
C.M. Melgoza Villagomez
Laboratorio de Biotecnologia Aplicada, Instituto Tecnologico El Llano, Aguascalientes, km 18 Carr, Aguascalientes-San Luis Potosi, CP 20330, Mexico
H. Silos Espino
Laboratorio de Biotecnologia Aplicada, Instituto Tecnologico El Llano, Aguascalientes, km 18 Carr, Aguascalientes-San Luis Potosi, CP 20330, Mexico
A. Nava Cedillo
Laboratorio de Biotecnologia Aplicada, Instituto Tecnologico El Llano, Aguascalientes, km 18 Carr, Aguascalientes-San Luis Potosi, CP 20330, Mexico
L.L. Valera Montero
Laboratorio de Biotecnologia Aplicada, Instituto Tecnologico El Llano, Aguascalientes, km 18 Carr, Aguascalientes-San Luis Potosi, CP 20330, Mexico
C. Perales Segovia
Laboratorio de Biotecnologia Aplicada, Instituto Tecnologico El Llano, Aguascalientes, km 18 Carr, Aguascalientes-San Luis Potosi, CP 20330, Mexico
C.A. Martinez Guerra
Rancho San Carlos, Ejido El Tule, Asientos, Aguascalientes, Mexico
G. Martinez Guerra
Rancho San Carlos, Ejido El Tule, Asientos, Aguascalientes, Mexico
A. Cruz Hernandez
Centro de Investigacion and de Estudios Avanzados del Instituto Politecnico Nacional, Unidad Guanajuato, Apdo, Postal 629, Irapuato, Gto, Esp 36500, Mexico
O. Paredes Lopez
Centro de Investigacion and de Estudios Avanzados del Instituto Politecnico Nacional, Unidad Guanajuato, Apdo, Postal 629, Irapuato, Gto, Esp 36500, Mexico
International Journal of Botany
Year: 2009 | Volume: 5 | Issue: 2 | Page No.: 160-165







ABSTRACT
Twenty four peach genotypes from the Central North Region of Mexico, were characterized based on morpho-physiological traits. Fruit weights from the genotypes Roxana (135 g), San Gabriel C-167 (141.9 g) and Zacatecas landrace (162.3 g) were the higher, each in its group, since all genotypes were grouped according to their readiness to harvest as early-, middle- and late-harvest, respectively. RAPDs analysis yielded 52 monomorphic and 93 polymorphic fragments that were related to desirable characteristics from the Prunus genotypes. This information provide us tools for early individual identification of high-performance trees when still growing in the nursery. Therefore, growers may use this technique for assisted breeding program on their Prunus genotypes.
PDF Abstract XML References Citation
How to cite this article
C.M. Melgoza Villagomez, H. Silos Espino, A. Nava Cedillo, L.L. Valera Montero, C. Perales Segovia, C.A. Martinez Guerra, G. Martinez Guerra, A. Cruz Hernandez and O. Paredes Lopez, 2009. Identification of Peach Genotypes (Prunus persica (L.) Batsch) in the North-Central Region, Mexico. International Journal of Botany, 5: 160-165.
DOI: 10.3923/ijb.2009.160.165
URL: https://scialert.net/abstract/?doi=ijb.2009.160.165
DOI: 10.3923/ijb.2009.160.165
URL: https://scialert.net/abstract/?doi=ijb.2009.160.165
INTRODUCTION
Peach (Prunus spp.) is a fruit well appreciated either fresh or canned due to its flavor, color and aroma. Additionally, this is a low-calorie fruit, useful in the human diet as a source of potassium, vitamin A and ascorbic acid (Skorza and Okie, 1990). Peaches are grown in temperate climates both sides of the Equator between parallels 30 and 40° (Joshi and Bhutani, 1995). In Mexico, Spaniards started peach cultivation in some areas and since then, only recently peach production has doubled in several states such as Chihuahua, Estado de México, Morelos, Sonora, Tlaxcala, Guanajuato, Hidalgo, Michoacán, Aguascalientes and Nayarit (Anonymous, 2008). These early and recent genotype introductions resulted into heterogeneous landraces and recently imported cultivars. As a consequence of these genotype population, characteristics of budding, flowering, ripening, vigor, quality and yield are heterogeneous among orchards (Gutierrez-Acosta and Padilla-Ramirez, 2004). Furthermore, low yields (averaging 10.8 t ha-1) and high labor required during the usually short harvest period make orchard management somewhat difficult.
