
A. Helmi
Department of Plant Protection, Faculty of Agriculture, Ain Shams University, PB. 68 Hadayek Shobra El Kheima, Cairo, Egypt
International Journal of Zoological Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 77-84







ABSTRACT
Whitefly, Bemisia tabaci (Genn.) is an important sucking plant sap pest of field, horticultural and ornamental plants causing feeding injuries besides spreading plant diseases by acting as a vector of Gemini-viruses. The polyphagous nature of the pest makes it as a highly complex species. The influence of six host plants belonging to three different plant families utilized by the species on the population differences at molecular level was attempted using Random Amplified Polymorphic DNA (RAPD) markers. Fifteen RAPD primers were screened seven of them were produced 218 DNA fragments, 209 of them were polymorphic while the other nine bands could be considered as common for B. tabaci. Total number of bands obtained from each primer ranged from 23-44 with an average of 36.33 bands per primer. RAPD-PCR analysis led to identification of 42 polymorphic markers holding specificity for these hosts' populations. Phylogenetic relationships among the studied populations using this technique clearly separated these six populations into two main clusters with similarity matrix percentage of 88 and 64%. These results indicated that B. tabaci may have different genotypes on adaptations to certain host plant species in Egypt.
PDF Abstract XML References Citation
Received: February 17, 2010;
Accepted: May 26, 2010;
Published: August 21, 2010
How to cite this article
A. Helmi, 2011. Host-Associated Population Variations of Bemisia tabaci (Genn.) (Hemiptera:Sternorrhyncha: Aleyrodidae) Characterized with Random DNA Markers. International Journal of Zoological Research, 7: 77-84.
DOI: 10.3923/ijzr.2011.77.84
URL: https://scialert.net/abstract/?doi=ijzr.2011.77.84
DOI: 10.3923/ijzr.2011.77.84
URL: https://scialert.net/abstract/?doi=ijzr.2011.77.84
INTRODUCTION
Bemisia tabaci (Gennadius) is a cosmopolitan agricultural pest and the so called B biotype is ranked as one of the world’s worst invasive organisms. It is broadly polyphagous, feeding on an estimated 900 host plants (Jones 2003; Ma et al., 2007). This species is considered not only cause aesthetic damage to plants, but also as an efficient vector of many plant viruses (Jones, 2003; Muniz et al., 2004; Brown, 2007). An important aspect of the biology of B. tabaci is the high genetic variability that exists among its populations (Iida et al., 2009). This variability is revealed by the existence of populations that differ in their ability to feed or reproduce on particular hosts and in their virus transmission characteristics (Simmons et al., 2009). This species was raised to the rank of a new species (Bellows et al., 1994) which named Bemisia argentifolii Bellows and Perring, that produces unique symptoms of squash silver leaf. Recent research has tended to prove that B. tabaci corresponds to a species or biotypes complex in phylogenetic evolution. Its distinctive characteristics were evaluated by using different methods commonly used in taxonomy (Perring, 1996; Amin et al., 2009; Qiu et al., 2009). Previous studies revealed that B. tabaci is genetically complex and composed of numerous well defined genetic groups (De Barro et al., 2005; Maruthi et al., 2001; De Barro et al., 2005; Boykin et al., 2007). These biotypes usually are recognized by the presence of specific phytotoxic reactions (Muniz et al., 2002) and characterized by esterase markers (Burban et al., 1992; Lisha et al., 2003) as well as several DNA fingerprinting techniques such as microsatellites which identify genetic variability based upon limited number of genetic regions, RAPD profiles generated with a number of RAPD primers are considered advantageous as they help to generate comparative profiles based on randomly chosen genetic regions all over the genetic element, DNA (De Barro and Driver, 1997; Guirao et al., 1997; Cervera et al., 2000; Lima et al., 2000; Martinez et al., 2000; Moya et al., 2001; Cenis, 2003; Gobbi et al., 2003; Sartor et al., 2008; Sharma et al., 2008; Perumal et al., 2009; Qiu et al., 2009). Although much of the information is available on the biotype prevalence based on the locations and regions, molecular information on the genetic diversity of populations based on host plants is scarce.
