
Halima, H. Salem
Santous University Medical College, China
Bahy A. Ali
Santous University Medical College, China
Tian H. Huang
Santous University Medical College, China
Qing D. Xie
Santous University Medical College, China
Biotechnology
Year: 2006 | Volume: 5 | Issue: 4 | Page No.: 413-420







ABSTRACT
The new Bacillus thuringiensis isolate with toxicity against snails was characterized in comparison with three B. thuringiensis standard strains representing three different serovars using Polymerase Chain Reaction (PCR), hybridization and protein analysis. The novel B. thuringiensis strain was preliminary bioassayed for its toxicity against infected, non-infected Chinese Oncomelania snails and the emitted cercariae. The new isolate reacted positively to the universal primers of cry1 and cry4 family genes. Each standard strain reacted positively to the primers of its characteristic family genes. Neither the three standard strains nor the new isolate responded to primers specific for the entire coding regions of 7 cry genes including cry1, cry3 and cry4. Moreover, the results of hybridization illustrated the presence of genes with homology to cry1, cry3 and cry4 family genes in the new isolate strain as well as the three reference strains. The results of SDS-PAGE displayed common protein bands at about 66, 43 and 35 kDa in the protein patterns of strains from different serovars as well as the new isolate in addition to distinct protein bands for each strain. However, characterization of the new isolate by different molecular methods indicated that this isolate is closely related to the reference strains. The spore-crystal suspensions of B. thuringiensis isolate has a molluscicidal activity on non-infected snails only.
PDF Abstract XML References
How to cite this article
Halima, H. Salem, Bahy A. Ali, Tian H. Huang and Qing D. Xie, 2006. Molecular Characterization of Novel Bacillus thuringiensis Isolate with Molluscicidal Activity Against the Intermediate Host of Schistosomes. Biotechnology, 5: 413-420.
DOI: 10.3923/biotech.2006.413.420
URL: https://scialert.net/abstract/?doi=biotech.2006.413.420
DOI: 10.3923/biotech.2006.413.420
URL: https://scialert.net/abstract/?doi=biotech.2006.413.420
INTRODUCTION
Bacillus thuringiensis is a Gram-positive bacterium that forms a parasporal crystal during the stationary phase of its growth cycle. B. thuringiensis was initially characterized as an insect pathogen and its insecticidal activity was attributed largely or completely (depending on the insect) to the parasporal crystals (Schnepf et al., 1998). More than 200 crystal protein genes (encoding Cry and Cyt proteins) have been described (Hernández et al., 2005). On the basis of the flagellar (H) antigens, B. thuringiensis has been classified in 69 serotypes comprising 82 serovars (Lecadet et al., 1999). B. thuringiensis is already a useful alternative or supplement to synthetic chemical pesticide application in commercial agriculture, forest management and mosquito control.
Schistosomiasis is a parasitic disease transmitted to man and other mammals via snails. Three major schistosome species are known to infect humans; Schistosoma haematobium, Schistosoma mansoni and Schistosoma japonicum (Ross et al., 2001). B. thuringiensis israelensis is used in localities where mosquito or blackfly larvae are present together with snail vectors of schistosomes and therefore the active metabolite could be useful in combined control of the mentioned parasite-transmitting vectors without any impact on nontarget organisms. Its usage can offer a barrier against transmission of other trematodoses, too (Horák et al., 1996). Preliminary studies (Weiser et al., 1992) have shown that a water-soluble metabolite of B. thuringiensis israelensis is toxic for aquatic snails including Biomphalaria glabrata and cercariae of flukes represented by Trichobilharzia szidati. B. thuringiensis was reported to be a good biocontrol agent against Egyptian Biomphalaria alexandrina snails (Osman et al., 1992). Toxicity of M-exotoxin of B. thuringiensis israelensis towards different cercariae of seven trematode species including Schistosoma mansoni has been reported (Horák et al., 1996). Biomphalaria glabrata snails showed high degree of sensitivity to Brevibacillus laterosporus (De Oliveira et al., 2004). Molluscicidal activity of B. thuringiensis towards Biomphalaria alexandrina (intermediate host of Schistosoma mansoni in Egypt) has been investigated (Salem, 2004). Exploration of biological factors for control of Oncomelania snails that act as intermediate host of Schistosoma japonicum in Asia is not developed. To our knowledge, no reports have described the responses of Oncomelania snails to B. thuringiensis strains. Despite many programs have controlled schistosomiasis in many areas in China but the disease is still endemic in Hunan, Hubei, Jiangxi, Anhui, Jiangsu and the mountain areas Sichuan, Yunnan provinces (Zhu et al., 2004).