Traditionally, in some Mexican regions, local identification and characterization of the genus Prunus spp. was mainly based on morphological, physiological and fruit traits, originating multiple names for the same genotype and scarce contribution to breeding programs. Nowadays, statistics report the presence of 52 genotypes (from USA) in Chihuahua, 12 genotypes (11 landraces and one genotype from Brazil) in Estado de Mexico and 4 genotypes in Michoacán (Martinez, 2005). Among the landraces of the last two states, Diamond is one outstanding genotype which has been used for breeding by the Colegio de Posgraduados. In Aguascalientes, the landrace San Gabriel has been present in the local commercial orchards over 30 years and has been the source of new genotypes with fruit and highly appreciated by the consumers, fruit flesh tightly adhered to the stone and yellow skin (Gutierrez-Acosta and Padilla-Ramirez, 2004; Perez, 2006).
Normally, for a peach three to develop completely all of its morphological features, the minimum required time is about 3-4 years. Nevertheless, it is possible to identify Prunus plants with good characteristics, without waiting too long, using molecular markers link to agronomic characteristics (Dirlewanger et al., 1998; Warburton et al, 1996). These techniques are: RFLPs, RAPDs, AFLPs and SSRs (Martinez-Gomez and Sánchez-Perez, 2005). From these, RAPDs (Random Amplified Polymorphic DNAs) are suitable for basic methods since is simple and does not require radioactivity (Bardacki, 2001). This specific technique has been applied to Prunus in order to identify cultivar and species (Martins and Tenreiro, 2003; MirAli and Nabulsi, 2003; Baranek et al., 2006) and genetic maps assembly (Warburton and Bliss, 1996; Dettori et al., 2001). Due to the fact that in Mexico we have not applied these techniques systematically, the objective of this study is to identify of the best peach (Prunus persica) genotypes through molecular markers (from RAPDs) associated to morpho-physiological descriptors.
MATERIALS AND METHODS
Morpho-physiological data was obtained from 24 peach (Prunus persica) genotypes (Table 1) grown under similar management (pruning, fertilization and irrigation) on a commercial orchard in Aguascalientes, Mexico. Additionally, DNA samples required to perform RAPDs analyses were taken from the leaf tissue of each genotype. Trees grown on that orchard are a heterogeneous collection of crosses derived from genotypes obtained from the North-Central region of México, Brazil, USA and Spain (Table 1). Fifty nine morphological traits (such as growing pattern, number of fruits per branch, fruit abscission and maturation pattern, color, texture, shape, oBrix and presence/absence of protuberance, etc.) were used to construct a phenogram by the simple matching coefficient procedure (Magurran, 1988). Genomic DNA was extracted from the 24 Prunus genotypes and the samples were run on 0.8% agarose gel.
