
S. Raveendar
Plant Biotechnology Unit, Entomology Research Institute, Loyola College,Chennai-600 034, Tamil Nadu, India
S. Ignacimuthu
Plant Biotechnology Unit, Entomology Research Institute, Loyola College,Chennai-600 034, Tamil Nadu, India
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 5 | Page No.: 256-263







ABSTRACT
Agrobacterium tumefaciens strain LBA4404 carrying binary vector pCAMBIA1305.1, with a hygromycin phosphotransferase gene (hpt) and a β-glucuronidase (GUS) gene interrupted with an intron, was used for transformation of Vigna unguiculata cotyledonary node explants. High concentration of acetosyringone (200 μM) in the Agrobacterium culture and co-cultivation medium with 1 g L-1 L-cysteine, 250 mg L-1 Na-thiosulphate and 150 mg L-1 dithiothretiol (DTT) proved to be indispensable for successful transformation. Three days old cotyledonary nodes were used for transformation studies. Binary vector pCAMBIA 1305.1 proved to be very efficient for transformation. Stable transformation with 1.61% efficiency was achieved using optimized conditions. Transformed green shoots were rooted on medium containing hygromycin. Transformed shoots tested positive for hpt gene by polymerase chain reaction. GUS activity was detected in cotyledonary nodes and leaves of the putative transformants. Southern analysis of putative transformants showed the integration of hpt into the plant genome.
PDF Abstract XML References Citation
Received: May 26, 2010;
Accepted: July 01, 2010;
Published: August 07, 2010
How to cite this article
S. Raveendar and S. Ignacimuthu, 2010. Improved Agrobacterium Mediated Transformation in Cowpea Vigna unguiculata L. Walp. Asian Journal of Plant Sciences, 9: 256-263.
DOI: 10.3923/ajps.2010.256.263
URL: https://scialert.net/abstract/?doi=ajps.2010.256.263
DOI: 10.3923/ajps.2010.256.263
URL: https://scialert.net/abstract/?doi=ajps.2010.256.263
INTRODUCTION
Cowpea (Vigna unguiculata L. Walp.) is an important food legume and a good source of protein. It is widely cultivated in the tropics and subtropics and temperate zones of the world. It occupies an area of 12.5 million ha in the world (FAO, 2000) with a total production of more than 3.3 million tones of dry grains. The average yield of cowpea is very low, due to numerous biotic and abiotic stresses. Genetic improvement will be useful for the development of cowpea with increased quality and yield (Popelka et al., 2004). However, routine genetic transformation protocols are limited in most of the tropical grain legumes, due to their inability to regenerate under tissue culture conditions (Somers et al., 2003). Generally, the family Leguminosae is highly recalcitrant due to high level of phenolics leading to oxidation of explants (Anthony et al., 1999).
Development of transformation procedure for any plant, especially for recalcitrant species involves a gene delivery method for targeting foreign DNA to regenerable cells. The most widely used method for the introduction of new genes into plants is based on the natural DNA transfer capability of Agrobacterium tumefaciens. In spite of the broad natural host range of A. tumefaciens, till today only a few grain legumes have been stably transformed using disarmed vectors (Jaiwal and Singh, 2003; Somers et al., 2003; Popelka et al., 2004).
The current trend in genetic transformation of recalcitrant grain legumes is the choice of meristem as a source of totipotent cells (Somers et al., 2003). Cotyledonary nodes are good explants for plant transformation because regeneration via direct shoot formation minimizes the risks of somaclonal variation. The optimization of some important aspects of transformation system components that affect the overall transformation efficiency is therefore, essential to enhance the virulence so as to increase the transformation frequency. In this paper, we describe a meristem targeted rapid and efficient transformation system in cowpea.