In this study a new B. thuringiensis isolate (toxic to Biomphalaria alexandrina snails) has been characterized in terms of protein analysis and cry gene content. In addition, the biocidal activity of this isolate was preliminary investigated against infected, non-infected Chinese Oncomelania snails and the emitted cercariae under laboratory conditions.
MATERIALS AND METHODS
Experiments of this study were carried out in Laboratory of Cell Biology and Medical Genetics Department, Shantou University, Medical College, People’s Republic of China during 2006.
Bacterial strains and growth conditions: Four B. thuringiensis strains were used in this study. B. thuringiensis tenebrionis (tt4) and B. thuringiensis israelensis (977) were kindly supplied by Prof. Priest FG, Heriot University, England). B. thuringiensis Berliner ACCC 10061 was bought from China Center of Industrial Culture Collection, CICC. The new B. thuringiensis isolate (designated 66) was recovered from dead Biomphalaria alexandrina snails in Egypt and selected from several isolates on the basis of bioassay of its toxicity against Biomphalaria alexandrina snails, in addition to sequence of morphological, biochemical and molecular tests (Salem, 2004). The strains were maintained at -20°C as cell and spore suspensions in 20% glycerol.
Preparation of genomic DNA and polymerase chain reaction (PCR): Genomic DNA was prepared from bacterial strains according to Beard et al. (2001). Genomic DNA from the four strains (tt4, 977, Berliner ACCC 10061 and 66) was amplified using three sets of primers; 6 primer pairs specific to conserved regions of the cry1, cry3 and cry4 genes (Table 1) (Carozzi et al., 1991), 7 primer pairs specific for entire coding regions of 7 cry genes from 6 different families (Table 2) (Beard et al., 2001) and 5 pairs of universal primers were selected from a region that is highly conserved in cry1, cry2, cry3, cry4 and cry7, 8 gene families (Table 3) (Ben Dov et al., 1997). One pair of mentioned primers was used in one reaction. The 45 μL PCR reaction mix including 40 pmol of each primer was added to 5 μL of each DNA template. The amplification programme of each set of primers was performed according to Carozzi et al. (1991), Beard et al. (2001) and Ben Dov et al. (1997), respectively. An isolate was considered to contain a determined gene only when the amplification product was of the expected size.
Probe preparation: In this study six probes were amplified from the three B. thuringiensis reference strains using the primers specific to conserved regions (Table 1) as follow: 490 and 986 bp fragments from the cry1 genes were amplified from chromosomal DNA of B. thuringiensis Berliner ACCC 10061; 797 and 1290 bp fragments from the cry4 genes were amplified from chromosomal DNA of B. thuringiensis israelensis (977); and 649 and 1060 bp fragments from the cry3 genes were amplified from chromosomal DNA of B. thuringiensis tenebrionis (tt4).
| Table 1: | Nucleotide sequences of primers specific to conserved regions |
 | |
| Table 2: | Nucleotide sequences of primers specific for entire coding regions |
 | |
| Table 3: | Nucleotide sequences of universal primers |
 | |
The amplification products were labelled with random hexanucleotide primers and digoxigenin (DIG) 11-dUTP according to the manufacturer’s instructions (Roche Applied Science).
Southern blot analysis: Genomic DNA was digested with the restriction enzyme EcoRI, electrophoresed on a 1% agarose gel in TBE buffer, transferred to nylon membranes according to the method of Southern (1975) and the DNA was cross-linked to the membrane by exposure to UV-light. The membranes were prehybridized at 62°C for at least 1 h using 20 mL prehybridization buffer (5XSSC, 1% blocking reagent, 0.1% N-laurylsarcosine, 0.02% SDS) (Hansen et al., 1998) for each 100 cm membrane. Prehybridization buffer was replaced by 10 mL hybridization buffer (20 μL of freshly denatured DIG-labeled PCR product was added to 10 mL prehybridization buffer). Hybridization was performed overnight at 62°C. After reassociation, hybridized DNA was detected using an antibody reaction linked to alkaline phosphatase according to the manufacturer’s instructions (Roche Applied Science).
