
A.A. Al- Khateeb
King Faisal University, Saudi-Arabia
Biotechnology
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 349-352







ABSTRACT
The effect of reducing the hormone concentration on bud and shoot multiplication of date palm (Phoenix dactylifera L.) cv. Sukry was investigated. The results indicated that low hormones promoted the formation of bud and shoot while high hormones inhibited the formation of shoots and buds. High hormones concentration resulted in abnormal growth and no sign of budding or shoot formation. The height of shoots was severely decreased by increasing the hormone concentration. Medium supplemented with 0.20 kinetin + 0.10 2ip + 0.10 BAP + 0.10 IAA + 0.10 NOA + 0.10 mg L-1NAA produced the highest fresh and dry weight.
PDF Abstract XML References
How to cite this article
A.A. Al- Khateeb, 2006. Role of Cytokinen and Auxin on the Multiplication Stage of Date Palm (Phoenix dactylifera L.) cv. Sukry. Biotechnology, 5: 349-352.
DOI: 10.3923/biotech.2006.349.352
URL: https://scialert.net/abstract/?doi=biotech.2006.349.352
DOI: 10.3923/biotech.2006.349.352
URL: https://scialert.net/abstract/?doi=biotech.2006.349.352
INTRODUCTION
Date palm is a major and dominant fruit crop in arid and semi-arid region including Kingdom of Saudi Arabia. The Kingdom is considered as one of the major countries for date palm production.
It is well known that date palm is propagated sexually through seeds and vegetatively by offshoots (Alkhateeb and Ali-Dinar, 2002). Seed propagation is not suitable because of heterozygosity and dioecious nature of the date palm which may result in producing offtype plant. Offshoot propagation is slow method and the mortality of offshoot is usually high (Alkhateeb et al., 2006). Therefore, the use of plant tissue culture technique for propagation and large scale production is very important. Normally, date palm is propagated in vitro by two methods: the first method is by embryogenesis in which vegetative embryos can continuously be formed from embryogenic callus (McCubbin et al., 2000; Letouze et al., 2000; Alkhateeb, 2005). The second procedure is organogenesis which provides date palm buds that eventually gives plantlets without passing through the callus stage. However, since plantlets are produced directly from tissues of mother plant without passing through callus stage, they are typically identical to mother plant (Beauchesne, 1983; Beauchesne et al., 1986; Aaouine, 2000). It has been reported that there are some major obstacles in practical application of organogenesis method which may limit the use of this method such as; low and irregular multiplication rate, decrease of regeneration capacity and loss of totipotency for some varieties (Zaid and de Wet, 1999; Alkhateeb, 2005).
It has been reported that the success of many in vitro techniques in higher plants depends on the success of plant regeneration (Flick et al., 1983a, b) which usually control by auxin and cytokinen concentration. In addition, it is noted that the propagation of date palm through organogenesis requires as low as possible growth substances in the media (Zaid and de Wet, 1999). Khierallah and Bader (2006) reported that sub-culturing the formed bud formation to multiplication medium supplemented with 4.0 mg L-1 2ip, 2 mg L-1 BA, 1.0 mg L-1 NAA and 1.0 mg L-1 NOA gave the optimum average of buds number (8.6 buds).
Therefore, the present research was designed to study the best auxin/cytokinen levels for bud and shoot multiplication of date palm cv. Sukry.
MATERIALS AND METHODS
This experiment was conducted in the Tissue Culture Lab. of the Date Palm Research Center, King Faisal University, Kingdom of Saudi Arabia during 2005. Date palm offshoots of cv. Khanezi approximately 3 year old and weighing 5-7 kg were separated from healthy mother trees. Offshoots were thoroughly cleaned and outer leaves were carefully removed to expose the shoot tip and lateral bud region. The exposed region was excised and immediately placed in antioxidant solution containing 150 mg-1 ascorbic acid and 100 mg-1 citric acid. The shoot tip and lateral buds were sterilized in 20% v/v Clorox solution for 15 min, followed by rinsing 3 times with distilled water. The tissues were kept in the antioxidant solution until explant excision for culturing was done.
| Table 1: | Auxins and cytokinins concentrations added to the media |
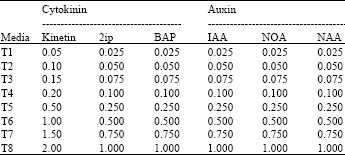 | |
The shoot tip and lateral buds were sectioned into approximately 1 cm explants which were used for organogenesis media as described by Alkhateeb and Ali-Dinar (2002). Two buds resulted from direct organogenesis were transferred to flask (250 mL) filled with 100 mL of modified Murashige and Skoog (1962) medium supplemented with 170 mg L-1 NaH2PO4.2H2O; 125 mg L-1 Inositol; 200 mg L-1 Glutamine; 1 mg L-1 Nicotinic acid; 1 mg L-1 Pyridoxine HCl; 1 mg L-1 Biotin; 1 mg L-1 Calcium Pantothenate; 7 g L-1 agar and 30 g L-1 sucrose. The media were supplemented with different auxins and cytokinins concentrations (Table 1).
Each treatment represented by 5 replicates and 2 buds per replicate in a randomized complete block design. Cultures were incubated at a 23-25°C in a 16 h photoperiod with three 40 W fluorescent lamp. Data were taken after 8 weeks of culture growth. Number of shoots (the length is 4 cm and above), buds and roots was determined. The number of cultures that produced roots was determined and expressed as percentage. Shoots and roots lengths were determined by measuring the longest shoot and root. Culture fresh weight was obtained before placing it in a forced air oven at 75°C for the determination of dry weight.
