
S. Nwodo Chinedu
Department of Biological Sciences, College of Science and Technology, Covenant University,KM 10 Idiroko Road, CanaanLand, PMB 1023 Ota, Ogun State, Nigeria
Angela O. Eni
Department of Biological Sciences, College of Science and Technology, Covenant University,KM 10 Idiroko Road, CanaanLand, PMB 1023 Ota, Ogun State, Nigeria
Adebayo I. Adeniyi
Department of Biological Sciences, College of Science and Technology, Covenant University,KM 10 Idiroko Road, CanaanLand, PMB 1023 Ota, Ogun State, Nigeria
Janet A. Ayangbemi
Department of Biological Sciences, College of Science and Technology, Covenant University,KM 10 Idiroko Road, CanaanLand, PMB 1023 Ota, Ogun State, Nigeria
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 3 | Page No.: 118-125







ABSTRACT
The aim of the study was to isolate and identify filamentous microfungi involved in wood-waste decomposition in Canaanland, Ota, South-West Nigeria and to evaluate their potentials for cellulose saccharification. Microbiological techniques were used to isolate and identify the fungi. Four filamentous microfungi, identified as Aspergillus niger, Aspergillus flavus, Penicillium chrysogenum and Trichoderma sp., were isolated. All the isolates, particularly Trichoderma sp., grew rapidly on Sabouraud’s agar and Czapek-Dox agar. Two of the isolates, Aspergillus niger and Trichoderma sp., was cultivated for 168 h by submerged fermentation in modified Czapek-Dox liquid medium containing cellulose as sole carbon source and harvested at 24 h intervals. The mycelia weight of the harvested cultures, and the protein content and cellulase activity of the filtrates were determined. The peak mycelia weight of 4.6 and 3.0 mg mL-1 was, respectively obtained for Trichoderma sp. and A. niger at 48 h. The protein and cellulase activity of Trichoderma sp. peaked at 72 h whereas for A. niger, the peak protein content and peak cellulase activity was obtained at 96 and 72 h, respectively. The peak protein and cellulase activity values of A. niger were 0.175 and 0.077 unit mL-1, respectively. Trichoderma sp. yielded a protein peak of 0.180 mg mL-1 and peak cellulase activity of 0.108 unit mL-1. There is a correlation between the protein content and cellulase activity of the culture filtrates. The strains of A. niger and Trichoderma sp. obtained from this study are potential tools for the saccharification and bioconversion of cellulosic materials.
PDF Abstract XML References Citation
How to cite this article
S. Nwodo Chinedu, Angela O. Eni, Adebayo I. Adeniyi and Janet A. Ayangbemi, 2010. Assessment of Growth and Cellulase Production of Wild-Type Microfungi Isolated from Ota, Nigeria. Asian Journal of Plant Sciences, 9: 118-125.
DOI: 10.3923/ajps.2010.118.125
URL: https://scialert.net/abstract/?doi=ajps.2010.118.125
DOI: 10.3923/ajps.2010.118.125
URL: https://scialert.net/abstract/?doi=ajps.2010.118.125
INTRODUCTION
Cellulose, a major component of all vegetation, is one of the world's most abundant resources (Lynd et al., 2002). It is constantly replenished by photosynthesis and accounts for nearly one-half of the 18 to 20 metric tons of organic carbon annually fixed by photosynthesis (Romanelli et al., 1975). Cellulose is also a major component of solid waste daily produced in large quantities from such activities as food processing, lumbering operations, paper making, cereal grain harvesting, sugar cane processing, domestic and office activities (Abu et al., 2000). Attempts at utilizing cellulose as a commercial fermentation substrate have been unsatisfactory mainly due to the low rate of hydrolysis by mesophilic microorganisms (Romanelli et al., 1975).Cellulase is a generic name for the group of enzymes which catalyze the hydrolysis of cellulose and cellooligosaccharide derivatives (Chinedu et al., 2008). The enzymes play important role in natural biodegradation processes in which plant lignocellulosic materials are degraded by a wide variety of fungi, bacteria, actinomycetes and protozoa (Peciulyte, 2007). They are widely used for the extraction of valuable compounds from plant cells, improving nutritional values of animal feed and in preparing plant protoplasts for genetic research (Abdul et al., 1999). They are also applied in the chemical industry for the production of fermentable sugars, ethanol, organic acids, detergents and other chemicals (Howard et al., 2003). Cellulases also find important application in the pulp and paper industry, textile industry, animal feed and food industry and also in processing cellophane and rayon (Peciulyte, 2007). Cellulase is potentially useful for industrial saccharification of cellulosic biomass (Chinedu et al., 2008). The growing interest in the conversion of lignocelluloses into bulk chemicals and biofuels as a means of alleviating energy shortages has heightened the search for highly active, thermostable and specific cellulases (Howard et al., 2003; Chinedu et al., 2008).