This study is an attempt to address our hypothesis that the population of the whitefly varies widely depending upon the host plants being utilized by poly-crop systems.
MATERIALS AND METHODS
Insect population samples: Whitefly pre-pupal instar were collected from the middle leaves of six host plants belonging to three families; Cucurbitaceae (Squash, Cucurbita pepo ovifera and Watermelon, Citrullus vulgaris); Solanaceae (Egg-plant, Solanum melongena and Tomato, Solanum lycopersicum) and Malvaceae (Cotton, Gossypium barbadensa and Okra Hibiscus esculantus) from Shalakan farm at Qalyubiya Governorate during 2009 and allowed for adult emergence using insect emergence cages at the laboratory. All individual populations were maintained continuously for three successive generations on the respective host plants which were grown in pots at rearing cages. The purity of individual populations was ensured by raising new population from the parental adults of an earlier generation after these were shifted from the old screen cages to new cages. The emerging adult females from the third generation were separated and preserved immediately at -20°C in Eppendorf tubes until DNA extraction.
Randomly Amplified Polymorphic DNA of the Polymerase Chain Reaction (RAPD-PCR)
DNA extraction: DNA was extracted from adult females of each population. Two adult females of each population were crushed in 1.5 mL Eppendorf tubes to extract the genomic DNA filled with 200 μL of extraction buffer (200 mM Tris-HCl pH 8.5, 250 mM NaCl, 25 mM EDTA, 0.5% SDS). Then, 100 μL of 3 M sodium acetate pH 5.2 was added and tubes were placed at -20°C for 10 min. Tubes were then centrifuged for 5 min at 10.000 rpm and the supernatant transferred to new Eppendorf tubes. An equal volume of isopropanol was added and the precipitated DNA was pelleted by centrifugation at 13.000 rpm for 20 min. After washing with 70% ethanol, the pellet was dried and resuspended in 100 of TE buffer (10 mM Tris-HCl, 1 mM EDTA pH 8.0).
Polymerase Chain Reaction (PCR): Comparative RAPD-PCR analysis of whitefly from different hosts was performed using whitefly DNA as template and fifteen RAPD primers (Operon biotechnologies, Inc, Alameda, California) were screened RAPD-PCR amplifications were performed in a total volume of 25 μL. Each reaction contained 2.5 μL of 10X buffer, 2.5 μL dNTPs, 2.5 μL MgCl2, 3 μL primer, 1 μL taq polymerase, 2 μL genomic DNA with the volume adjusted to 25 μL with distilled water. Amplifications were carried out in Techne system, England using the following program, one cycle at 95°C for 5 min, followed by 40 cycles each at 94°C for 30 sec, 37°C for 1 min, 72°C for 2 min and final extension at 72°C for 12 min. PCR Products were separated in 1.2% agarose gels at 100 volts. Gels were stained with ethidium bromide and products were visualized by UV light and photographed.
Analysis of RAPD-PCR profiles for genetic relatedness: All gels resulted from RAPD-DNA were scanned using Bio-Rad Gel Doc. 2000 Quantity one software version 4.0.3. To calculate the pair-wise differences matrix among the six B. tabaci populations, all the individual bands of different host specific whitefly samples in RAPD banding profile were scored as presence (1) or absence (0) and a genetic relatedness dendrogram was constructed using the Statistical Package for Social Science (SPSS ver. 10) soft ware program.