MATERIALS AND METHODS
Plant material, bacterial strain and vector: The mature seeds of V. unguiculata cultivars VBN-1, VBN-2, Co-6 and Co(cp)-7 were obtained in the year of 2006 from the Tamil Nadu Agricultural University, Coimbatore, India. Of these, the commercially grown cultivar Co(cp)-7 was used for detailed studies. The disarmed A. tumefaciens strain LBA4404, harboring a binary vector pCAMBIA1305.1, which contained a β-glucuronidase (GUS) gene and a hygromycin phosphotransferase gene (hpt) both driven by the Cauliflower mosaic virus (CaMV)35S promoter, was used for transformation (Fig. 1). The GUS gene contained an intron in its coding region to ensure that the observed GUS activity occured in the plant cell and was not due to the presence of residual Agrobacterium cells.
Preparation of explants and plant regeneration: The preparation of explants and regeneration of plants were done following the method of Raveendar et al. (2009). Healthy uniform seeds were rinsed in 70% alcohol for 1 min and then surface sterilized with 0.1% aqueous solution of HgCl2 (w/v) for 5 min. Various media used in the study are listed in Table 1. The seeds were subsequently washed several times in sterile distilled water and cultured on MSB5-G medium containing 13.3 μM BAP for 3 days at 25±2°C under dark conditions. The cotyledonary node explants (5 mm) were excised from 3-d-old in vitro raised seedlings by removing both the cotyledons and cutting both the epicotyls and hypocotyls approximately 2 mm above and below the nodal region. The explants were cultured in a vertical upright position with the hypocotyls end slightly embedded in MSB5 medium containing 6.6 μM BAP (MSB5-MB) and solidified with 0.7% agar (Hi-media,Mumbai).
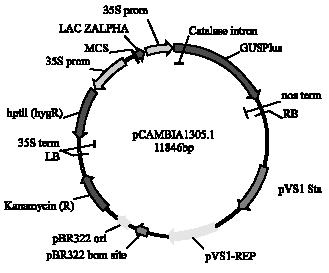 | |
| Fig. 1: | Binary vector pCAMBIA1305.1 used for transformation |
The cultures were maintained at 25±2°C under 16 h cool white fluorescent light of 50 μmol m-2 sec-1 intensity. After 2-3 weeks of multiple bud induction, the shoot buds were transferred to shoot elongation medium containing 0.5 μM BAP (MSB5-SE). After 2-3 weeks, the individual shoots were separated from the explants and transferred to MSB5 basal medium (MSB5-R) for rooting. The rooted shoots were transferred to pots containing vermiculite: soil at 1:1 ratio. Each pot was covered with a polythene bag to maintain high humidity initially for the first few days. Subsequently, the humidity was reduced by making holes in the polythene bags to harden the plants.
Transformation and plant regeneration: A. tumefaciens strain LBA4404 (pCAMBIA1305.1) was grown on AB solid medium containing 50 mg L-1 kanamycin and 10 mg L-1 rifampicin at 28°C. A single bacterial colony was inoculated into 3 mL of liquid AB medium containing the same antibiotics and grown overnight on a rotary shaker at 180 rpm at 28°C. Three milliliter aliquot of bacterial suspension was added to 30 mL of AB liquid medium containing 50 mg L-1 kanamycin and 10 mg L-1 rifampicin and grown overnight. Bacteria were pelleted at 6000 rpm for 10 min and resuspended in liquid MSB5 medium (MSB5-AS) containing 6.6 μM BAP and 200 μM acetosyringone (AS) at a density of 1OD at 600nm. The cotyledonary node explants excised from 3-d-old seedlings were immersed in bacterial suspension for 25-30 min with occasional shaking. The Agrobacterium treated explants were then blotted on sterile filter paper and transferred to coculture medium (MSB5-CCM) containing 0.8% agar and supplemented with 1 m L-1 L-cysteine, 250 mg L-1 Na-thiosulphate and 150 mg L-1 dithiothretiol (DTT) for 3 days under 16 h photoperiod at 25±2°C. After cocultivation, the explants were washed three to four times with sterile distilled water and blotted dry on sterile filter paper. The explants were cultured on MSB5 selection medium (MSB5-MBH) containing 6.6 μM BAP, 30 mg L-1 hygromycin and 500 mg L-1 cefotaxime for shoot regeneration. The explants were transferred onto shoot elongation medium (MSB5-SEH) containing 0.5 μM BAP and the same levels of antibiotics after every 2 weeks for a total of 4-6 weeks, until the shoots attained a height of 2-3 cm.