Dot blot analysis: Five microliter of each PCR product, destined to constitute the six probes mentioned above, was individually spotted onto super charged nylon membrane (NYTRAN). Genomic DNA from the new B. thuringiensis isolate and the strain (977) was labelled with random biotinylated octamers and biotinylated dATP using NEBlot Phototop kit (Bio Labs). Hybridization was performed at 68°C overnight in hybridization solution (6XSSC, 5XDenhardt's solution, 0.5% SDS, 100 μg mL-1 denatured salmon sperm DNA) according to standard hybridization protocols. Hybridized DNA was detected using protocols from the Phototop-Star detection kit (Biolabs).
Protein profile: Control and tested B. thuringiensis strains were grown on Petri dishes with nutrient agar and incubated at 30°C for 3-4 days. Spores plus crystals were harvested and washed in distilled water and then spore-crystal suspensions were centrifuged at 12,000 rpm for 10 min at 4°C. Soluble toxin protein was extracted from the pellets according to Gill and Hornung (1987) with some modifications as following: The pellets were solubilized in 200 μL of 0.05 M NaOH for 3 h at 37°C. Seventy five microliter preheated gel loading buffer was added and then the suspensions were boiled for 10 min. The spores were pelleted by centrifugation and the supernatants (soluble proteins) electrophoresed on 10% SDS-polyacrylamide gel according to Laemmli (1970). After electrophoretic separation, the gel was stained with Coomassie blue.
Preparation of spore-crystal suspensions: Spore-crystal suspensions of B. thuringiensis strains (66) and (977) were prepared according to Sun et al. (1996). Briefly, B. thuringiensis strains were cultured on nutrient agar 4 days at 30°C until most of the cells had lysed and crystals had been released. The spores and crystals (biomass yield) were harvested and weighed. Known weight of wet biomass was suspended into 20 mL of dechlorinated tap water. The spore-crystal suspensions were used directly as biomolluscicide.
Oncomelania hupensis snail bioassay: The intermediate host of Schistosoma japonicum was collected from Xichang city of Sichuan province in China. To detect possible infections by Schistosoma japonicum, snails were kept in 200 mL jars in tap water and under light for 3 h. The water was examined under light microscope for the existence of cercariae. The snails were transferred to container containing sterile wet soil. The container was maintained in an aquarium, with little amount of stayed (dechlorinated) tap water, at room temperature. The aquarium was covered with a stainless steel mesh. The snails were allowed to adapt to these conditions for 1-3 weeks.
Treatment involved the immersion of twenty Oncomelania hupensis snails of uniform size in 20 mL suspension of wet biomass in dechlorinated tap water (final concentration, 1-3 mg mL-1), for 72 h. And then, the snails were washed with and reimmersed in dechlorinated tap water to recover them for 24 h. Two replicates were tested for each strain. The mortality percentage was observed daily and calculated as the number of dead snails to the starting number. The sample was replaced by dechlorinated tap water in the control.
RESULTS
Identification of cry genes by universal primers: The DNA of each B. thuringiensis strain served as the template in five reactions with five pairs of universal primers (Table 3) to detect 20 cry1, 3 cry2, 4 cry3, 2 cry4, 2 cry7 and 3 cry8 genes. The results showed that the DNA of the B. thuringiensis (66) strain reacted positively to primers of cry1 and cry4 groups by producing fragments of 277 and 439 bp, respectively (Fig. 1A and B).