Fungi produce extracellular cellulase enzymes that break down cellulose into two or three glucose units which are readily degraded and assimilated as glucose monomers (Willey et al., 2008). Fungal cellulases are extracellular and inducible (Abdul et al., 1999) and consist of three components namely exoglucanase, endoglucanase and β-glucosidase (Khan, 1980). The ability of filamentous fungi to secrete cellulases and other cell wall hydrolyzing enzymes into their culture media has led to the harvesting of these useful enzymes (Berry and Patterson, 1990; Chinedu et al., 2008). In this study, the growth, protein and cellulase production of some microfungi isolated from wood-waste dump in Covenant University, Canaanland, Ota, Nigeria were evaluated. The potentials of two of the isolates for cellulose saccharification are indicated.
MATERIALS AND METHODS
Sample collection and preparation:
Sample of decomposing wood-waste was collected in a sterile bag from a construction site at Covenant University, Ota, Ogun State in March 2, 2009 and stored at room temperature. One gram of the sample was aseptically introduced into 9.0 mL of sterile distilled water in a sterile test-tube and shaken vigorously. The stock culture was used for fungal isolation.
Media preparation:
Sabouraud’s Agar (Merck, Germany) was prepared according to manufacturer’s instruction.
Czapek-dox agar:
One liter of Czapek-Dox agar consists of: 30.0 g Sucrose, 2.0 g NaNO3, 1.0 g KH2PO4, 0.5 g KCl, 0.5 g MgSO4.7H2O and 0.01 g FeSO4.7H2O and 20.0 g Agar (Oxoid, England) and 1.0 mL of trace solution (containing 1.0 g ZnSO4 and 0.5 g CuSO4. 5H2O L-1).
Modified czapek-dox agar had the same composition as Czapek-Dox Agar except that sucrose was replaced with equal quantity of crystalline cellulose.
Modified Czapek-Dox liquid medium had the same composition as Modified Czapek-Dox Agar except that it did not contain agar. Each medium was supplemented with 0.05% (w/v) Ampicillin to prevent bacterial growth. The pH was adjusted to 5.4 before sterilization by autoclaving at 121°C for 15 min.
Isolation of the fungal isolates:
One milliliter of the stock culture was inoculated on freshly prepared modified Czapek-Dox agar and incubated at 25°C for 120-168 h. Pure fungal isolates were obtained through careful isolation and successive sub-culturing of the growing fungal colony on fresh Sabouraud’s agar.
Identification of the fungal isolates:
Each isolate was identified based on its colonial and cultural properties, and the microscopic features of its sporulating structures (Harrigan and McCance, 1966). The characteristics of the vegetative mycelia (hyphae) and color of the colonies were observed over a period of 24 to 168 h. The morphology of the isolates, stained with lactophenol-cotton blue, was studied using a light microscope and compared according to the description given by Thom and Raper (1945), Alexopoulos (1952), Talbot (1972) and Deacon (1980).
Growth studies:
Colony diameter: A flamed, sterile cork borer (Pore size; 5.0 mm diameter), was used to bore a hole enclosing a disc of the pure culture of the respective pure fungus on Czapek-Dox agar plate. One disc of each fungus was aseptically transferred and placed at the center of sterile, freshly prepared Sabouraud’s and Czapek-Dox agar plates. The average diameter of the growing colony was measured at intervals of 24 for 120 h using a calibrated transparent ruler.
Mycelia weight:
Ten milliliter of 1.0% Tween 80 was placed on top of 3 day Czapek-Dox agar plate containing the pure isolate of fungus. Conidia were harvested using sterile cotton swab and transferred into a sterile test tube and diluted with equal volume of sterile distilled water. Two milliliter of the spore suspension was inoculated into 100 mL of sterile modified Czapek-Dox liquid medium in 250 mL Erlenmeyer flask. The flask was covered with sterile cotton wool and incubated in the dark at 25°C with continuous agitation at 100 rpm using the Griffin flask shaker. Cultures were harvested at 24 h intervals by filtration over a 168 h period. The mycelium was washed and dried in the oven at 80°C for 2 h, and carefully weighed. The cell-free filtrates were used for protein and enzyme analyses.