| Table 1: | Media used in the study |
 | |
The shoots were rooted on MSB5 basal medium (MSB5-RH) containing 5 mg L-1 hygromycin. The putative transformed plants were established in soil and transferred to greenhouse.
Histochemical assay for the GUS gene: The histochemical assay of GUS gene expression (β-D-Glucuronidase) was performed in cotyledonary node and leaf explants with 5-Bromo-4-Chloro-3-Indolyl Glucuronide (X-Gluc) as a substrate using the established method (Jefferson, 1987). The putative transformed explants were incubated in sodium phosphate buffer (50 mM NaPO4, pH 6.8) that contained 1% Triton X-100 at 37°C for 1 h and were later incubated overnight in a solution containing 1.0 mM X-Gluc, 10 mM ETDA, 100 Mm NaH2PO4, 0.1x TritonX-100 and 50% methanol (pH 5.8). The tissue was washed twice in 99% methanol for 2 h to remove chlorophyll pigment. The number of tissue that stained blue was counted as GUS positive.
PCR analysis of putative transgenic plants: Total genomic DNA was extracted from fresh leaves of putative transformed (T0) and non-transformed (control) plants by the CTAB (cetyl trimethyl ammonium bromide) method (Doyle and Doyle, 1987). PCR analysis was carried out using the following primers (5’ primer, 5’ AAAGCCTGAACTCACCGC 3’ 3’primer, 5’ GCTTTCCACTATCGGCGA 3’). These primers amplified a 1.1kp fragment from the hpt gene. PCR analysis was carried out in a reaction volume of 25 μL containing the template genomic DNA (100 ng), 2.5 μL 10X PCR amplification buffer, 0.5 μL 10 mM dNTPS, 1.2 μL 50 mM MgCl2, 3 μM (2.5 μL) of each primer, 13.6 μL sterile distilled water, 1 unit (0.20 μL of Taq DNA polymerase (Genei). The samples were heated to 94°C for 5 min and then subjected to 30 cycles of 30 sec melting at 94°C, 30 sec annealing at 60°C and 1 min synthesis at 72°C and followed by another 10 min final extension at 72°C. The amplified products were assayed by electrophoresis on 0.8% agarose gels, stained with ethidium bromide (EtBr; 0.5 μg mL-1), visualized and photographed under ultraviolet light.
Southern blot analysis: The T0 tranformants were subjected to southern blot hybridization using the coding sequence of hpt gene. Southern blot analysis was done by using PCR samples of putative transgenics. The PCR samples were resolved on 0.8% agarose gels. DNA was transferred to positively charged nylon membrane (as per the manufacturer’s instructions) for southern hybridization (Southern, 1975). The coding sequences of hpt genes were labelled with biotin -11-dUTP using biotin decalabel DNA labeling kits (Fermentas Life sciences) and used as a probe. The blot was subjected to detection by overnight colour development using biotin chromogenic detection kit (Fermentas Life sciences).
RESULTS
Sensitivity of explants to hygromycin and cefotaxime: Prior to transformation, an effective concentration of antibiotic for the selection of transformants was determined by culturing cotyledonary node explants on MSB5-MB and MSB5-SE media, respectively, containing various concentrations of hygromycin (0, 10, 20, 30, 40 and 50 mg L-1). Hygromycin at a concentration of 30 mg L-1 caused almost total inhibition of shoot initiation on cotyledonary node explants (Table 2); hence this concentration was used for selection of transformed shoots. All the cotyledonary node explants produced shoots on hygromycin-free MSB5-MB medium. Influence of cefotaxime on shoot initiation and on subsequent growth was also checked by culturing explants on MSB5-MB and MSB5-SE medium containing 500 mg L-1 cefotaxime. This concentration had no effect on shoot initiation and subsequent growth, but effectively controlled the Agrobacterium growth.