RESULTS AND DISCUSSION
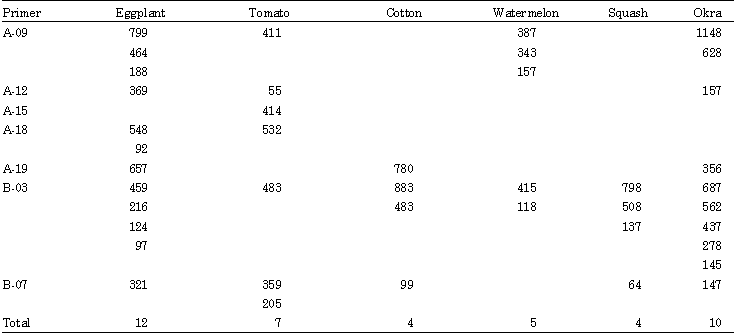
RAPD-PCR analysis: To select genetically heritable markers, each individual whitefly population was sampled from the third grown generation for all the six host specific populations to identify only genetically inheritable bands/markers in the RAPD profiles. Fifteen 10-mer random primers were screened against each individual whitefly population out of them seven primers produced fragment bands. These primers are: A-09, A-12, A-15, A-18, A-19, B-03 and B-07 with the following sequences (5’GGGTAACGCC’3), (5’TCGGCGATAG’3), (5’TTCCGAACCC’3), (5’AGGTGACCGT’3), (5’CAAACGTCGG’3), (5’CATCCCCCTG’3) and (5’GGTGACGCAG’3) for the seven primers, respectively. Those amplified distinct polymorphic bands were used for studying genetic variability among different host specific populations. These seven primers amplified a total number of 218 DNA fragments 209 of these fragments were polymorphic (Fig. 1A-G). While the other nine fragments were detected as common among the six tested populations. The total number of bands generated from each primer ranged from 23 (OPA15) to 44 (OPA18) with an average on 36.33 bands per primer. The size of bands was ranged from 27 to 3277 bp. The total numbers of bands amplified by these seven primers for each population were 36, 30, 47, 36, 33 and 36 bands for B. tabaci populations on eggplant, tomato, cotton, watermelon, squash and okra, respectively. The comparative analysis of these RAPD profiles from different whitefly types resulted in identification of a number of 42 polymorphic markers holding specificity for the studies hosts populations. These genetic differences were visualized as host specific polymorphic bands and amplified from particular host specific whitefly represented host specific molecular markers for respective whitefly genotype. The highest number of these host specific markers (17 bands) was produced by OPB-03 primer, while the smallest number of these markers was one band produced by OPA-15 primer for tomato population. The total numbers of host specific markers per each whitefly genotype were 12, 7, 4, 5, 4 and 10 for populations from eggplant, tomato, cotton, watermelon, squash and okra, respectively (Table 1). The number of common bands of each two hosts-associated populations of B. tabaci amplified by seven random primers was ranged between 24 bands for populations on watermelon and cotton to two bands for populations on eggplant and okra (Table 2).
Random amplified polymorphic DNA polymerase chain reaction (RADP-PCR) is a relatively simple, inexpensive and rapid technique, revealing polymorphisms which are useful as genetic and taxonomic markers (Welsh and McClelland, 1990). RAPD has been applied to study of insects (Haymer, 1994) and to differentiate whiteflies, including the identification of different biotypes of B. tabaci (Guirao et al., 1997; De Barro and Driver, 1997; Cervera et al., 2000; Moya et al., 2001). Sharma et al. (2008) detected genetic variability due to host plants in B. tabaci populations that collected from six different host plants using RAPD-PCR. They mentioned that the whitefly types holding specificity for some host plants were studied and cited that the sequence information on these RAPD-DNA markers can be used to design more efficient, specific molecular markers with specificity to different host plants. Perumal et al. (2009) found differences among B. tabaci populations collected from four different host plants at the same ecosystem were found at the same point of time but collected from various locations of Tamil Nadu using RAPD-PCR.