Protein assay:
Protein content of the culture filtrates was determined by the folin ciocalteau method described by Lowry et al. (1951) using Bovine Serum Albumin (BSA) as standard.
Enzyme assay:
A modified reducing sugar method of Khan (1980) as described by Chinedu et al. (2008) was used to assay for cellulase (EC 3. 2. 1. 4) activity. Carboxymethyl-cellulose (CMC) was used as enzyme substrate. The released reducing sugar was measured using 3, 5-dinitrosalicylic acid and read at 540 nm using a spectrophotometer (Miller, 1959). A unit of activity was defined as amount of enzyme required to liberate 1 μmol of Glucose per minute under the assay conditions.
RESULTS
Isolation and identification:
Four different filamentous microfungi were isolated from the wood-waste and identified as Aspergillus niger, Aspergillus flavus, Penicillium chrysogenum and Trichoderma species based on the colonial and cultural characteristics and microscopic features of the sporulating structures of the fungal isolates (Table 1). Colony of the fungal isolate growing on Sabouraud’s agar plate and the microscopic features of the sporulating structure are shown in Fig. 1a,b-4a,b.
Growth studies
Growth on sabouraud’s and czapek-dox agar:
Figure 5 shows the growth of the fungal isolates on Sabouraud’s agar. The growth on Czapek-Dox Agar is shown in Fig. 6. All the organisms grew rapidly on both media, filling the entire plate (8.5 cm diameter) between 72 to 120 h of incubation. The fastest growth on the Sabouraud’s and Czapek-Dox Agar was obtained for Trichoderma sp.
| Table 1: | Colonial and microscopic characteristics of the fungal isolates |
 | |
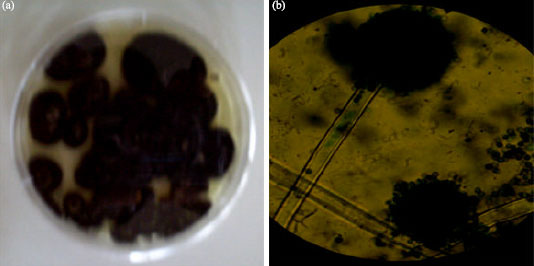 | |
| Fig. 1: | Aspergillus niger: (a) 5-day culture on Sabouraud’s agar plate and (b) Microscopic picture (x100) of the sporulating structure |
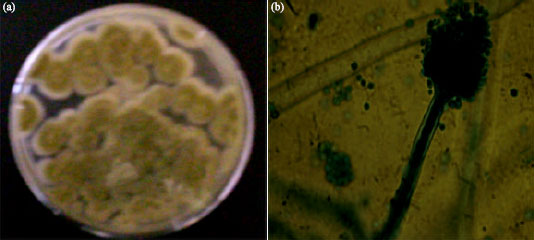 | |
| Fig. 2: | Aspergillus flavus: (a) 5-day culture on Sabouraud’s agar plate and (b) Microscopic picture (x100) of the sporulating structure |
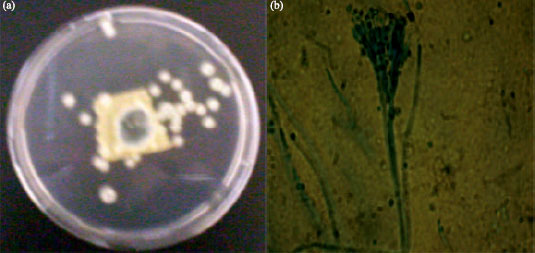 | |
| Fig. 3: | Penicillium chrysogenum: (a) 3-day culture on Sabouraud’s agar plate and (b) Microscopic picture (x100) of the sporulating structure |
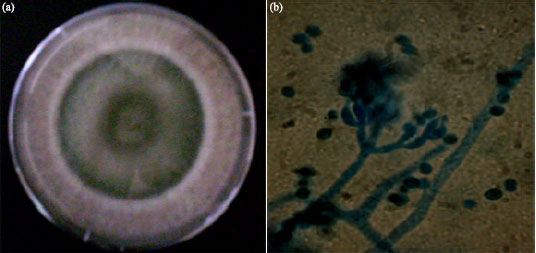 | |
| Fig. 4: | Trichoderma sp.: (a) 3-day culture on Sabouraud’s agar plate and (b) Microscopic picture (x100) of the sporulating structure |
Growth on modified czapek-dox liquid medium:
Figure 7 shows the mycelia weights of Aspergillus niger and Trichoderma sp. in modified Czapek-Dox liquid medium. The peak mycelium weight of both organisms was obtained at 48 h. Thereafter, there was a steady decline in the mycelia weight. A peak mycelia weight of 3.0 and 4.6 mg mL-1 was obtained for A. niger whereas Trichoderma sp. respectively.