Transformation: Inoculation of explants in the Agrobacterium culture for 25-30 min was done as longer periods of incubation posed problems in the elimination of bacteria and contamination in subsequent cultures of cowpea explants in vitro. After inoculation of explants for 25-30 min the explants were transferred to MSB5-CCM medium and incubated for cocultivation for 3 days. At the end of the cocultivation period of 3 days, the explants were transferred to selection medium (MSB5-MBH) and cultured for 2 weeks. After 2 weeks of selection, the fresh survived explants were transferred to MSB5-SEH medium for second round of selection that selected only those cells receiving and expressing the gene transfer casette.
| Table 2: | Effect of different concentrations of hygromycin on shoot regeneration from 3-day-old cotyledonary node explants of cowpea cv. Co(cp)-7 |
 | |
| *Data represent Mean±SD of six replicates. Means followed by the same letters within a column do not differ significantly according to Duncan's multiple range test at a 5% probability level. Data were scored after 6 weeks of culture | |
A few explants (25 out of 305 explants treated with Agrobacterium strain with pCAMBIA1305.1) survived and formed shoots (Fig. 2a, b) on MSB5-SEH medium containing (30 mg L-1) hygromycin after 4-5 weeks of culture. The shoots did not grow and elongate further after 6 weeks of culture on MSB5-SEH medium supplemented with 30 mg L-1 hygromycin. The untreated explants cultured on MSB5-SEH medium containing 30 mg L-1 hygromycin did not survive. Following cocultivation with Agrobacterium, the regenerating explants were placed for selection medium (MSB5-SH) containing 30 mg hygromycin for 2 week followed by 2-3 weeks in MSB5-SEH medium. A total of 305 explants, in three different experiments, produced 25 shoots on hygromycin selection medium (Table 3). After 6 weeks, the elongated shoots were transferred to rooting medium containing lower concentration (5 mg L-1) of hygromycin, where only 16 shoots (64%) produced roots.
GUS analysis: All the cotyledonary node explants showed GUS activity after cocultivation with A. tumefaciens LBA4404/pCAMBIA1305.1 harboring GUS gene, predominantly in the regenerable sites (Fig. 2). While endogenous gues expression was not detected in the tissues of control plants, a strong and stable gus expression was observed in cotyledonary node and leaf of all the hygromycin resistant putative transformants.
| Table 3: | Summary of the transformation of 3-day-old cotyledonary node explants of Vigna unguiculata cv. Co(cp)-7 co-cultivated with Agrobacterium tumefaciens strain LBA4404 harboring a binary vector pCAMBIA1305.1 |
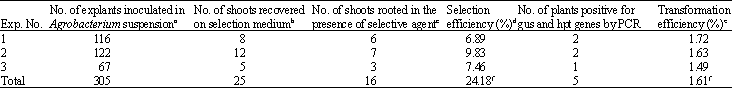 | |
| aExplants cocultured with Agrobacterium for 3 days. bExplants produced shoots on selection medium: MSB5-SE + hygromycin (30 mg L-1) and cefotaxime (500 mg L-1) for 4-6 weeks of culture. cShoots rooted on MSB5-R + hygromycin (5 mg L-1) and (500 mg L-1)cefotaxime. dSelection efficiency: percentage of hygromycin-resistant plants. eTransformation efficiency: percentage of initial explants that developed to plants positive for gus and hpt genes by PCR. fAverage | |
 | |