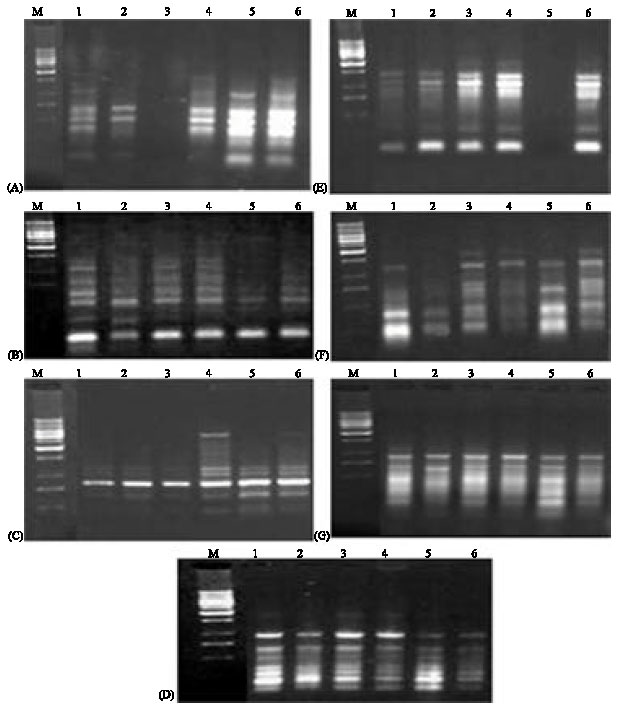 | |
| Fig. 1: | RAPD-PCR banding patterns of six B. tabaci host-associated populations by using seven random primers. (A) Primer A9; (B) Primer A12; (C) Primer A15; (D) Primer A18; (E) Primer A19; (F) Primer B3; (G) Primer B7, M, DNA marker; bp, base pair 1: Eggplant; 2: Lantana; 3: Cotton; 4: Watermelon; 5: Squash; 6: Okra |
They reported that there were at least two different biotypes of this insect pest based on these four host plants. While, Frohlich et al. (1999) stated that B. tabaci populations and their respective host plants occur in the same geographical region at the same point of time the identified genetic diversity appears to be a rare example of the evolutionary transition leading to sympatric speciation.
| Table 1: | RAPD molecular markers (bp) associated with host specificity in B. tabaci |
 | |
| Table 2: | The similarity matrix among different six host-associated populations of B. tabaci based on RAPD-PCR |
 | |
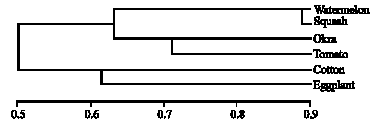 | |
| Fig. 2: | Phylogenetic relationships among different six host-associated populations of B. tabaci based on seven primers of RAPD-PCR |
Genetic relatedness amongst the studied B. tabaci populations on six host plants: Genetic similarities and genetic relatedness amongst the six host-associated populations of B. tabaci were based on data obtained of seven random primers as molecular markers of RAPD-PCR. These data were subjected to using SPSS computer program to support the existence of high level of genetic relatedness amongst the investigated whitefly types. This genetic relatedness was not clear amongst the plant family populations (Fig. 2). While, relatedness dendrogram was indicated two main clusters with similarity matrix percentage of 54%. The first cluster was divided into two sub-clusters; the first one included squash and watermelon with similarity matrix percentage of 88%; the second sub-cluster included okra and tomato B. tabaci populations with similarity matrix percentage of 74%. While the second cluster was included cotton and eggplant B. tabaci populations with similarity matrix percentage of 64%. From this similarity dendrogram it could be concluded that the relationship among plant families and B. tabaci populations was not clear except in plants belonging to family Cucurbitaceae that showed a high similarity matrix. While this similarity matrix was not so in the plants belonging to the other two families requires further biological studies. Also there was host specificity amongst under study B. tabaci populations based on molecular markers amplified by these seven random primers.
In this respect, Sharma et al. (2008) found three groups of host specific populations of B. tabaci on six host plants grown in the same locality and in the same time based on RAPD-PCR using nine random primers. While many studies of geographic distributions of B. tabaci genetic groups around the globe. This level of genetic diversity had identified at least 33 different biotypes (Perring, 2001; Simon et al., 2003; Zang et al., 2006). The global phylogenies of geographic groups of B. tabaci suggested that B. tabaci was a cryptic species complex. The overall analysis suggested that there were at least 24 species making up the complex (Boykin et al., 2007; Dinsdale et al., 2010).