Cellulase activity
Protein yield and cellulase activity:
Protein content and cellulase activity of culture filtrates of A. niger incubated in modified Czapek-Dox liquid medium is shown in Fig. 8. The peak protein content and cellulase activity of A. niger was obtained at 96 and 72 h, respectively. Figure 9 shows the protein content and cellulase activity of culture filtrates of Trichoderma sp. incubated in modified Czapek-Dox liquid medium. Both the protein and cellulase activity of Trichoderma sp. peaked at 72 h. The peak protein and cellulase activity values of A. niger were 0.175 and 0.077 unit mL-1, respectively. The cellulase activity of Trichoderma sp. was significantly higher than that of A. niger. Trichoderma sp. yielded a protein peak of 0.180 mg mL-1 and peak cellulase activity of 0.108 unit mL-1.
Mycelia weight and cellulase activity:
The mycelia weights and cellulase activity of the culture filtrates of A. niger and Trichoderma sp. harvested at 24 h intervals over a period of 168 h are shown in Fig. 10 and 11, respectively.
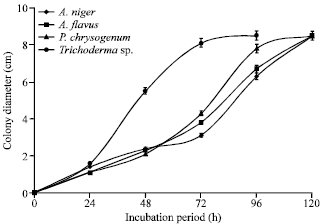 | |
| Fig. 5: | Colony diameter of fungal isolates incubated at 25°C on Sabouraud’s agar plates |
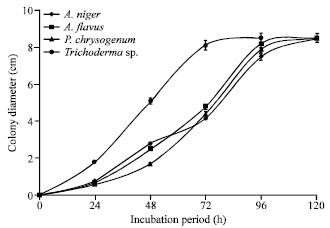 | |
| Fig. 6: | Colony diameter of fungal isolates incubated at 25°C on Czapek-Dox agar plates |
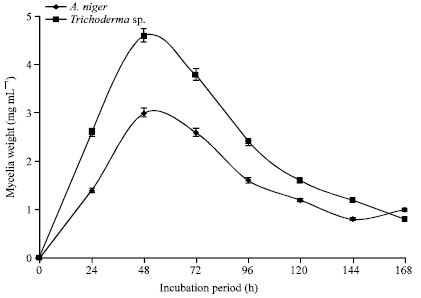 | |
| Fig. 7: | Mycelia weights of Aspergillus niger and Trichoderma sp. incubated at 25°C in modified Czapek-Dox liquid medium |
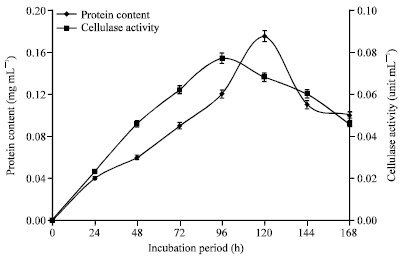 | |
| Fig. 8: | Protein content and cellulase activity of culture filtrates of A. niger incubated at 25°C in modified Czapek-Dox liquid medium |
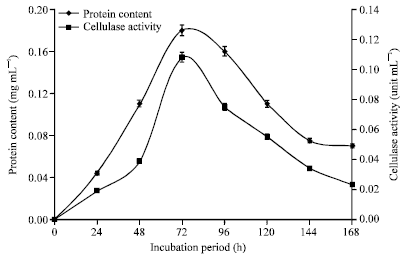 | |
| Fig. 9: | Protein and cellulase activity of culture filtrates of Trichoderma sp. incubated at 25°C in modified Czapek-Dox liquid medium |
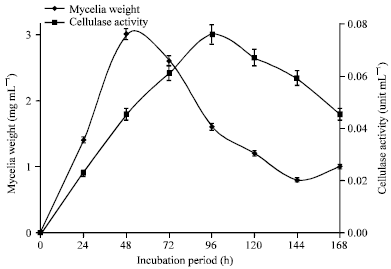 | |
| Fig. 10: | Mycelia weight and cellulase activity of culture filtrates of A. niger incubated at 25°C in modified Czapek-Dox liquid medium |
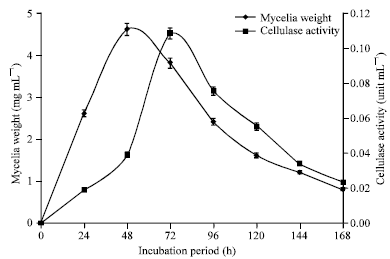 | |
| Fig. 11: | Mycelia weight and cellulase activity of culture filtrates of Trichoderma sp. incubated at 25°C in modified Czapek-Dox liquid medium |
The peak mycelia weight for each organism was obtained at 48 h of incubation. The peak cellulase activity for A. niger was obtained at 96 h whereas Trichoderma sp. gave its maximum cellulase activity at 72 h.