| Fig. 2: | Agrobacterium-mediated transformation of Vigna unguiculata cv. Co(cp)-7. (a) Explants inoculated with Agrobacterium tumefaciens strain LBA4404 (pCAMBIA1305.1) and cultured on MSB5-MBH medium containing 30 mg L-1 hygromycin showing regeneration of green shoots. (b) Direct shoot regeneration from cotyledonary node explants on MSB5-SEH medium supplemented with 0.5 μM BAP after 4-5 weeks of culture. (c) Induction of roots from in vitro regenerated shoots. (d) A fertile transgenic plant growing in pot. (e) Cotyledonary node explant showing transient GUS activity after 3 days of cocultivation with Agrobacterium tumefaciens LBA4404 (pCAMBIA1305.1). (f) Stable GUS activity in leaf of putative tyransformants |
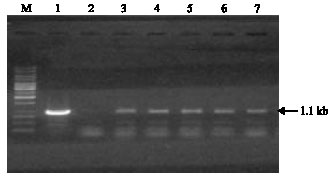 | |
| Fig. 3: | PCR analysis of primary transformants using the hpt primers. Lane M, Marker DNA; lane 1: +ve, plasmid DNA; lane 2: DNA from untransformed control; lanes 3-7: T0 transformed plants |
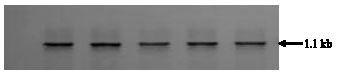 | |
| Fig. 4: | Southern blot analysis of putative transformants |
We determined the stable transformation efficiency based on the plants positive for Gus and hpt genes by PCR. A total of five independently derived transgenic plants were obtained from a total of 305 explants, giving an average transformation frequency of 1.61%.
Molecular analysis of putative transgenics: PCR amplification was performed using hpt gene primers; five plants out of 16 plants showed PCR amplification (Fig. 3). Further these PCR samples were tested for confirmation in Southern analysis; five samples, which showed clear amplification in PCR, were Southern positive (Fig. 4). Southern hybridization was carried out with genomic DNA from GUS expressing, hygromycin resistant shoots. The hpt gene sequence was detected as a fragment of expected size (1.1 kb) in transformed plants. Since hpt probe hybridized to genomic DNA from transgenic plants but not to DNA from non-transformed control plants, the result indicated that hpt DNA was incorporated into cowpea genome. PCR amplified plant DNA sequence was subsequently probed with hpt coding region to identify fragments between the T-DNA border. An internal 1.1 kb fragment corresponding to hpt gene was detected in transgenic plants by hpt probe (Fig. 4). The bands from genomic DNA, identified with hpt probe, contained sequences derived from plant DNA. The size of the bands (1.1 kb) confirmed the integration of the hpt gene into plant nuclear genome.
DISCUSSION
Reports on stable Agrobacterium-mediated transformation confirmed by southern analysis of primary transformants are available (Muthukumar et al., 1996; Kononowicz et al., 1997; Monti et al., 1997). Muthukumar et al. (1996) used cotyledons excised from 3 day old seedlings. The cotyledons were cocultivated with A. tumefaciens and transformed tissues selected on 25 mg L-1 hygromycin. Our preliminary work on the effect of hygromycin on in vitro regeneration of untransformed cowpea has established significant inhibition levels at 30 mg L-1 hygromycin (Table 2) which is similar to the earlier report of Obembe et al. (2000). Muthukumar et al. (1996) reported that 15-19% of explants produced shoots on hygromycin selection medium.
Our protocol for Agrobacterium-mediated transformation of cowpea is summarized in Fig. 5. The 3 days old cotyledonary node explants were inoculated with an Agrobacterium culture. After 3 days of cocultivation, the explants were transferred to selection and regeneration media. After 4-6 weeks, regenerated shoots were transferred to rooting medium. The resulting rooted shoots were subjected to screening for transformants by PCR. The positive plants were then potted into soil. A major advantage of our cowpea transformation method is that regenerated transformants can be obtained within 7 weeks after Agrobacterium inoculation.
