
R. Offei- Oknye
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, Normal, Alabama, United States of America
J.L. Patterson
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, Normal, Alabama, United States of America
L.T. Walker
Food Chemistry Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, Normal, Alabama, United States of America
J. Boateng
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, Normal, Alabama, United States of America
M. Verghese
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama Agricultural and Mechanical University, Normal, Alabama, United States of America
International Journal of Cancer Research
Year: 2015 | Volume: 11 | Issue: 2 | Page No.: 52-66







ABSTRACT
Phytochemicals are known to have antioxidative, antibacterial, antifungal, anti-inflammatory and anticancer properties. In this study, HepG2 hepatoma cells were incubated after 12 and 24 h with 80% methanol extracts of processed ginger: Oven dried, sun dried, freeze dried and fresh ginger (control). The objectives of this study were to investigate the chemopreventive potential of ginger extracts in hepatoma cell lines (induction of cytotoxicity and apoptosis) and to determine the effects of ginger extracts on selected detoxification and anti-oxidative enzymes: Glutathione-S-transferase, Glutathione reductase, Glutathione peroxidase, Superoxide dismutase, Catalase activity and Glutathione in cell lines (hepatoma). Processed ginger extracts induced higher cytotoxicity (%) in HepG2 (SD 60 μg mL-1 [92.89]). Induction of cytotoxicity (%) was higher after 24 h of incubation compared to 12 h. Histone related DNA fragmentation (enrichment factor), an indicator of apoptosis was higher after 12 h compared 24 h of incubation except with cell incubated with FD (240 μg mL-1). Detoxification and antioxidative enzymes studied generally showed a higher induction after 24 h compared to 12 h of incubation in SOD, GR, CAT, GPx and GSH (antioxidant substrate). These findings indicate that ginger may exerts its chemopreventive effects by its cytotoxic activity and the induction of detoxification and antioxidative enzymes.
PDF Abstract XML References Citation
Received: November 30, 2014;
Accepted: March 10, 2015;
Published: June 06, 2015
How to cite this article
R. Offei- Oknye, J.L. Patterson, L.T. Walker, J. Boateng and M. Verghese, 2015. Chemopreventive Potential of Ginger on Hep2G Cells. International Journal of Cancer Research, 11: 52-66.
DOI: 10.3923/ijcr.2015.52.66
URL: https://scialert.net/abstract/?doi=ijcr.2015.52.66
DOI: 10.3923/ijcr.2015.52.66
URL: https://scialert.net/abstract/?doi=ijcr.2015.52.66
INTRODUCTION
Pathogenesis of many chronic disease such as cardiovascular and cancer has been attributed to metabolic generated free radical species (Harman, 1992; Das, 2002; Winyard et al., 2005). However, phytochemicals found in fruits and vegetables are known to impact health benefits (Aggarwal and Shishodia, 2006).
Hepatocellular carcinoma is ranked third in cancer motality and morbidity world (Chen et al., 2006) but relatively rare in the western countries including the US (Altekruse et al., 2009). The increasing cost of cancer treatment (Schrag, 2004) partly as a result of resistance to conventional treatment methods, such as chemo and radiotherapy (Boehm et al., 1997), has heightened the interest in the need for preventive method (Liu, 2003). Dietary source of antioxidants may prevent oxidative-related damage such as cancer from reactive oxygen species (Halliwell and Gutteridge, 1995).
The damage of biological molecules by Reactive Oxygen Species (ROS) has been linked to weakened immune system, degenerative diseases (Coyle and Puttfarcken, 1993) and cancer (Ames, 1989; Loft and Poulsen, 1996; Valko et al., 2004). However, antioxidants in diet may reduce the damaging effects of free radicals (Machlin and Bendich, 1987). Dietary source of phytochemicals or antioxidants are also known to activate endogenous antioxidative enzymes (Surh et al., 2008).
Surh (2003) reviewed the chemopreventive properties of phytochemicals. The contribution of phenols as chemopreventive agent is as a result of its antioxidative and anti-inflammatory properties (Surh et al., 2001). The action of phytochemicals as chemopreventive agents can be grouped into two: cancer-blocking and cancer-suppressing agents (Russo, 2007). Detoxification, scavenging Reactive Oxygen Species (ROS) and apoptosis are important mechanisms in chemoprevention (Hatono et al., 1996; Yu et al., 1998; Mates, 2000).
Research by Prakash and Srinivasan (2010) indicated increase in activity of antioxidant enzymes, such as superoxide dismutase (SOD), Catalase (CAT), Glutathione Reductase (GR) and Glutathione-S-Transferase (GST) after Wistar rats were fed with varieties of spices. The first line of defense against ROS are the SODs, they catalyze the dismutation of superoxide anions (Zelko et al., 2002). In tumors, there is an increased production of superoxide radicals (Huang et al., 2000) and a decrease in enzyme activities such as SOD and CAT (Oberley and Oberley, 1986). However, tumors have inconsistent levels of GPx activity compared to normal tissues (Oberley and Buettner, 1979). The functions of GSH includes catalysis, metabolism and transport (Meister, 1988), it acts as an important electron donor to GPx in the reduction of hydrogen peroxide to water (Diplock, 1994; Halliwell and Gutteridge, 1995).
Ginger (Zingiber officinale roscoe), (Zingiberaceae) is a herbaceous perennial plant (Salmon et al., 2012) which has been used as and for the treatment of human ailments for many years (Ali et al., 2008). Clinical studies have reported ginger to be effective in the treatment of various conditions. Specifically, ginger has been shown to possess anti-inflammatory, antioxidant, lipid-lowering, anti-platelet and hypotensive effects (Nicoll and Henein, 2009). Ginger is extensively cultivated in tropical and subtropical countries (Berza et al., 2012) and has been used as food flavoring agent for many years (John and Amanda, 2000). The chemoprevention of specific compounds in ginger such as [6]-gingerol and [6]-paradol have been studied (Lee and Surh, 1998). However, limited studies exit on the effect of different processing methods of ginger: Oven Dried (OD), Sun Dried (SD), Freeze Dried (FD) and Fresh Ginger (FG) on antioxidant and detoxification enzymes in HepG2 cell.