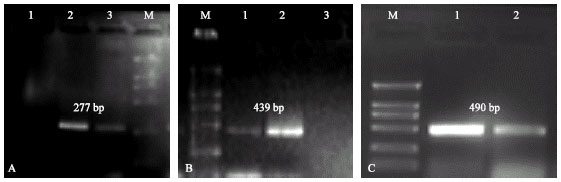 | |
| Fig. 1: | Agarose gel electrophoresis of PCR products obtained with: A) Universal primers for the cry1: lane 1, negative control (without DNA); lane 2, DNA of B. thuringiensis Berliner ACCC 10061; lane 3, B. thuringiensis isolate (66); B) Universal primers for the cry4 lane 1, B. thuringiensis isolate (66); lane 2, DNA of B. thuringiensis israelensis (977); lane 3, negative control (without DNA) and C) specific primers for the cry1: lane 1, DNA of B. thuringiensis Berliner ACCC 10061; lane 2, B. thuringiensis isolate (66). Lane M, DNA marker (DL2, 000) |
While the DNA of standard B. thuringiensis strains reacted to primer pairs as following: B. thuringiensis Berliner ACCC 10061 reacted positively to primer pairs of cry1 and cry2; B. thuringiensis israelensis (977) reacted to cry1 and cry4 group primers and B. thuringiensis tenebrionis (tt4) reacted to cry1 and cry3 primers producing the expected size (Table 3).
Identification of cry genes by specific primers: All B. thuringiensis strains responded positively to the specific primers (Table 1) and gave PCR products of characteristic size for cry1, cry3 and cry4 genes. B. thuringiensis Berliner ACCC 10061 responded positively to primer pairs of cry1 genes by producing 490 and 986-bp fragments; 797 and 1290-bp fragments from the cry4 genes were amplified from genomic DNA of strain B. thuringiensis israelensis (977) with primer pairs specific for cry4 genes; cry3 genes were identified in strain B. thuringiensis tenebrionis (tt4) by 649 and 1060-bp products. It was noticed that the new B. thuringiensis isolate was positive to specific primer of cry1 genes only (Fig. 1C). All B. thuringiensis strains did not identified by any specific primer of whole genes (Table 2).
Hybridization: Labelled genomic DNA of B. thuringiensis (66) as well as of strain 977 hybridized with six PCR amplicons derived from cry1, cry3 and cry4 family genes (Fig. 2). Southern blots of total EcoRI digested DNA from B. thuringiensis (66) and the three B. thuringiensis standard strains (ACCC 10061, 977 and tt4) hybridized with the six probes described above.
The hybridizations between the 6 probes; 2 cry1; 2 cry3; and 2 cry4 probes, respectively and the investigated strains resulted in lower intensities than the hybridization with the corresponding positive controls; ACCC 10061; tt4; and 977, respectively.
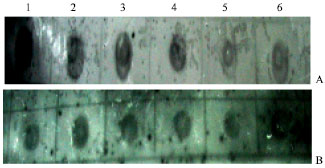 | |
| Fig. 2: | Dot-blot hybridization of cry3A (1), cry3B (2), cry4AB (3), cry4A (4), cry1Aabc (5) and cry1Ac (6) PCR product with labelled genomic DNA from B. thuringiensis strains: 977 (A) and 66 (B) |
Protein profile: Alkali-solubilized toxin protein from the isolated B. thuringiensis strain (66) was analysed by 10% SDS-PAGE and compared with that of known toxin proteins produced by three references strains (977, ACCC 10061 and tt4). The result showed that the Cry protein profiles of the four strains were variable (Fig. 3). Multiple polypeptide bands were detected in the spore/crystal alkali extract of B. thuringiensis (66). Among them, there were five major protein bands of about 66, 50, 43, 40 and 35 kDa. It also had very faint molecular weight protein of approximately 130 kDa. All strains synthesized a main protein band of about 66,43 and 35 kDa. The band of around 120 kDa appeared clear in protein profile of strain ACCC 10061 but very faint in the protein pattern of strain tt4.
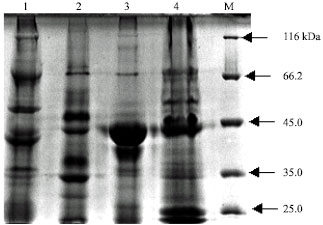 | |
| Fig. 3: | Parasporal crystal protein profiles of four B. thuringiensis strains: lane 1, ACCC 10061; lane 2, new isolate; lane 3, tt4; lane 4, 977; lane M, Protein molecular weight marker |
Effect of B. thuringiensis on infected and non-infected field collected Oncomelanea hupensis snails: In the first experiment, non-infected snails were exposed to spore-crystal suspensions of B. thuringiensis strains (66) and (977) at a concentration of 1 mg mL-1 and the mortality rates were 61.5 and 61%, respectively. With respect to the control group, 27% mortality was recorded. The second experiment was conducted with the infected snails that were tested for the existence of cercariae one week before the beginning of the experiment. The spore and crystal suspensions of B. thuringiensis strains (66) and (977) at a concentration of 3 mg mL-1 were tested against infected snails at the time of shedding cercariae. It was observed that the infected snails showed limited susceptibility to spore suspension of B. thuringiensis (66) (10% mortality) as compared with those of non-infected snails. The mortality percentage of snails treated with B. thuringiensis (977) as well as in the control group was 2.5%. However, no mortality was observed in the emitted cercariae from snails during exposure time to the spore and crystal complexes of both strains.