The difference in transformation efficiency in different studies may be attributed to the genotype of the crop species, explant type, cocultivation procedure followed and the Agrobacterium strains used in the genetic transformation experiments. Solleti et al. (2008) reported transgenic cowpea using cotyledonary node as explants and A. tumefaciens LBA4404 harboring plasmid pSB1, while Chaudhury et al. (2007) used Agrobacterium strain EHA105. Similarly variation in transformation frequency was observed in cowpea using different strains of Agrobacterium and genotypes (Kononowicz et al., 1997; Monti et al., 1997). In the present study, transformation efficiency was measured as number of confirmed transgenic plants out of total number explants used for transformation experiment. In the present study cowpea variety Co(cp)-7 was transformed with Agrobacterium strain LBA4404 carrying the binary vector pCAMBIA1305.1 and the percentage of confirmed transgenic plants appeared to be similar when compared to the previous reports of cowpea (1.64%) reported by Solleti et al. (2008).
 | |
| Fig. 5: | Steps of transformation protocols for cowpea |
The T-DNA transfer is mediated by products encoded by the vir (virulence) region of the Ti-plasmid, which are activated by signal molecules, mainly small phenolics, certain class of monosaccharides and acidic pH acting synergistically with phenolic compounds (De la Riva et al., 1998). Addition of 200 μM acetosyringone to the bacterial resuspension medium as well as cocultivation medium resulted in significant increase in transformation frequency from 7.6% in cultures without acetosyringone to 82.6% with acetosyringone with large GUS positive sector(s). Acetosyringone enhanced vir functions during transformation (Stachel et al., 1986) and has been found to increase transformation potential of Agrobacterium strain with moderately virulent vir region in several plant species (Atkinson and Gardner, 1991; Janssen and Gardner, 1993; Kaneyoshi et al., 1994).
Cocultivation of explants in the presence of mixtures of thiol compounds, DTT and L-cysteine increased the frequency of Agrobacterium-mediated transformation and recovery of transgenic plants at a high frequency. Cocultivation in presence of thiol compounds has been described to be beneficial in recovering transgenic plants in soybean (Olhoft et al., 2001). Moreover, cocultivation of explants in the presence of mixtures of the thiol compounds combined with geneticin selection resulted in enhanced recovery of transgenic plants to an average of 1.67%. Transformation efficiencies for thiol compounds treated cotyledonary node explants using PPT selection in cowpea were 0.001-0.003% (Popelka et al., 2006). Employing extra copies of vir genes and geneticin-based selection and recovery of transgenics in cowpea were also reported. (Chaudhury et al., 2007). In the present study, high concentrations of acetosyringone (200 μM) in the Agrobacterium culture and cocultivation medium with 1 mg L-1 L-cysteine, 250 mg L-1 Na-thiosulphate and 150 mg L-1 dithiothretiol (DTT) proved to be indispensable for successful transformation (Fig. 6, 7).
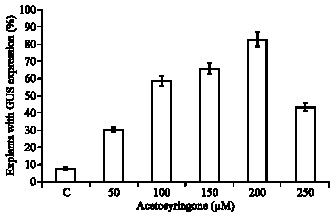 | |
| Fig. 6: | Effects of Acetosyrigone on transformation of 3-day-old cotyledonary node explants of Vigna unguiculata cv. Co(cp)-7 co-cultivated with Agrobacterium tumefaciens strain LBA4404 harboring a binary vector pCAMBIA1305.1 |
Recovery of fertile transgenic plants in our case took approximately 2 months which was significantly less than previous reports in cowpea (Sahoo et al., 2000; Popelka et al., 2006; Chaudhury et al., 2007).
Confirmation of the transgenic nature of the plants was done using PCR amplification and Southern hybridization. When probed with the 1.1 kb PCR amplified hpt gene fragment as a probe, the expected fragment of 1.1 kb was detected in the Southern analysis (Fig. 4). In the T0 generation plants, the plant genomic DNA fragments after PCR amplification that hybridized to the non-radioactive hpt probe were longer than 1.1 kb proving that the T-DNA got stably integrated into the cowpea plant genome.