The objectives of this study were to determine the chemopreventive potential of fresh and processed ginger extracts (oven-driedODin relation to its cytotoxicity, antioxidative enzyme activity (SOD, CAT, GPx, GR and GSH) and detoxification enzyme (GST) in HepG2 cells.
MATERIALS AND METHODS
Cell culture: A HepG2 human liver cell (ATCC HTB-37) was obtained from the American Type Culture Collection (ATCC, Manassas, VA). Cells were maintained in Dulbecco’s modified Eagles Media (DMEM) with 10% fetal bovine serum. For assay, 5×104 HepG2 cells were seeded in a 24 well culture plate and incubated at 37°C and 7% CO2 until development of a monolayer. After the formation of a monolayer, 400 μL of fresh media (DMEM serum free) was added to the wells. Different concentrations of ginger extracts (oven dried, powdered, fresh and freeze dried ginger) were made up to 100 μL with saline, added and incubated for 12 and 24 h.
Ginger extracts: Ethanol extracts of Fresh Ginger (FG), oven Dried Ginger (OD), sun dried ginger (SD) and freeze dried ginger (FD) were prepared at concentrations of 30-240 μg mL-1 from mature ginger rhizomes.
Determination of lactate dehydrogenase (LDH): The cytotoxic effects of ginger extracts (30, 60, 120 and 240 μg mL-1) was measured by the release of lactate dehydrogenase (LDH) from the HepG2 cells into the culture supernatant. The cells were incubated with the extracts for 12 and 24 h. For positive control, 10% triton-X solution was used and the supernatant was collected. LDH was quantified using a colorimetric cytotoxicity detection kit (LDH) (Roche Diagnostics, Indianapolis, IN) according to the manufacturer’s instructions. The absorbance was read at 490/600 nm using a plate reader (Synergy HT, Bio Tek Instruments, Vermont, USA).
Cell lysate preparation: HepG2 cells were treated with ginger extracts at concentrations of 30, 60, 120 and 240 μg mL-1 and incubated for 12 and 24 h. The cells were extracted with PBS (pH 7) and centrifuged at 10,000 g for a period of 5 min. Cell lysis buffer was added to the pellet and cells were lysed by repeated cycles of freezing and thawing in liquid nitrogen. Supernatant was obtained after the cell suspension was centrifuged at 15,000 g for 15 min.
Glutathione-S-transferase activity in cell lysates: Glutathione-S-transferase activity in cell lysates was determined using the manufacturer’s specifications (Cayman Chemical Company, Ann Arbor, MI). Kinetic absorbance was read at 340 nm at 5 min intervals using a plate reader (Synergy HT, Bio Tek Instruments, Vermont, USA).
Determination of catalase activity: Catalase activity was determined using a microplate reader (BioTek Synergy HT) at 240 nm as described by Aebi (1984). Cell lysate (20 μL) was added to phosphate buffer (250 mM KH2PO4, 10 mM EDTA, 0.1% BSA pH 7.5). Briefly, 30 μL of methanol (5.9 M) and 20 μL of H2O2 (0.12%) were added and the mixture incubated for 20 min at room temperature. The reaction was stopped by the addition of 30 μL of KOH (7.8M), 30 μL purpald and potassium periodate (10 μL).
Determination of superoxide dismutase (SOD) activity: Superoxide dismutase activity was determined in cell lysates using the manufacturer’s (Cayman Chemical Company, Ann Arbor, MI) specifications. Absorbance was read at 440 nm using a plate reader (Synergy HT, Bio Tek Instruments, Vermont, USA).
Determination glutathione levels: Glutathione level was determined using manufacturer’s specifications (Cayman Chemical Company, Ann Arbor, MI). Kinetic absorbance was read at 405 nm at 5 min intervals for 30 min using a plate reader (Synergy HT, Bio Tek Instruments, USA).
Glutathione peroxidase and glutathione reductase activity: Glutathione peroxidase and glutathione reductase activities were estimated using manufacturer’s (Cayman Chemical Company, Ann Arbor, MI) specifications and absorbance was read at 340 nm using a plate reader (Synergy HT, Bio Tek Instruments, USA).
Apoptosis: Histone-related DNA fragmentation was used to determine the induction of apoptosis by ginger extracts.
DNA fragmentation: To determine the effect of selected ginger extracts in induction of apoptosis, 1×104 HepG2 cells were treated with ginger extracts for 12 and 24 h. Cells were washed with PBS after treatment. Cell lysates were prepared using cell lysis buffer and centrifuged at 200 g for 10 min. Apoptosis was determined by measuring internucleosomal DNA fragmentation using a cell death detection ELISA kit (Cell Death Detection ELISA, Roche Molecular Biochemicals, Indianapolis, IN) according to the instructions provided by the manufacturer. Spectrophotometric analysis was carried out using a plate reader (Synergy HT, Bio Tek Instruments, USA) at 405 nm to measure the histone related DNA fragmentation.
Caspase-3 activity: Cells were treated with extracts (30-240 μg mL-1) for 12 and 24 h. Cells were then extracted with PBS (pH 7) and centrifuged at 10,000×g for 15 min. Cell lysis buffer was added to the pellet, incubated for 10 min and centrifuged at 15,000×g for 15 min. The supernatant was collected and Caspase-3 activity was determined using a caspase kit (Promega Corporation, Madison, WI) according to the manufacturer’s instructions.