This study confirms that there exists population isolations based on host plants among the whitefly, B. tabaci population. Therefore, this point needs further molecular analysis to understand the physiological and evolutionary relationships, which may through some lights for taxonomical perspectives and pest management decisions. Also these results need further biological studies such as the ability to transmit plant viruses (Brown, 2007), the rate of female fecundity (Iida et al., 2009; Xu et al., 2010) and the ability to accelerate pesticides resistance (Horowitz et al., 2005).
CONCLUSION
From the above mentioned results it could be concluded that genetic variability assessment in B. tabaci populations originating from different host plants at Qalyubiya Governorate, Egypt indicates that the population was diversified based on the host species. Clustering pattern observed in the dendrogram showed that at least two distinct genotypes exist among the populations collected within the narrow region of Egypt. These differences may be influencing the virus vectoring capabilities of the whitefly population and also their susceptibility to insecticides, which needs further studies.
ACKNOWLEDGMENTS
Author would like to express his deep thanks and gratitude to Dr. Abeer, A. Abdel-Bary and Dr. Ahmed Abd El Khalek staff members in Genetic Department, Faculty of Agriculture, Ain Shams University, for their helping, providing facilities and advices me with their experts throughout the molecular analysis.
REFERENCES
- Bellows Jr., T.S., T.M. Perring, R.J. Gill and D.H. Headrick, 1994. Description of a species of Bemisia (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am., 87: 195-206.
CrossRefDirect Link - Boykin, L.M., R.G. Shatters, R.C. Rosell, C.L. McKenzie, R.A. Bagnall, P.J. de Barro and D.R. Frohlich, 2007. Global relationships of Bemisia tabaci (Hemiptera: Aleyrodidae) revealed using Bayesian analysis of mitochondrial CO1 DNA sequence. Mol. Phylogenet. Evolut., 44: 1306-1319.
CrossRef - Burban, C., L.D.C. Fishpool, C. Fauquet, D. Fargette and J.C. Thouvenel, 1992. Host associated biotypes within West African populations of the whitefly Bemisia tabaci (Genn.) (Homoptera, Aleyrodidae). J. Applied Entomol., 113: 416-423.
Direct Link - De Barro, P.J. and F. Driver, 1997. Use of RAPD PCR to distinguish the B biotype from other biotypes of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Aust. J. Entomol., 36: 149-152.
CrossRefDirect Link - Dinsdale, A., L. Cook, C. Riginos, Y.M. Buckley and P.J. de Barro, 2010. Refined global analysis of Bemisia tabaci (Gennadius) (Hemiptera: Sternorrhyncha: Aleyrodoidea) mitochondrial CO1 to identify species level genetic boundaries. Ann. Entomol. Soc. Am., 103: 196-208.
CrossRefDirect Link - Frohlich, D.R., I. Torres-Jerez, I.D. Bedford, P.G. Markham and J.K. Brown, 1999. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol., 8: 1683-1691.
CrossRefPubMedDirect Link - Guirao, P., F. Beitia and J.L. Cenis 1997. Biotype determination of Spanish populations of Bemisia tabaci (Hemiptera: Aleyrodidae). Bull. Entomol. Res., 87: 587-593.
CrossRef - Haymer, D., 1994. Arbitrary (RAPD) primer sequences used in insect studies. Insect Mol. Bio., 3: 191-194.
CrossRefDirect Link - Horowitz, A.R., S. Kontsedalov, V. Khasdan and I. Ishaaya, 2005. Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance. Arch. Insect Biochem. Physiol., 58: 216-225.
CrossRefPubMedDirect Link - Iida, H., T. Kitamura and K. Honda, 2009. Comparison of egg-hatching rate, survival rate and development time of the immature stage between B- and Q-biotypes of Bemisia tabaci (Gennadius)(Homoptera: Aleyrodidae) on various agricultural crops. Applied Entomol. Zool., 44: 267-273.
Direct Link - Xu, J., P.J. de Barro and S.S. Liu, 2010. Reproductive incompatibility among genetic groups of Bemisia tabaci supports the proposition that the whitefly is a cryptic species complex. Bull. Entomol. Res., 100: 359-366.
PubMedDirect Link - Jones, D.R., 2003. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol., 109: 195-219.