DISCUSSION
In the present study, novel B. thuringiensis strain that has been selected for being toxic against Biomphalaria alexandrina snails (Salem, 2004) was preliminary bioassayed for toxicity against infected, non-infected Chinese Oncomelania snails and the emitted cercariae. B. thuringiensis (977) strain belonging to serovar israelensis, which revealed toxicity for aquatic snails including Biomphalaria glabrata and cercariae of flukes represented by Trichobilharzia szidati (Weiser et al., 1992), was used for comparison. The results showed that the spore-crystal suspensions of these strains have a molluscicidal activity on non-infected snails only. The mortality percentage observed in control groups of the two experiments, may be due to changes in snail habitats (Ohmae et al., 2003), were 2.5 and 27%, respectively. It is possible that there is a correlation between the schistosome infection and the degree of susceptibility of infected snails to both B. thuringiensis strain and the changes in snail habitats. Soomro et al. (2005) reported that the increase in the number of amoebocytes during developmental stages of schistosome in vector snails Biomphalaria globrata and Bulinus natalensis especially when cercariae emerge from the host. Sasaki et al. (2005) also reported that the differences in hemocyte responses between the schistosome-susceptible and-resistant Oncomelania nosophora.
In our previous study, it has been shown that the spore and crystal suspensions of the same strains were highly toxic (80% mortality) for both non-infected field collected and laboratory-reared Biomphalaria alexandrina snails in Egypt at a concentration of 2 mg mL-1 (Salem, 2004). On the other hand, the different preparations of B. thuringiensis as Thuricide, SAN 401, 402 and 415 had lethal effect at 2000 ppm on Biomphalaria alexandrina snails after 72 h while, 250 ppm of the same preparations reduced the number of egg masses as well as the number of eggs per egg masses (Osman et al., 1992).
The cercariae bioassay result may be due to the non-susceptibility of cercariae to the tested strains or the concentration used. The high mortality in cercariae of seven trematode species was caused by M-exotoxin of B. thuringiensis israelensis eluate at high concentrations (Horák et al., 1996). The bioassay data for Oncomelania snails presented in this study encourage further experimentation to more fully evaluate the potential use of B. thuringiensis as bioinsecticide for the control of this organism.
The B. thuringiensis (66) was further characterized using molecular approaches in comparison with three B. thuringiensis standard strains known to contain the genes encoding three major classes of B. thuringiensis crystal proteins (Carozzi et al., 1991; Ben Dov et al., 1997). The PCR data showed that B. thuringiensis (66) was positive with the family primers for cry1 genes (Table 1 and 3) but negative with specific primers for cry1 genes (Table 2). In addition, the new isolate (66) was positive with universal primers of cry4 genes but not identified by any specific primer of cry4 genes (Table 1 and 2). It may contain at least 1 of the cry1 and cry4 family genes or a new cry1 and cry4 genes. DNA from the B. thuringiensis standard strains (Israelensis (977) and Tenebrionis tt4), known to contain the cry4 and cry3 genes, respectively was also amplified with cry1 universal primers. Similarly, B. thuringiensis Berliner ACCC 10061 known to contain the cry1 genes was amplified with cry1 primers (Table 1 and 3) in addition to cry2 primers (Table 3). This observation is consistent with that of Ben Dov et al. (1997) reporting that several known strains have been found to include additional combinations of genes from other groups such as, cry1, cry2 and cry7 or cry8. These authors also reported that most of the standard strains with at least one cry1 gene also contained cry2Ab. Most B. thuringiensis isolates contained several cry genes and had cry type gene combinations (Zhang et al., 2000).