DNA concentration was measured with a spectrometer (model GBC Cintra 10e UV-visible) (Doyle and Doyle, 1990). RAPD reactions were performed in a 25 μL volume consisting of 10X buffer solution [10 mM Tris-HCl buffer (pH 8.0), 50 mM KCl2], 2.5 mM MgCl2, 2.5 unit of Taq DNA polymerase (Promega), 100 μM dNTP, 50 ng genomic ADN and 0.4 μM OPM series (Table 2) primer (Operon Technologies, Alameda, CA, USA) according to Williams et al. (1990). A total of 20 μL of mineral oil was placed over the reaction mixture (Williams et al., 1990). Amplifications were carried out in a DNA thermocycler (Model FPR0G02Y Techne Progene, England), with the following conditions: an initial denaturation step of 2 min at 94°C, followed by 35 cycles of 1 min at 94°C, 1 min at 35°C and 2 min at 72°C with a final extension step of 7 min at 72°C (Williams et al., 1990). Amplification products were analyzed by electrophoresis in a 1.2% agarose gel run at 100 V for 4 h and detected by staining the gel with ethidium bromide (10 ng/100 mL of agarose solution in TBE) (Williams et al., 1990). All visible and unambiguous fragments amplified by the primers chosen were entered under the heading of total visible fragments. Fragment data were entered on a spreadsheet to form a binary matrix, where (1) represented fragment presence and (0) absence for each fragment-accession combination. Cluster analysis was conducted by converting the data matrix into a similarity matrix using a simple matching coefficient (Nei, 1973). This coefficient was calculated by dividing the number of matches (0-0 and 1-1) by the total number of comparisons (Nei and Li, 1979). A cluster analysis was then done using the unweighted pair group method, with arithmetical averages (UPGMA) process in the S-Professional Plus 2000 program.
| Table 1: | Morpho-physiological and productive description in the genotypes |
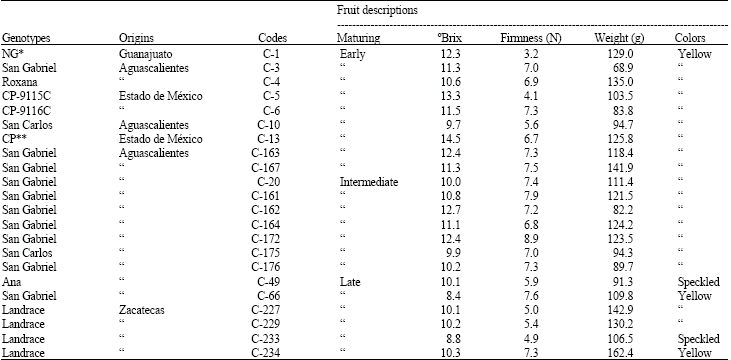 | |
| *Norte de Guanajuato genotype, **Colegio de Postgraduados genotype | |
| Table 2: | Amplified fragments from 24 peach (Prunus persica L. Batsch) genotypes by means of 14 primers series OPM grown in Central North Mexico |
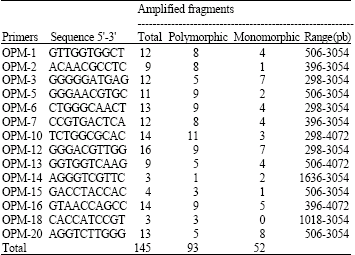 | |
RESULTS AND DISCUSSION
Morpho-physiological traits and productivity analysis: Table 1 shows a brief description of the used genotypes according to investigated traits such as place of origin, ripening, οBrix, fruit firmness, weight and color. Identified genotypes as early maturing were harvested between 15th of June and 15th of July; intermediate genotypes were harvested between the 16th of July and 15th of August; and late maturing genotypes were harvested after the 16th August. Comparing among the genotypes of the early maturing group, Roxana was found superior on fruit quality and higher yields (41.6 kg tree-1). Likewise, San Gabriel (C-161) was the best among the genotypes from intermediate maturing group with yields of 54.67 kg tree-1 and good fruit weight (121.5 g). In the late maturing group, Zacatecas landrace (C-234) showed the highest fruit weight (162.3 g); although, yields were poor (13.64 kg tree-1).