RESULTS
Cytotoxicity in HepG2: Percentage cytotoxicity (LDH release) in HepG2 cells incubated for 12 and 24 h with FG, OD, SD and FD ginger is shown in Fig. 1. The lowest LDH release was seen in cells treated with FG after 12 and 24 h incubation. However, the highest % cytotoxicity was seen in cells treated with SD (60 μg mL-1 (24 h)). After 12 h of incubation, cytotoxicity (%) ranged from 1.54-12.53 in cells treated with FG (120 μg mL-1) which was lower compared to 60 and 240 μg mL-1. The LDH release in HepG2 cells treated with OD ginger extracts was time dependent with a higher release seen after 24 h compared to 12 h. The highest LDH release (21.56) was seen after 24 h (60 μg mL-1). LDH release was lower at the 2 highest concentrations (120 and 240 μg mL-1) compared to 60 μg mL-1.
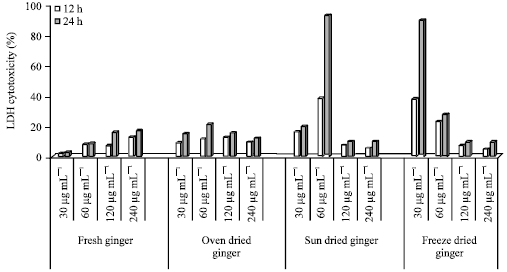 | |
| Fig. 1: | Percentage cytotoxicity of ginger-treated HepG2 cells treated with ginger extracts for 12 and 24 h |
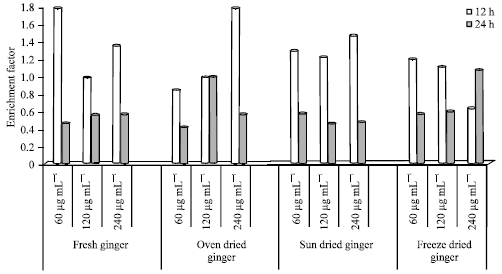 | |
| Fig. 2: | Histone related DNA fragmentation in HepG2 cells treated with ginger extracts for 12 and 24 h |
The highest (%) cytotoxicity (92.98) was seen in cells treated with SD (60 μg mL-1) for 24 h. The LDH release at 60 μg mL-1 was 38% at 12 h, however more than double (92.89) after 24 h of incubation. Incubation with the 2 highest doses of 120 and 240 μg mL-1 resulted in the lowest cytotoxicity after both 12 and 24 h of incubation.
The LDH release was highest at the lowest concentration (30 μg mL-1) of FD after both 12 and 24 h. The cells incubated with FD ginger had the lowest LDH release at the 2 highest concentrations (120 and 240 μg mL-1).
Histone related DNA fragmentation in HepG2 cells: Histone related DNA fragmentation (enrichment factor) in HepG2 cells is shown in Fig. 2. The FG, OD, SD and FD extracts were added to HepG2 cells at 60, 120 and 240 μg mL-1 and incubated for 12 and 24 h.
The Enrichment Factor (EF), with the exception of FD (240 μg mL-1), was higher at 12 h compared to 24 h of incubation. The lowest EF was 0.41 for OD extracts at 60 μg mL-1 (12 h) and the highest was 1.78 for OD extracts at 240 μg mL-1 (24 h). The EF for FG after 12 h of incubation ranged from 0.98-1.77. However, EF after 24 h ranged from 0.46-0.57. The EF for FG (120 μg mL-1) after 12 h of incubation was lower compared to 60 and 240 μg mL-1. The EF for FG at 24 h increased marginally with increasing concentration from 0.46, 0.56, 0.57 at 60, 120 and 240 μg mL-1.
The EF for cells treated with OD ginger extracts ranged from 0.84-1.78 and 0.42-0.99 for 12 and 24 h incubation, respectively. There was marginal difference in EF after 12 h incubation in cells treated with 60 and 120 μg mL-1. However, the EF at 240 μg mL-1 (12 h) was higher (1.78). The EF for OD ginger extracts after 24 h of incubation was higher at 120 μg mL-1 compared to 60 and 240 μg mL-1.
For cells treated with SD ginger extracts, the EF ranged from 1.22-1.46 and 0.58-0.45 at 12 and 24 h incubation, respectively. However, the EF was lower at 120 μg mL-1 compared to 60 and 240 μg mL-1 (12 h incubation). The EF (indicating apoptosis/DNA fragmentation) was higher at 60 μg mL-1 compared to 120 and 240 μg mL-1 at 24 h incubation. The EF was higher at 12 h compared to 24 h at all concentrations. The EF for FD ginger extracts was higher after 12 h compared to 24 h incubation at 60 and 120 μg mL-1.
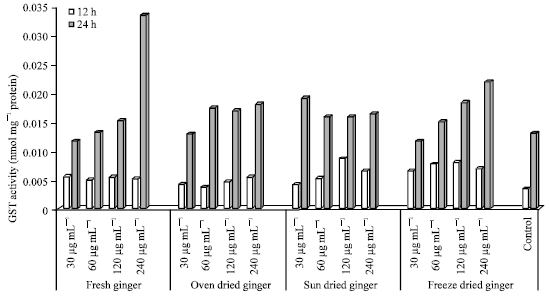 | |
| Fig. 3: | GST activity in HepG2 cells treated with ginger extracts for 12 and 24 h |
However, a reverse was seen at 240 μg mL-1. At 12 h incubation, there was a marginal difference between the EF at 60 and 120 μg mL-1. Cells incubated with FD at 240 μg mL-1 had a lower EF compared to FD concentrations at 60 and 120 μg mL-1 (12 h). The EF for cells treated with FD ginger extracts was 1.08; the highest EF for HepG2 cells incubated at 24 h for processed (OD, SD) and FG (control).