The data were statistically analyzed according to the technique of analysis of variance (ANOVA) for completely randomized design (Gomez and Gomez, 1984). The treatment means were compared using Least Significant Difference (LSD) at 5% level of probability (Waller and Duncan, 1969). All statistical analyses were performed using the facility of computer and SAS software package (SAS, 2001).
RESULTS AND DISCUSSION
Data presented in Fig. 1 and 2 showed that low hormones concentration promoted the formation of bud and shoot while high hormone concentrations were extremely inhibited the formation of shoots and buds. Medium concentration of 0.20 kinetin + 0.10 2ip + 0.10 BAP + 0.10 IAA + 0.10 NOA + 0.10 mg L-1 NAA promoted the shoot and bud formation.
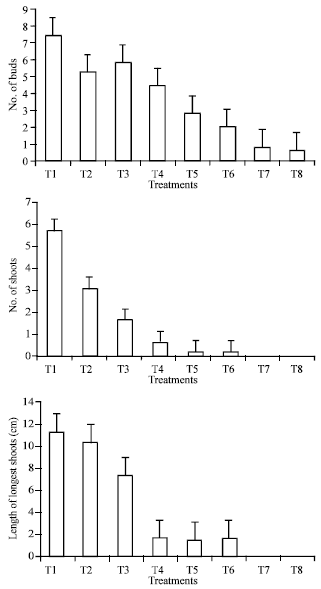 | |
| Fig. 1: | Number of buds and shoots/culture as well as length of longest shoot of date palm (Phoenix dactylifera) cv. Sukary in vitro tissue culture as affected by different concentration of cytokinin and auxin |
In addition, the quality of bud and shoot in previous mentioned medium was looking healthy compared with low hormones concentration. The shoots and buds were thicker than that of low hormones (Fig. 3). On the other hand, higher hormone concentration (T7 and T8) resulted in abnormal growth and no sign of budding or shoot formation. These results are in accordance with those obtained by Beauchesne (1983), Aaouine (2000), Zaid and de Wet (1999) and Zaid et al. (2006). They obtained a high rate of multiplication when the culture was transferred to media supplement with low levels of hormones. The results for shoot height, which presented in Fig. 1 indicated that as the concentration of hormones decreased, the height of shoots increased significantly.
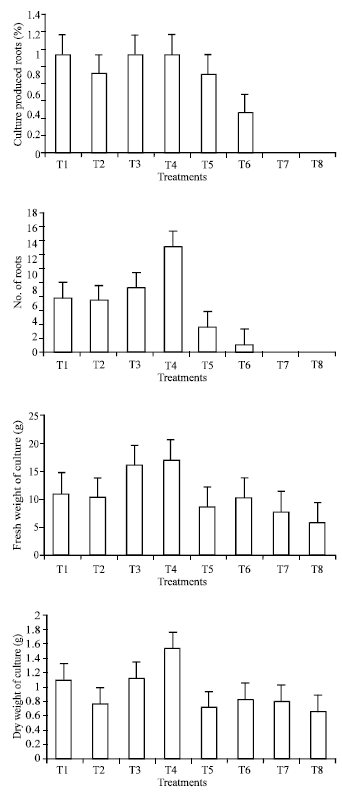 | |
| Fig. 2: | Percentage of culture produced roots, number of roots, fresh and dry weights of date palm (Phoenix dactylifera) cv. Sukary in vitro tissue culture as affected by different concentration of cytokinin and auxin |
The reduction in shoot height was more pronounced at concentration of T4-T8 compared with the other treatments which exhibited a little reduction in shoot height.
 | |
| Fig. 3: | Effect of cytokinin and auxin concentrations on bud and shoot formation of date palm (Phoenix dactylifera) cv Sukry in vitro |
Concerning the dry weight of culture, the data clearly showed that T4 had significantly the highest values. No significant effects were observed among other treatments, except treatment 1 which was exhibited slightly increase.
In this present study, the sever reduction in the bud and shoot formation of date palm at higher level of hormones may related to the failure of tissue to uptake the sugar from the medium as a result of high hormones. Therefore, the high concentration of hormone may restricted the uptake of sugar from the medium and this could cause the reduction of bud formation. The alternative explanation is the balance between auxin and cytokinin may favored the auxin. This was supported by the establishing data which showed auxin promoted callus and root formation whereas cytokinin enhanced the bud formation (Flick et al., 1983b; Alkhateeb et al., 2002).
It is noted that, in low level of hormones, the formation of healthy shoot was not yet achieved where shoots were very thin and weak (Fig. 3). Also, it is expected that in rooting stage, the formation of roots will be weak. Therefore, the chance for these plantlets to survive in acclimatization stage is highly not expected. In order to get strength and firmness for such culture it is recommended to transfer it from media 1 to 4.
These data clearly indicated the important role of hormones which can play a significant effect in influencing the rate and quality of date palm multiplication. It is clear from the results that the multiplication rate was high when the concentration of hormones was low. Buds grown on medium containing high concentration of hormones had the lowest multiplication rate.
REFERENCES
- Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Letouze, R., F. Daguin, L. Hamama, K. Paquier, F. Marionnet and J. Javouhey, 2000. Mass-propagation of date palm (Phoenix dactylifera L.) through somatoic embryogenesis. Histological study of embryo formation and cultivar identification by RAPD markers. Proceedings of the Date Palm International Symposium, 2000, Windhoek, Namibia, pp: 55-64.
- Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Waller, R.A. and D.B. Duncan, 1969. A bayes rule for the symmetric multiple comparisons problem. J. Am. Stat. Assoc., 64: 1484-1503.
CrossRefDirect Link