their gratitude to the PAPCA Program 2006-2007 of FES-Iztacala, UNAM, for financing the project El sistema lagunar de Alvarado: Biodiversidad, hábitats y propuestas para su conservación (Alvarado Lagoon System: Biodiversity, Habitats and proposal for its conservation), which allowed the development and culmination of this study.
REFERENCES
- Adams, S.M., 1976. Feeding ecology of eelgrass fish communities. Trans. Am. Fish. Soc., 105: 514-519.
CrossRefDirect Link - Arceo, C.D., J. Franco, G.L. Waggy and R. Chavez, 2004. Trophic comparison of two species of needlefish (Belonidae) in the alvarado lagoonal system, Veracruz, Mexico. Gulf Caribbean Res., 16: 81-88.
Direct Link - Bayer, R.D., 1981. Shallow-water intertidal ichtyofauna of the yaquina estuary, oregon. Northwest Sci., 55: 182-193.
Direct Link - Benfiel, C.M. and T.J. Minello, 1996. Relative effects of turbidity and light intensity on reactive distance and feeding of an estuarine fish. Environ. Biol. Fish., 46: 211-216.
CrossRefDirect Link - Castro-Aguirre, J.L., 1999. Estuarine-Lagoonal Icthyofauna and Vicariance of Mexico. 1st Edn., Limusa, Mexico, DF., ISBN: 9681857747.
Direct Link - Chávez, L.R. and J.L. Franco, 1993. Response of a fish community before an environmental impact in Boca Camaronera, Alvarado, Veracruz. Hidrobiológica, 2: 25-33.
Direct Link - Flores-Verdugo, F., F. Gonzalez-Farias, O. Ramirez-Flores, F. Amezcua-Linares, A. Yanez-Arancibia, M. Alvarez-Rubio and J.W. Day, 1990. Mangrove ecology, aquatic primary productivity and fish community dynamics in the Teacapan-agua brava lagoon-Estuarine system (Mexican Pacific). Estuaries, 13: 219-230.
CrossRefDirect Link - Flores-Coto, C. and J. Álvarez, 1980. Preliminary study of distribution and abundance of ichthyoplankton in Términos Lagoon, Campeche. Ann. Centro Cienc. del Mar y Limnol., 7: 67-78.
Direct Link - Flores-Coto, C., V. Ducoing, F. Zavala-García, A. Velarde and S. Méndez., 1986. Effect of the tide in the passage of the larvae of some species of the family Clupeidae (Pises) in the Boca del Carmen, Términos Lagoon, Campeche, Mexico. Ann. Centro Cienc. del Mar y Limnol., 14: 53-68.
Direct Link - Franco, L.J. and R.L. Chávez, 1993. Summaries on the knowledge of the icthyofauna of Tamiahua Lagoon, Veracruz, Mexico. Hidrobiologica, 2: 53-63.
Direct Link - Gibson, R.N., A.D. Ansell and L. Robb, 1993. Seasonal and annual variations in abundance and species composition of fish and macrocrustacean communities on a Scottish sandy beach. Mar. Ecol. Prog. Ser., 98: 89-105.
Direct Link - Gibson, R.N., L. Robb, M.T. Burrows and A.D. Ansell, 1996. Tidal, diel and longer term changes in the distribution of fishes on a Scottish sandy beach. Mar. Ecol. Prog. Ser., 130: 1-17.
Direct Link - Guidetti, P., 2000. Differences among fish assemblages associated with nearshore Posidonia oceanica seagrass beds, rocky-algal reefs and unvegetated sand habitats in the Adriatic Sea. Estuar. Coast. Shelf Sci., 50: 515-529.
CrossRef - Helfman, G.S., 1978. Patterns of community structure in fishes: Summary and overview. Environ. Biol. Fish, 3: 129-148.
CrossRef - Hori, M., 2008. Between-habitat interactions in coastal ecosystems: Current knowledge and future challenges for understanding community dynamics. Plankton Benthos Res., 3: 53-63.
Direct Link - Jenkins, G.P. and M.J. Wheatley, 1998. The influence of habitat structure on nearshore fish assemblages in a southern Australian embayment: Comparison of shallow seagrass, reef-algal and unvegetated sand habitats, with emphasis on their importante to recruitment. J. Exp. Mar. Biol. Ecol., 221: 147-172.
CrossRef - Marshall, S. and M. Elliott, 1997. A comparison of univariate and multivariate numerical and graphical techniques for determining inter- and intraspecific feeding relationships in estuarine fish. J. Fish Biol., 51: 526-545.
CrossRefDirect Link - Methven, D.A., R.L. Haedrich and G.A. Rose, 2001. The fish assemblage of a Newfoundland Estuary: Diel, Monthly and Annual Variation. Estuar. Coast. Shelf Sci., 52: 669-687.
CrossRefDirect Link - Platell, M.E. and I.C. Potter, 2001. Partitioning of food resources amongst eighteen abundant benthic carnivorous fish species in marine waters on the lower west coast of Australia. J. Exp. Mar. Biol. Ecol., 261: 31-54.
CrossRefDirect Link - Post, J.C. and G.C. Lundin, 1996. Guidelines for integrated coastal zone management. World Bank Publications, Series: Environmentally Sustainable Development Studies and Monographs, Washington. http://www-wds.worldbank.org/servlet/WDSContentServer/WDSP/IB/1996/08/01/000009265_3961219091924/Rendered/PDF/multi_page.pdf.
- Roman, H.U., J.Z. Valdez and F.G. Zavala, 2006. Composition and abundance of ichthyoplankton during the low-water mark season in the Tampamachoco Lagoon, Veracruz, Mexico. Revista Científica UDO Agrícola, 6: 138-149.
Direct Link - Schoener, T.W. and D.A. Spiller, 1987. High population persistence in a system with high turnover. Nature, 330: 474-477.
CrossRefDirect Link - Sheaves, M., 2001. Are there really few piscivorous fishes in shallow estuarine habitats? Mar. Ecol. Prog. Ser., 222: 279-290.
Direct Link - Travers, M.J. and I.C. Potter, 2002. Factors influencing the characteristics of fish assemblages in a large subtropical marine embayment. J. Fish Biol., 61: 764-784.
CrossRefDirect Link - Twilley, R.R., R.H. Chen and T. Hargis, 1992. Carbon sinks in mangroves and their implications to carbon budget of tropical coastal ecosystems. Water Air Soil Pollut., 64: 265-288.
CrossRefDirect Link - Yañez-Arancibia, A. and R. Nugent, 1977. The ecological role of fish in estuaries and coastal lagoons. Ann. Inst. Cienc. Mar y Limnol., 4: 107-117.
Direct Link