GST activity in HepG2 cells: GST activity was highest (0.03343) after 24 h incubation with FG extracts at 240 μg mL-1 (0.03 nmol mg-1 protein) and lowest in the control after 12 h incubation (0.003 nmol mg-1 protein) as showed in Fig. 3.
GST activity in HepG2 cells treated with ginger extracts (30-240 μg mL-1) for 12 and 24 h is shown in Fig. 3. The highest GST activity was seen in cells treated with FG at 240 μg mL-1 after 24 h of incubation. The control (untreated) cells after 12 h had the lowest GST activity compared to other treatments.
The GST activity in cells treated with FG (after 12 h of incubation) was similar at the 4 concentrations tested (30-240 μg mL-1). There was however a small increase seen with increasing concentrations of FG (30-120 μg mL-1). A 2-fold increase in GST activity was however seen at the highest concentration (240 μg mL-1) after 24 h.
In HepG2 cells treated with ginger extract (OD), higher GST activity was seen after 24 h of incubation compared to 12 h. The lowest GST activity in cells treated with OD (24 h) was seen at 30 μg mL-1.
In cells treated with SD ginger, the GST activity was again higher at 24 h compared to 12 h of incubation, the highest GST activity was seen in cell treated with SD at 120 μg mL-1. However, at 24 h GST activity was highest at 30 μg mL-1 incubation.
In cells treated with FD ginger, there were small increases seen in GST activity from 30-240 μg mL-1 after 24 h (dose response effect seen). However, GST activity was lowest at the highest concentration (240 μg mL-1) in cells treated with FD ginger (after 12 h). The untreated (control) cells had lower GST activity after both 12 and 24 h compared to the FG, OD, SD and FD treated cells.
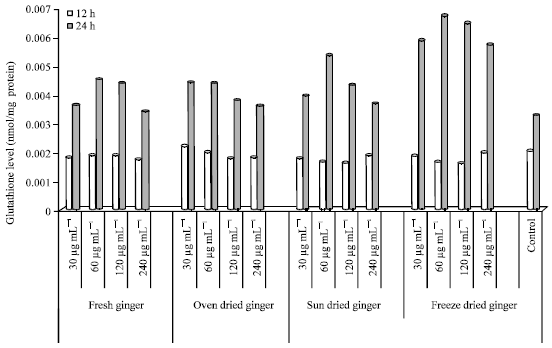 | |
| Fig. 4: | Glutathione level in HepG2 cells treated with ginger extracts for 12 and 24 h |
GSH level in HepG2 cells: The GSH levels in HepG2 cells are showed in Fig. 4, after incubation with FG, OD, SD and FD ginger extracts. GSH level (a substrate for detoxification enzymes) was higher after 24 h incubation in FD (60 μg mL-1) and lowest in the control (untreated cells). However, the highest GSH level in HepG2 cells after 12 h incubation was seen in OD ginger (30 μg mL-1) and the lowest in FD (120 μg mL-1).
Incubation with FG after 12 h resulted in highest GSH level at 60 μg mL-1 and there were only slight differences among the selected concentrations. The GSH level increased from 30-60 μg mL-1. However, GSH levels decreased from 60-240 μg mL-1 after 24 h of incubation and there was only a small difference in GSH levels in cells at 60 and 120 μg mL-1.
OD ginger extracts resulted in higher GSH levels at 30 μg mL-1 compared to 60, 120 and 240 μg mL-1 after 12 h of incubation. There were slight differences between the GSH levels at 120 and 240 μg mL-1. After 24 h of incubation, the GSH level was higher at 30 μg mL-1 compared to 60, 120 and 240 μg mL-1.
The GSH levels in cells treated with SD ginger were highest at 60 μg mL-1 (24 h). However, GSH levels decreased from 60-240 μg mL-1 after 24 h incubation concentrations (30, 60, 120 and 240 μg mL-1).
Incubation with FD ginger after 12 h of incubation resulted in the highest GSH level at 240 μg mL-1. However, there were only small differences in GSH levels at 30, 60, 120 and 240 μg mL-1. The GSH levels after 24 h incubation increased from 30-60 μg mL-1 but decreased from 60-240 μg mL-1. GSH levels were similar at the lowest (30 μg mL-1) and highest (240 μg mL-1) concentrations after 24 h. The GSH level in control (untreated cells) after 12 h incubation was either higher or similar compared to FG, OD, SD and FD. However, GSH level after 24 h incubation was lower compared to the processed and unprocessed ginger.
SOD activity in HepG2 cells: SOD activity in HepG2 cells after 12 and 24 h incubation with selected concentrations of FG, OD, SD and FD is shown in Fig. 5.
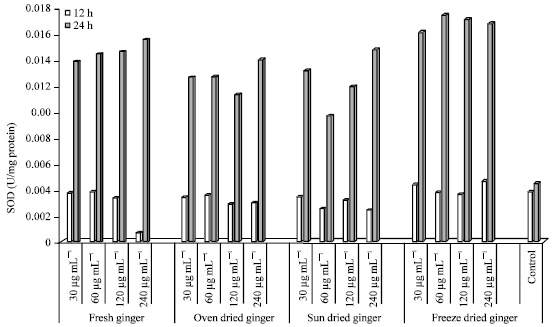 | |
| Fig. 5: | SOD activity in HepG2 cells treated with ginger extracts for 12 and 24 h |
The SOD activity was lowest in cells treated with FG (12 h) at the highest concentrated (240 μg mL-1). There were small differences seen in SOD activity in cells treated with 30-120 μg mL-1 (12 h). After 24 h of incubation, the highest SOD activity was seen at the highest concentration (240 μg mL-1), with small difference seen in cells incubated with 30-120 μg mL-1.