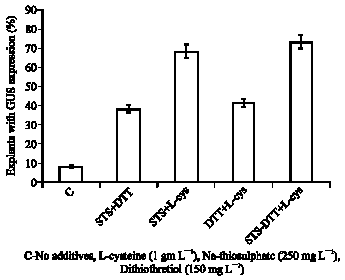 | |
| Fig. 7: | Synergistic effects of thiol compounds on transformation of 3-day-old cotyledonary node explants of Vigna unguiculata cv. Co(cp)-7 co-cultivated with Agrobacterium tumefaciens strain LBA4404 harboring a binary vector pCAMBIA1305.1 |
Our method is efficient for the recovery of stable transgenics and introduction of desirable agronomic traits into cowpea.
REFERENCES
- Chaudhury, D., S. Madanpotra, R. Jaiwal, R. Saini, P.A. Kumar and P.K. Jaiwal, 2007. Agrobacterium tumefaciens-mediated high frequency genetic transformation of an Indian cowpea (Vigna unguiculata L. Walp.) cultivar and transmission of transgenes into progeny. Plant Sci., 172: 692-700.
CrossRef - De la Riva, G.A., J. Gonzalez-Cabrera, R. Vazquez-Padron and C. Ayra-Pardo, 1998. Agrobacterium tumefaciens: A natural tool for plant transformation. Elect. J. Biotechnol., 1: 118-133.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Jefferson, R.A., 1987. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep., 5: 387-405.
CrossRefDirect Link - Kaneyoshi, J., S. Kobayashi, Y. Nakamura, N. Shigemoto and Y. Doi, 1994. A simple and efficient gene transfer system of trifoliate orange. Plant Cell Rep., 13: 541-545.
CrossRef - Kononowicz, A.K., K.T. Cheah, M.L. Narasimha, L.L. Murdock and R.E. Shade et al., 1997. Developing a Transformation System for Cowpea (Vigna unguiculata L. Walp.). In: Advances in Cowpea Research, Singh, B.B., D.R. Mohan-Raj, K.E. Dashiell and L.E.N. Jackai (Eds.). IITA, Ibadan, Nigeria, pp: 361-371.
- Olhoft, P.M., K. Lin, J. Galbraith, N.C. Nielsen and D.A. Somers, 2001. The role of thiol compounds in increasing Agrobacterium-mediated transformation of soybean cotyledonary-node cells. Plant Cell Rep., 20: 731-737.
Direct Link - Popelka, J.C., S. Gollasch, A. Moore, L. Molvig and T.J.V. Higgins, 2006. Genetic transformation of cowpea (Vigna unguiculata L.) and stable transmission of the transgenes to progeny. Plant Cell Rep., 25: 304-312.
Direct Link - Raveendar, S., A. Premkumar, S. Sasikumar, S. Ignacimuthu and P. Agastian, 2009. Development of a rapid, highly efficient system of organogenesis in cowpea Vigna unguiculata (L.) Walp. South Afr. J. Bot., 75: 17-21.
CrossRef - Solleti, S.K., S. Bakshi and L. Sahoo, 2008. Additional virulence genes in conjunction with efficient selection scheme, and compatible culture regime enhance recovery of stable transgenic plants in cowpea via Agrobacterium tumefaciens-mediated transformation. J. Biotechnol., 135: 97-104.
PubMed - Somers, D.A., D.A. Samac and P.M. Olhoft, 2003. Recent advances in legume transformation. Plant Physiol., 131: 892-899.
Direct Link - Southern, E.M., 1975. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol., 98: 503-517.
PubMed - Stachel, S.E., E.W. Nester and P.C. Zambryski, 1986. A plant cell factor induces Agrobacterium tumefaciens vir gene expression. Proc. Natl. Acad. Sci. USA., 83: 379-383.
Direct Link