CrossRefDirect Link - Lima, L.H.C., D. Navia, P.W. Inglis and M.R.V. De Oliveira, 2000. Survey of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biotypes in Brazil using RAPD markers. Genet. Mol. Biol., 23: 781-785.
Direct Link - Lisha, V.S., B. Antony, M.S. Palaniswami and T.J. Henneberry, 2003. Bemisia tabaci (Homoptera: Aleyrodidae) biotypes in India. J. Econ. Entomol., 96: 322-327.
CrossRefDirect Link - Ma, D., K. Gorman, G. Devine, W. Luo and I. Denholm, 2007. The biotype and insecticide-resistance status of whiteflies, Bemisia tabaci (Hemiptera: Aleyrodidae), invading cropping systems in Xinjiang Uygur Autonomous Region, Northwestern China. Crop Prot., 26: 612-617.
CrossRef - Martinez, S.S., A.O.R. Carvalho, L.G. Vieira, L.M. Nunes and A. Bianchini, 2000. Identification, geographical distribution and host plants of Bemisia tabaci (Genn.) biotypes (Homoptera Aleyrodidae) in the state of Parana, Brazil. Ann. Soc. Entomol. Bras., 29: 597-603.
CrossRefDirect Link - Maruthi, M.N., J. Colvin and S. Seal, 2001. Mating compatibility, life-history traits and RAPD PCR variation in Bemisia tabaci associated with the cassava mosaic disease pandemic in East Africa. Entomol. Exp. Appli., 99: 13-23.
CrossRefDirect Link - Moya, A., P. Guirao, D. Cifuentis, F. Beitia and J.L. Cenis, 2001. Genetic diversity of Iberian populations of Bemisia tabaci (Hemiptera: Aleyrodidae) based on random amplified polymorphic DNA-polymerase chain reaction. Mol. Ecol., 10: 891-897.
CrossRefDirect Link - Muniz, M., G. Nombela and L. Barrios, 2002. Within-plant distribution and infestation pattern of the B- and Q-biotypes of the whitefly, Bemisia tabaci, on tomato and pepper. Entomol. Exp. Applicata, 104: 369-373.
CrossRef - Muniz, M., Y.X. Jiang, G. Nombela, C. de Blas and I. Bedford, 2004. Short communication. Effect of Bemisia tabaci biotype in the transmission of tomato yellow leaf curl sardinia virus (TYLCSV-ES) between tomato and common weeds. Spanish J. Agric. Res., 2: 115-119.
Direct Link - Perring, T.M., 2001. The Bemisia tabaci species complex. Crop Prot., 20: 725-737.
CrossRefDirect Link - Qiu, B., Y. Chen, L.L.W. Peng, X. Li and M.Z. Ahmed et al., 2009. Identification of three major Bemisia tabaci biotypes in China based on morphological and DNA polymorphisms. Prog. Natural Sci., 19: 713-718.
CrossRef - Sartor, C., S. Demichelis, J.L. Cenis, A.K. Coulibaly and D. Bosco, 2008. Genetic variability of Bemisia tabaci in the Mediterranean and Sahel regions. Bull. Insectol., 61: 161-162.
Direct Link - Simmons, A.M., K.S. Ling, H.F. Harrison and D.M. Jackson, 2009. Weet potato leaf curl virus: Efficiency of acquisition, retention and transmission by Bemisia tabaci (Hemiptera: Aleyrodidae). Crop Protect., 28: 1007-1011.
CrossRef - Simon, B., J.L. Cenis, S. Demichelis, C. Rapisarda, P. Caciagli and D. Bosco, 2003. Survey of Bemisia tabaci (Hemiptera: Aleyrodidae) biotypes in Italy with the description of a new biotype (T) from Euphorbia characias. Entomol. Res., 93: 259-264.
CrossRef - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Zang, L.S., T. Jiang, J. Xu, S.S. Liu and Y.J. Zhang, 2006. SCAR molecular markers of the B biotype and two non-B populations of the whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Chinese J. Agric. Biotech., 3: 189-194.
CrossRef