The SOD activity in cells treated with OD was higher at 24 h compared to 12 h of incubation. SOD was similar after (after 12 h) at all concentrations tested (30-240 μg mL-1). However, at 24 h of incubation, SOD activity was highest (0.01401) at the highest concentration (240 μg mL-1). SOD activity was higher at the lowest concentration compared to cells incubated with 60 and 120 μg mL-1.
A similar response in SOD activity was seen in cells treated with SD ginger after 24 h of incubation. The highest activity was seen at the highest concentration (240 μg mL-1), however the lowest SOD activity was seen in cells treated with 60 μg mL-1 (24 h). The SOD activity (U/mg protein) (after 12 h) was lower in cells incubation with 30 and 120 μg mL-1. Incubation with SD ginger extracts resulted in higher SOD activity after 24 h compared to 12 h of incubation. Among all the ginger extracts used, incubation with FD ginger resulted in the highest SOD activity (U/mg protein) after 24 h of incubation, with the highest activity seen in cells treated with 60 μg mL-1.
Incubation with FD ginger resulted in lower SOD activity at 12 h with very small differences seen at all concentrations treated (30-240 μg mL-1). The SOD activity in control (untreated) cells after 12 h was either similar or even higher compared to the treated cells. However, all treated cells (30-240 μg mL-1) had higher SOD activities compared to the control (after 24 h of incubation).
Catalase activity in HepG2 cells: Catalase (CAT) activity after 12 and 24 h incubation is shown in Fig. 6. CAT activity was higher at 24 h compared to 12 h of incubation in most treated cells (except FG 60 μg mL-1 and SD 30 μg mL-1).
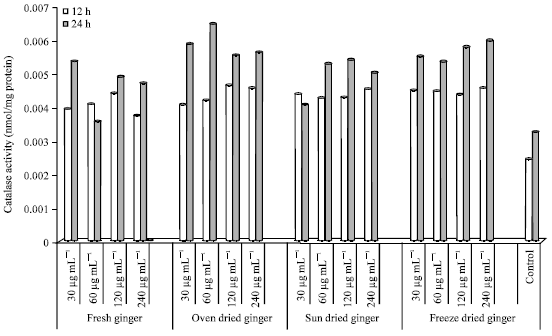 | |
| Fig. 6: | Catalase activity in HepG2 cells treated with ginger extracts for 12 and 24 h |
Cells treated with FG showed a dose response effect from 30 to 120 μg mL-1 (12 h). CAT activity however, decreased from 0.0044 (120 μg mL-1) to 0.0038 (240 μg mL-1) (12 h). After 24 h of incubation with FG extracts, CAT activity was lowest in cells treated with 60 μg mL-1 and highest at the lowest concentration (30 μg mL-1).
Cells treated with OD ginger extracts had the highest CAT activity at 60 μg mL-1 (24 h); a 2-fold increase compared to the control. A dose dependent relationship was seen after 12 h incubation at concentrations of 30, 60 and 120 μg mL-1. However, after 24 h incubation there was an increase seen at concentrations of 30 and 60 μg mL-1. However, CAT activity was similar at 120 and 240 μg mL-1 after 12 and 24 h incubation.
CAT activity in SD treated cells ranged from 0.0043-0.0045 (12 h) and 0.0041-0.0054 (24 h). The highest CAT enzyme activity (24 h) was seen at a concentration of 120 μg mL-1. Cells treated with SD extracts (60-240 μg mL-1) had higher CAT activities after 24 h compared to 12 h.
HepG2 cells incubated for 12 and 24 h with FD ginger extracts had the highest CAT activity at 240 μg mL-1 (24 h). However, cells treated with FD at 60 μg mL-1 had the lowest CAT activity (24 h). Similar CAT enzyme activity was seen at all the selected concentrations (12 h) which ranged from 0.0043-0.0045 nmol/mg protein.
GPx activity in HepG2 cells: GPx activity in HepG2 cells after 12 and 24 h incubation with ginger extracts (30-240 μg mL-1) is shown in Fig. 7. GPx activity (protect cells from oxidative damage) was higher at 24 h compared to 12 h incubation period except in cells treated with OD ginger (120 and 240 μg mL-1). The control (24 h) had the highest GPx activity compared to cells incubated with selected concentrations of ginger extracts. However, the control (12 h) had a higher GPx activity compared to all concentrations except OD (120 and 240 μg mL-1).
Incubation with FG extracts resulted in the highest GPx activity after 12 h in cells treated with 60 μg mL-1. A decrease in GPx activity was seen from concentrations of 60, 120 and 240 μg mL-1. GPx activity ranged from 0.0195-0.0080 (12 h). After 24 h incubation, GPx activity decreased from 0.0435-0.0212 in cells treated with FG (30-240 μg mL-1).
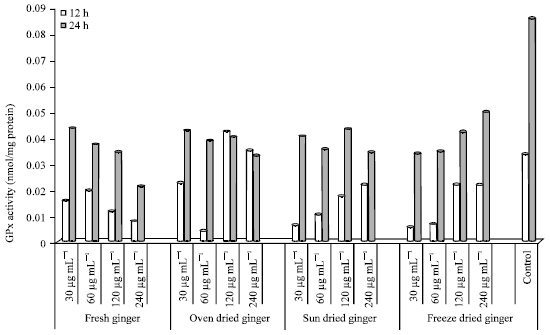 | |
| Fig. 7: | GPx activity in HepG2 cells treated with ginger extracts for 12 and 24 h |
The lowest GPx activity was seen in cells treated with OD 60 μg mL-1 (0.0044); an 8-fold decrease compared to the control. The highest GPx activity was seen in cells treated with 30 μg mL-1 (24 h) and 120 μg mL-1 (12 h). The GPx activity was lower in cells treated with 120 and 240 μg mL-1 at 24 h compared to 12 h.