Two groups arose from the morphological trait analysis were obtained (data no showed). San Gabriel (C-3) was placed in Group I; nevertheless, this genotype seemed to be independent since it showed protuberance, high abscission rate, erectile plant growth, small fruits (60.7 g fruit-1) and shorter maturation period as compared to other genotypes within the early maturation group. Group II was characterized by horizontal branching, higher fruits and different maturation behavior. This morphological analysis allowed finding the relationship between genotypes obtained from different sites from Aguascalientes and Zacatecas. As an example, San Gabriel (C-167) and Zacatecas landrace C-175 are different only on the maturation period the rest of the characteristics appeared the same; furthermore, San Gabriel coded as C-175 and C-176 were very similar in both maturation and fruit firmness.
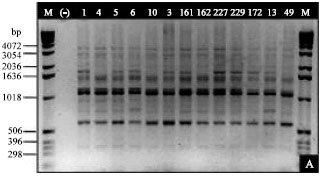 | |
 | |
| Fig. 1: | RAPD analysis on peach genotypes (Prunus persica L. Batsch) grown by means of 14 primers OPM series: OPM-10 (A) and OPM 12 (B) in Central North Mexico region |
Other findings were: both genotypes C-1 and C-229 (from Guanajuato and Zacatecas, respectively) showed the lowest fruit firmness. Genotypes C-164 and C-172 (San Gabriel) were slightly different on οBrix. Finally, Roxana (C-4) and San Gabriel (C-163) were similar on the maturation period.
RAPDs analysis results: Amplification of 14 fragments was obtained using primers OPM-12, OPM-10 and OPM-16, while using primers OPM-14 and OPM-18 yielded only 3 discrete bands each (Table 2). In general, amplicons ranged approximately from 300 to 4070 bp in size. Some of them were found to be genotype-specific as shown in Table 2 and Fig. 1a, b. Genotype-specific fragments shown were for genotype C-277 (3054 bp amplified with OPM-18) and genotypes C-175 and C-234 (5054, 2036 bp, respectively) amplified with OPM-13 (no showed). Fortunately, the above mentioned genotypes have good qualities for breeding (Table 1). Once the dendrogram was obtained using only the polymorphic fragments, it was evident the presence of two main groups: Group I includes only to genotype C-279 which is a Prunus domestica (May Bell Plum) imported from South Carolina USA, while Group II includes to the rest of the genotypes (Prunus persica) (Fig. 2a).
 | |
| Fig. 2: | (a) Dendrogram obtained from 24 peach (Prunus persica) genotypes grown in the Central-North region of Mexico, (b) C-167 and (c) C-233 |
Genotype C-6 (NG) and C-227 (Zacatecas landrace) showed the similarity coefficients highest (0.97349). In spite of that similarity, these genotypes differ on fruit firmness and maturation period. Low similarity coefficients (0.6091 and 0.6122) were obtained when compared genotypes C-279 to genotypes C-6 (CP-9116C) and C-227 (Zacatecas landrace), respectively. Among peaches, the lowest similarity values (0.8584 and 0.8622) were found when genotype C-176 was compared to C-4 and C-161, respectively. These results would be indicative of the close genetic relatedness of the peach genotypes grown in the Central-Northern Region of Mexico.
DISCUSSION
Due to the high cross pollination rate present in the genus Prunus spp., new genotypes can be developed with higher yields or better fruit quality. This breeding process could be accelerated if assisted by molecular markers associated with desired traits (Jun et al., 2002), helping in the process of selecting candidates from germinating plants. At the same time, molecular marker analyses would help on standardization for genotype naming or labeling all over Mexican peach production regions, in order to avoid confusion and promoting conservation of valuable gene sources. This study is considered the one of the first attempts in Mexico for establishing genetic fingerprints on peach that may help on taking decisions for potential introduction of the appropriate cultivars for the specific regions. It may be a good start focusing on the genotypes with higher yields and good fruit quality such as Roxana (C-4) from the early maturing group, San Gabriel (C-172) from the intermediate maturing group and a Zacatecas landrace (C-234) from the late maturing group as model for peach breeding. Indeed, the remaining samples were differentiated and clustered together as a group of cultivars or genotypes without any significant relation to some botanical or agricultural traits as is indicated by Raddova et al. (2003) when analyzed peaches provide by different countries. Better distinguishing results were detected by means of other amplification strategy in the work of Jun et al. (2002) were they found RAPDS and SCAR markers linked to the flesh adhesion gene (F) in peach (Prunus persica) and Cheng (2007) classified the peach groups as yellow, honey, flat, red leaf, crisp, bitao and juicy. In the future, the local collection from peach (Prunus spp.) growers will be extended and probably will be used for further plant breeding (gene identification o genetic transformation) activities.