Neither the three standard strains nor the new isolate that we screened responded to primers specific for the entire coding regions of 7 cry genes (Table 2). This is may be due to the difference in the source of template DNA used in this study and those used by Beard et al. (2001). These authors stated, A probe that incorporates full length gene sequences should in theory be capable of detecting a greater diversity for novel genes.
Both dot blot and Southern blot hybridization approaches resulted in detection of five additional genes in the new B. thuringiensis strain (66) with homology to the cry1, cry3 and cry4 families despite the fact that PCR had detected only a single cry gene, with homology to the cry1 family. It has been reported that PCR analysis will fail to detect novel genes with mismatches in the region of the PCR primers, even though the genes may overall possess extensive homology to a particular cry family (Beard et al., 2001; Hansen et al., 1998), which supports the use of hybridization analysis in preference to PCR. Similarly, cry1, cry3 and cry4 genes were detected in all B. thuringiensis standard strains that represent three different serovars. This observation is consistent with that of Rivera and Priest (2003), who found that 31 serovars contained at least one strain with a closely related or identical RAPD pattern to a strain from a different serovars. In the present study, we found that the most strong hybridization signals produced from positive controls from which the probes were derived. For example, two cry4 probes hybridized to B. thuringiensis israelensis (977) with strong signals while the more weakly hybridizing bands (with variable intensities) produced from the other investigated B. thuringiensis strains which may represent hybridization to other cry4 genes (Liu et al., 1996; Thanabalu and Porter, 1996). This observation may also reflect the percentage of sequence identity between the genes in the probe and in the tested strain (Beard et al., 2001).
The genes were located on 15 kb plasmid band which was observed in both non-digested and EcoRI digested genomic DNA patterns of all tested strains. B. thuringiensis has been known to have several circular/linear plasmids and cry genes are generally found in these plasmids (Carson et al., 1996). It has been found that the 15 kb plasmid band was present in 37 cry gene-positive isolates (Apaydin et al., 2005). In this study we also have found that the dot blot yielded greater signal intensity with a given quantity of PCR product (conserved regions of known genes), presumably due to the greater molar concentration of the probe used. Therefore dot blot only using of conserved regions of known genes may be more convenient for detecting diverse range of cry genes in a single step.
Common protein bands at about 66.43 and 35 kDa in the protein patterns of strains from different serovars as well as the new isolate also suggested similarities between all strains. On the other hand, distinct protein pattern for isolated strain and each serovar confirmed genetic diversity among different B. thuringiensis serovars, which previously reported (Priest et al., 1994; Burtseva et al., 1995; Rivera and Priest, 2003). The high molecular weight protein, band of around 120 or 130 kDa, appeared clear in protein profile of strain ACCC 10061 but very faint in other strains also confirmed this genetic diversity and indicated that the additional genes (at least cry1 genes) in all strains that gave positive with PCR and hybridization were unknown genes with similarity to the tested cry gene families. Previously, RAPD-PCR patterns obtained with total of seventeen random primers revealed that the average of genetic similarities between the new isolate (66) and ACCC 10061, tt4 and (977) was 42.47, 51.10 and 45.90, respectively (Salem et al., 2006).
ACKNOWLEDGMENTS
The authors wish to thank Drs. Zheng-Zhong Feng and Chang-De Li who provided us with the snails. The authors gratefully acknowledge the support of the leaders of Shantou University Medical College.
REFERENCES
- Abreu, E.F.M. and F.J.L. Aragao, 2007. Isolation and characterization of a myo-inositol-1-phosphate synthase gene from yellow passion fruit (Passiflora edulis f. flavicarpa) expressed during seed development and environmental stress. Ann. Bot., 99: 285-292.
CrossRef - Beard, C.E., C. Ranasinghe and R.J. Akhurst, 2001. Screening for novel cry genes by hybridization. Lett. Applied Microbiol., 33: 241-245.
Direct Link - Ben-Dov, E., A. Zaritsky, E. Dahan, Z. Barak and R. Sinai et al., 1997. Extended screening by PCR for seven cry-group genes from field-collected strains of Bacillus thuringiensis. Applied Environ. Microbiol., 63: 4883-4890.