A dose response effect was seen in cells treated with SD at 30, 60, 120 and 240 μg mL-1 (12 h) which ranged from 0.0062-0.0223. The GPx activity was highest after incubation with OD ginger (120 μg mL-1) after 24 h. GPx activity in cells treated with 60 μg mL-1 (0.0356) and 240 μg mL-1 (0.0342) were similar (24 h).
Gpx activity in cells treated with FD (24 h) also showed a dose effect from concentrations of 30-240 μg mL-1 which ranged from 0.0343-0.0501. The highest GPx activity in cells treated with FD ginger (240 μg mL-1) was 1.7-fold lower compared to the control (24 h).
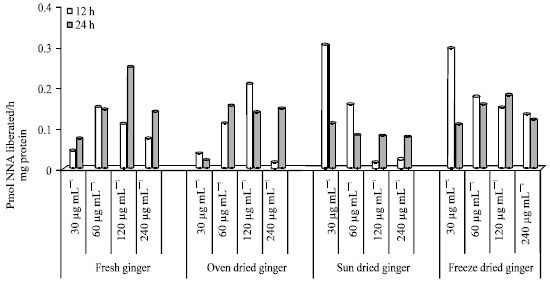
Glutathione Reductase (GR) activity in HepG2 cells: The GR activity in HepG2 cells treated with different concentrations of FG, OD SD and FD for 12 and 24 h is shown in Fig. 8.
The GR activity in control (untreated cells) was higher after 24 h compared to 12 h of incubation. FG treated cells incubated after 24 h showed a decrease in dose response from concentrations of 30-240 μg mL-1, ranging from 25.17-10.57 nmol mg-1 protein. However, the reverse was seen in SD (0.827-24.72). After 12 h incubation, a decrease in GR activity in OD (30, 60, 120 and 240 μg mL-1) was seen which ranged from 18.258-2.586. The lowest GR activity was seen in cells (12 h) treated with FG at 60 μg mL-1, however, the highest activity (24 h) was seen in cells treated with 30 μg mL-1 FG. The lowest activity after 24 h was seen in cells treated with 30 and 60 μg mL-1.
Caspase-3 activity in HepG2: Caspase-3 activity an indicator of apoptosis is shown in Fig. 9. The caspase activity in HepG2 cells incubated with FG was higher after 24 h compared to 12 h of incubation, except for hepG2 cells incubated at a concentration of 60 μg mL-1. After 24 h of incubation, there was a dose-dependent increase in caspase activity in cells incubated with fresh ginger extracts at concentrations of 30-60 μg mL-1. which ranged from 0.044-0.075.
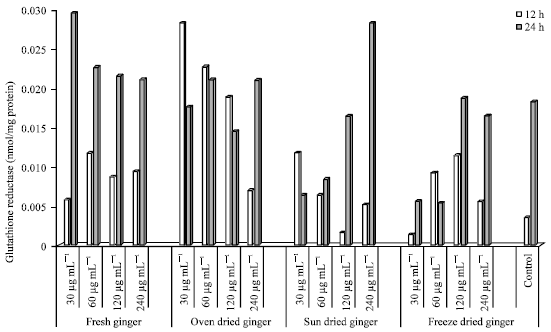 | |
| Fig. 8: | Glutathione reductase activity in HepG2 cells treated with ginger extracts for 12 and 24 h |
 | |
| Fig. 9: | Caspase-3 activity in HepG2 cells treated with ginger extracts for 12 and 24 h |
However, caspase activity was lower in cells incubated in extract concentration of 240 μg mL-1 compared to cells incubated in extracts concentration of 120 μg mL-1. A similar trend was seen after 12 h of incubation, with the highest caspase activity in cells incubated at a concentration of 60 μg mL-1.
For the processed ginger; OD, SD and FD, there was a dose-dependent decrease after 12 h of incubation in SD and FD at all concentrations. However, the reverse was seen in cells incubated with SD and FD at 30 μg mL-1.
DISCUSSION
Epidemiological studies have shown that phytochemicals have the capacity to interfere in the development of chronic diseases, such as cancer (Abdul Hanif et al., 2005). The anticarcinogenic, antitumor and antiproliferation properties of ginger has been widely studied (Kundu et al., 2009). Furthermore, epidemiological studies suggest that the consumption of spices may be a factor in reducing cancer occurrences (Alia et al., 2005).
The present study evaluated the cytotoxicity and antiproliferative properties of treating Hep2G cells with processed ginger extracts, specifically as it related to LDH release, antioxidative and detoxification enzyme activity and mechanism of cell death. HepG2 cells were incubated over a period of 12 and 24 h with processed ginger, with Fresh Ginger (FG) as the control at concentration of 30-240 μg mL-1. Enzymes measured included SOD, GPx, CAT, GSH GR and GST. Uncontrollable proliferation of cells is one characteristic of cancer (Hanahan and Weinberg, 2000); this study showed that ginger extracts (processed and unprocessed) exhibited a time-dependent cytotoxicity, thus preventing further proliferation. Results showed that processed ginger had greater cytotoxicity at lower concentrations compared to fresh ginger. A decrease in LDH release is an indicator of increased number of dead cells as LDH is only released when living cells are damaged (Decker and Lohmann-Matthes, 1988). The effect of concentration on cytotoxity in HepG2 cells were similarly observed by Granado-Serrano et al. (2006). Granado-Serrano et al. (2006) studied HepG2 cells incubated with quercetin. Ginger has been shown to have high amount quercetin (Ghasemzadeh et al., 2010).
The enrichment factor levels from HepG2 cells after incubation is an indicator of the antiproliferative properties of ginger. The enrichment factor (indicator of apoptosis) as shown in Fig. 2 was higher after 12 h of incubation except for HepG2 cells incubated with freeze dried ginger (240 μg mL-1) which was similarly reported by Sakinah et al. (2007) in Zerumbone induced apoptosis in liver cancer cells.