ACKNOWLEDGMENTS
Thanks to Eng. Leonel Quesada Parga and Aguascalientes peach growers for their support to C.M. Melgoza Villagómez while pursuing her Master´s degree at the ITEL. Our thanks to Eng. Pablo Arturo Alemán and Eng. Alberto Peralta Bringas for the help provided. Finally, we appreciate the great labor provided by Dr. Pérez y Dr. Rodríguez, since some genotypes were obtained by their peach breeding programs.
REFERENCES
- Baránek, M., J. Raddová and M. Pidra, 2006. Comparative analysis of genetic diversity in Prunus L. As revelated by RAPD and SSR markers. Scientia Hort., 108: 253-259.
CrossRefDirect Link - Bardakci, F., 2001. Random Amplified Polymorphic DNA (RAPD) markers. Turk. J. Biol., 25: 185-196.
Direct Link - Cheng, Z., 2007. Genetic characterization of different demes in Prunus persica revealed by RAPD markers. Scientia Hort., 111: 242-247.
CrossRefDirect Link - Dettori, M.T., R. Quarta and I. Verde, 2001. A peach linkage map integrating RFLPs, SSRs, RAPDs and morphological markers. Genome, 44: 783-790.
Direct Link - Dirlewanger, E., V. Pronier, C. Parvery, C. Rothan, A. Guye and R. Monet, 1998. Genetic linkage map of peach [Prunus persica (L.) Batsch] using morphological and molecular markers. Theor. Applied Genet., 97: 888-895.
CrossRefDirect Link - Jun, J.H., K.H. Chung, S.B. Jeong and H.J. Lee, 2002. Identification of RAPD and SCAR markers linked t the flesh adhesion gene F in peach (Prunus persica (L.) Batsch). J. Hort. Sci. Biotechnol., 77: 598-603.
Direct Link - Martinez-Gomez, R. and M. Sanchez-Perez, 2005. Application of recent biotechnologies to Prunus tree crop genetic improvement. Cien. Inv. Agric., 32: 73-96.
Direct Link - Martins, M., R. Tenreiro and M.M. Oliveira, 2003. Genetic relatedness of portuguese almond cultivars assessed by RAPD and ISSR markers. Plant Cell Rep., 22: 71-78.
CrossRefDirect Link - MirAli, N. and I. Nabulsi, 2003. Genetic diversity of almonds (Prunus dulcis) using RAPD technique. Scientia Hort., 98: 461-471.
CrossRefDirect Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Raddova, J., M. Baranek, I. Oukropec, M. Vachum and M. Pidra, 2003. RAPD analysis of peaches within Czech national collection. Czech. J. Genet. Plant Breed., 39: 113-119.
Direct Link - Warburton, M.L., V.L. Becerra-Velasquez, J.C. Goffreda and F.A. Bliss, 1996. Utility of RAPD markers in identifying genetic linkages to genes of economic interest in peach. TAG Theoret. Applied Genet., 93: 920-925.
CrossRefDirect Link - Warburton, M.L. and F.A. Bliss, 1996. Genetic diversity in peach (Prunus persica L. Batch) revealed by randomly amplified polymorphic DNA (RAPD) markers and compared to inbreeding coefficients. J. Am. Soc. Hortic. Sci., 121: 1012-1019.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Nei, M., 1973. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA., 70: 3321-3323.
PubMedDirect Link