Direct Link - Burtseva, L.I., V.A. Burlak, G.V. Kalmikova, H. de Barjac and M.M. Lecadet, 1995. Bacillus thuringiensis novosibirsk (Serovar H24a24c), a new subspecies from the West Siberian plain. J. Invertebrate Pathol., 66: 92-93.
CrossRefDirect Link - Carozzi, N.B., V.C. Kramer, G.W. Warren, S. Evola and M.G. Koziel, 1991. Prediction of insecticidal activity of Bacillus thuringiensis strains by polymerase chain reaction product profiles. Applied Environ. Microbiol., 57: 3057-3061.
Direct Link - Carson, C.R., T. Johenson, M.M. Lecadet and A.B. Kolsto, 1996. Genomic organization of the entomopathogenic bacterium Bacillus thuringiensis subsp. Berliner. Microbiology, 142: 1625-1634.
CrossRef - De Oliveira, E.J., L. Rabinovitch, R.G. Monnerat, L.K.J. Passos and V. Zahner, 2004. Molecular characterization of Brevibacillus laterosporus and its potential use in biological control. Applied Environ. Microbiol., 70: 6657-6664.
CrossRefDirect Link - Gill, S.S. and J.M. Hornung, 1987. Cytolytic activity of Bacillus thuringiensis proteins to insect and mammalian cell lines. J. Invertebrate Pathol., 50: 16-25.
Direct Link - Hansen, B.M., P.H. Damgaard, J. Eilenberg and J.C. Pedersen, 1998. Molecular and phenotypic characterization of Bacillus thuringiensis isolated from leaves and insects. J. Invertebrate Pathol., 71: 106-114.
CrossRef - Horak, P., J. Weiser, L. Mikes and L. Kolarova, 1996. The effect of Bacillus thuringiensis M-exotoxin on trematode cercariae. J. Invertebrate Pathol., 68: 41-49.
CrossRef - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lecadet, M.M., E. Frachon, V.C. Dumanoir, H. Ripouteau, S. Hamon, P. Laurent and I. Thiery, 1999. Updating the H-antigen classification of Bacillus thuringiensis. J. Applied Microbiol., 86: 660-672.
PubMed - Liu, J.W., A.G. Porter, B.Y. Wee and T. Thanabalu, 1996. New gene from nine Bacillus sphaericus strains encoding highly conserved 35.8-kilodalton mosquitocidal toxins. Applied Environ. Microbiol., 62: 2174-2176.
PubMed - Osman, G.Y., A.M. Mohamed and K.J. Al-Layl, 1992. Uber die molluscizide wirkung von Bacillus thuringiensis-praparaten zur biologischen bekampfuing von Biomphalaria alexandrina-schnecken, den zwischenwirten des bilharziose-saugwurms Schistosoma mansoni. Anz Schadlingskde Pflanzenschutz Umweltschutz, 65: 67-70.
- Priest, F.G., D.A. Kaji, Y.B. Rosato and V.P. Canhos, 1994. Characterization of Bacillus thuringiensis and related bacteria by ribosomal RNA gene restriction fragment length polymorphisms. Microbiology, 140: 1015-1022.
Direct Link - Gaviria, R.A.M. and F.G. Priest, 2003. Molecular typing of Bacillus thuringiensis serovars by RAPD-PCR. Syst. Applied Microbiol., 26: 254-261.
Direct Link - Ross, A.G.P., A.C. Sleigh, Y. Li, G.M. Davis and G.M. Williams et al., 2001. Schistosomiasis in the peoples republic of china prospects and challenges for the 21st century. Clin. Mirobiol. Rev., 14: 270-295.
CrossRef - Salem, H.H., T.H. Huang, B.A. Ali and Q.D. Xie, 2006. Genetic similarity among four Bacillus thuringiensis subspecies based on random amplified polymorphic DNA (RAPD). J. Boil. Sci., 6: 781-786.
CrossRefDirect Link - Sasaki, Y., M. Kirinoki and Y. Chigusa, 2005. Comparative studies of the defense mechanism against Schistosoma japonicum of schistosome-susceptible and resistant Oncomelania nosophora. Parasitol. Int., 54: 157-165.
Direct Link - Schnepf, E., N. Crickmore, J. van Rie, D. Lereclus and J. Baum et al., 1998. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev., 62: 775-806.
Direct Link - Southern, E.M., 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol., 98: 503-517.
PubMed