Glutathione linked enzymes: Glutathione-S-Transferase (GST), Glutathione Reductase (GR) and glutathione peroxidase (GPx) are involved in the modulation toxicity (Velmurugan et al., 2001). GST a phase II detoxification enzyme uses GSH as substrate to conjugate electrophilic molecules. On the other hand, GR catalyzes the reduction of GSSH to GSH (Liska et al., 2006; Williams et al., 2007). Figure 3 showed higher GST activity in HepG2 after 24 compared to 12 h of incubation. Williamson et al. (1997) reported similar results in HepG2 incubated with extracts of cruciferous vegetables.
The antioxidative enzymes studied were superoxide dismutase (SOD), GPx and Catalase (CAT) as shown in Fig. 5-7, respectively. The removal of superoxide and peroxides in cells are catalyzed by SOD, GPx and CAT (Mates et al., 1999). There was an increase in antioxidant enzymes activity after 24 h incubation. Shivananjappa et al. (2013) reported similar increase in antioxidative enzymes in HepG2 cell after incubation with Terminalia arjuna an initial decrease in SOD activity after 12 h of incubation may probably due to a reduction in the production of superoxides. After 24 h, a dose-dependent increase in SOD activity in Hepg2 cells incubated with OD and SD was seen at concentrations 60-240 μg mL-1. However, the SOD activity in cells incubated at concentration of 30 μg mL-1 (OD and SD) was higher compared to enzyme activity at concentrations of 60 and 120 μg mL-1.
CAT and GPx catalyze the decomposition of hydrogen peroxide to less toxic compounds (Alia et al., 2005). Increased induction of CAT and GPx enzymes activity (Fig. 6 and 7) may have occurred due to the production of hydrogen peroxide after 24 h compared to 12 h of incubation. The GPx activity in the control was higher compared to cells incubated with ginger extracts may be due to the accumulation of hydrogen peroxides. There was a dose-dependent decrease in GSH in cells incubated with FD ginger for 12 h as shown in Fig. 4 which corresponded to an increase in the GST and GPx. This trend is attributed to GSH being the substrate for GST and GPx enzymes (Liska et al., 2006).
Caspase activity, an indicator, of apoptosis was higher at lower concentration in processed and unprocessed ginger after 12 h compared to 24 h of incubation, there was a similar trend in the measure LDH (percentage cytotoxity).
CONCLUSION
In conclusion, this study HepG2 cells incubated over a period of 12 and 24 h with processed ginger; Oven Dried (OD), Sun Dried (SD) and Freeze Dried (FD) with Fresh Ginger (FG) as the control at concentration of 30-240 μg mL-1 showed a decrease in antioxidant (GPx) as the control was higher compared to the treated cells. Ginger extracts also exhibited cytotoxity and the induction of apoptosis.
ACKNOWLEDGMENT
This project was funded by Alabama Agricultural Experimental Research Station, Normal AL 35762.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Aggarwal, B.B. and S. Shishodia, 2006. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol., 71: 1397-1421.
CrossRefPubMedDirect Link - Ali, B.H., G. Blunden, M.O. Tanira and A. Nemmar, 2008. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol., 46: 409-420.
CrossRefPubMedDirect Link - Alia, M., S. Ramos, R. Mateos, L. Bravo and L. Goya, 2005. Response of the antioxidant defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2). J. Biochem. Mol. Toxicol., 19: 119-128.
CrossRefDirect Link - Altekruse, S.F., K.A. McGlynn and M.E. Reichman, 2009. Hepatocellular carcinoma incidence, mortality and survival trends in the United States from 1975 to 2005. J. Clin. Oncol., 27: 1485-1491.
CrossRefDirect Link - Ames, B.N., 1989. Endogenous oxidative DNA damage, aging and cancer. Free Radical Res. Commun., 7: 121-128.
CrossRefPubMedDirect Link - Bartley, J.P. and A.L. Jacobs, 2000. Effects of drying on flavour compounds in Australian-grown ginger (Zingiber officinale). J. Sci. Food Agric., 80: 209-215.
CrossRefDirect Link - Coyle, J.T. and P. Puttfarcken, 1993. Oxidative stress, glutamate and neurodegenerative disorders. Science, 262: 689-695.
CrossRef - Decker, T. and M.L. Lohmann-Matthes, 1988. A quick and simple method for the quantitation of lactate dehydrogenase release in measurements of cellular cytotoxicity and Tumor Necrosis Factor (TNF) activity. J. Immunol. Methods, 115: 61-69.
CrossRefDirect Link - Ghasemzadeh, A., H.Z.E. Jaafar and A. Rahmat, 2010. Antioxidant activities, total phenolics and flavonoids content in two varieties of malaysia young ginger (Zingiber officinale Roscoe). Molecules, 15: 4324-4333.
CrossRefPubMedDirect Link - Granado-Serrano, A.B., M.A. Martin, L. Bravo, L. Goya and S. Ramos, 2006. Quercetin induces apoptosis via caspase activation, regulation of Bcl-2 and inhibition of PI-3-kinase/Akt and ERK pathways in a human hepatoma cell line (HepG2). J. Nutr., 136: 2715-2721.
PubMedDirect Link - Halliwell, B. and J.M.C. Gutteridge, 1995. The definition and measurement of antioxidants in biological systems. Free Radic. Biol. Med., 18: 125-126.
CrossRefPubMedDirect Link - Hanahan, D. and R.A. Weinberg, 2000. The hallmarks of cancer. Cell, 100: 57-70.
CrossRefPubMedDirect Link - Abdul Hanif, H., N.A. Murad, W.Z.W. Ngah and Y.A.M. Yusof, 2005. Effects of zingiber officinale on superoxide dismutase, glutathione peroxidase, catalase, glutathione and malondialdehyde content in hepg2 cell line. Malaysian J. Biochem. Mol. Biol., 11: 36-41.
Direct Link - Harman, D., 1992. Role of free radicals in aging and disease. Ann. N. Y. Acad. Sci., 673: 126-141.
CrossRefPubMedDirect Link - Hatono, S., A. Jimenez and M.J. Wargovich, 1996. Chemopreventive effect of s-allylcysteine and its relationship to the detoxification enzyme glutathione s-transferase. Carcinogenesis, 17: 1041-1044.
PubMedDirect Link - Kundu, J.K., H.K. Na and Y.J. Surh, 2009. Ginger-derived phenolic substances with cancer preventive and therapeutic potential. Forum Nutr., 61: 182-192.
CrossRefPubMedDirect Link - Lee, E. and Y.J. Surh, 1998. Induction of apoptosis in HL-60 cells by pungent vanilloids, [6]-gingerol and [6]-paradol. Cancer Lett., 134: 163-168.
CrossRefPubMedDirect Link - Liska, D., M. Lyon and D.S. Jones, 2006. Detoxification and biotransformational imbalances. Explore: J. Sci. Healing, 2: 122-140.
CrossRefDirect Link - Liu, R.H., 2003. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr., 78: 517S-520S.
CrossRefPubMedDirect Link - Loft, S. and H.E. Poulsen, 1996. Cancer risk and oxidative DNA damage in man. J. Mol. Med., 74: 297-312.
PubMed - Machlin, L.J. and A. Bendich, 1987. Free radical tissue damage: Protective role of antioxidant nutrients. FASEB J., 1: 441-445.
CrossRefPubMedDirect Link - Mates, J.M., 2000. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology, 153: 83-104.
CrossRefPubMedDirect Link - Mates, J.M., C. Perez-Gomez and I.N. de Castro, 1999. Antioxidant enzymes and human diseases. Clin. Biochem., 32: 595-603.
CrossRefPubMedDirect Link - Meister, A., 1988. Glutathione metabolism and its selective modification. J. Biol. Chem., 263: 17205-17208.
PubMedDirect Link - Nicoll, R. and M.Y. Henein, 2009. Ginger (Zingiber officinale Roscoe): A hot remedy for cardiovascular disease? Int. J. Cardiol., 131: 408-409.
CrossRef - Oberley, L.W. and G.R. Buettner, 1979. Role of superoxide dismutase in cancer: A review. Cancer Res., 39: 1141-1149.
PubMedDirect Link - Russo, G.L., 2007. Ins and outs of dietary phytochemicals in cancer chemoprevention. Biochem. Pharmacol., 74: 533-544.
CrossRefPubMedDirect Link - Sakinah, S.A., S.T. Handayani and L.P. Hawariah, 2007. Zerumbone induced apoptosis in liver cancer cells via modulation of bax/bcl-2 ratio. Cancer Cell Int., Vol. 7.
CrossRefDirect Link - Salmon, C.N.A., Y.A. Bailey-Shaw, S. Hibbert, C. Green, A.M. Smith and L.A.D. Williams, 2012. Characterisation of cultivars of jamaican ginger (Zingiber officinale Roscoe) by HPTLC and HPLC. Food Chem., 131: 1517-1522.
CrossRefDirect Link - Surh, Y.J., 2003. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer, 3: 768-780.
CrossRefPubMedDirect Link - Surh, Y.J., K.S. Chun, H.H. Cha, S.S. Han, Y.S. Keum, K.K. Park and S.S. Lee, 2001. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res./Fund. Mol. Mech. Mutagen., 480-481: 243-268.
CrossRefDirect Link - Surh, Young-Joon, J.K. Kundu and Hye-Kyung Na, 2008. Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals. Planta Med., 74: 1526-1539.
CrossRefPubMedDirect Link - Valko, M., M. Izakovic, M. Mazur, C.J. Rhodes and J. Telser, 2004. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem., 266: 37-56.
CrossRefDirect Link - Velmurugan, B., V. Bhuvaneswari, S. Balasenthil and S. Nagini, 2001. Lycopene, an antioxidant carotenoid modulates glutathione-dependent hepatic biotransformation enzymes during experimental gastric carcinogenesis. Nutr. Res., 21: 1117-1124.
CrossRefDirect Link - Williams, D., M. Verghese, L.T. Walker, J. Boateng, L.T. Shackelford and C.B. Chawan, 2007. Flax seed oil and flax seed meal reduce the formation of Aberrant Crypt Foci (ACF) in azoxymethane-induced colon cancer in Fisher 344 male rats. Food Chem. Toxicol., 45: 153-159.
CrossRefPubMedDirect Link - Williamson, G., M.S. DuPont, S. Wanigatunga, R.K. Heaney, S.R.R. Musk, G.R. Fenwick and M.J.C. Rhodes, 1997. Induction of glutathione s-transferase activity in hepg2 cells by extracts from fruits and vegetables. Food Chem., 60: 157-160.
CrossRefDirect Link - Winyard, P.G., C.J. Moody and C. Jacob, 2005. Oxidative activation of antioxidant defence. Trends Biochem. Sci., 30: 453-461.
CrossRefDirect Link - Yu, R., S. Mandlekar, K.J. Harvey, D.S. Ucker and A.N. Kong, 1998. Chemopreventive isothiocyanates induce apoptosis and caspase-3-like protease activity. Cancer Res., 58: 402-408.
PubMedDirect Link - Zelko, I.N., T.J. Mariani and R.J. Folz, 2002. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2) and EC-SOD (SOD3) gene structures, evolution and expression. Free Radical Med. Biol., 33: 337-349.
CrossRefDirect Link