DISCUSSION
Wide variation in the nutritional requirements of microorganisms for growth and reproduction forms the basis for selective or differential media often engaged in microbiological studies (Harrigan and McCance, 1966). The ability of the isolated microfungi to thrive on modified Czapek-Dox agar containing cellulose as sole carbon source indicate that they can synthesize the enzymes required for the breakdown cellulose to meet their energy and carbon needs. The ability of some microfungi to grow on cellulosic materials was exploited in isolating the organisms from wood-wastes (Nwodo-Chinedu et al., 2005).
The growth of the organisms on different media (Sabouraud’s agar, Czapek-Dox agar, modified Czapek-Dox agar and modified Czapek-Dox liquid media) shows the versatility of the organisms to utilize different carbohydrate sources. However, these organisms utilized the carbon sources at different rates as indicated by their different rates of growth. The fastest of these organisms in terms of growth in all the media was Trichoderma sp. (Fig. 5-7). Trichoderma sp. belongs to a family of beneficial fungi commonly found in soils in nearly all parts of the world (Nederhoff, 2001). They are frequently the most prevalent culturable fungi in the soil where they basically live on plant debris and organic matter. Some strains are however parasitic on other fungi (Nederhoff, 2001). Fast growing strain of Trichoderma sp. such as the strain isolated in the present study can act as biological control agents, inhibiting the growth of phyto-pathogenic fungi that could destroy a large variety of crops (Cwdoza et al., 2005). Two of the isolated microfungi, Aspergillus flavus and A. niger, belong to the genus Aspergillus. This genus has been documented as source of the most prevalent airborne moulds (Nwodo-Chinedu et al., 2005). Aspergillus flavus is of great economic importance due to its ability to produce toxic and carcinogenic aflatoxins in agricultural commodities (Horn, 2006). Aspergillus flavus has, however, been safely used for many centuries in China in the production of various cheeses and soy sauce. Aspergillus niger is a filamentous ascomycete fungus that is ubiquitous in the environment and has been implicated in opportunistic infections of humans, yet industrial citric acid production by Aspergillus niger represents one of the most efficient, high-yield bioprocesses currently in use by industry (Baker, 2006). Strains of Penicillium sp. are perhaps best known for their use in pharmaceutical industry for the synthesis of penicillin and griseofulvin. Strains of Penicillium chrysogenum has been reported to produce significant levels cellulolytic enzymes (Nwodo-Chinedu et al., 2007).
There was a steady increase in the mycelia weight, protein released and cellulase activity in the culture media as the incubation progress until a specific period after which a decline set in (Fig. 7-10). The increase in the mycelia weight shows that the medium supports the growth of the organisms. Increase in protein level and cellulase activity of the culture filtrate shows that the organisms secrete extracellular proteins which have cellulolytic activity. Most cellulolytic fungi secrete hydrolytic enzymes for the breakdown of the polymers into their growth media; this largely accounts for the protein contents of the cell-free filtrates (Chinedu et al., 2008).
There were however different peak periods for mycelia, protein and cellulase production by the two microfungi studied. The mycelia weight after reaching the peak value in both organisms steadily declined afterwards (Fig. 7, 10, and 11). This trend was also noted in the protein content and cellulase activity (Fig. 8, 9). This is expected since there was no inflow of nutrients into the culture medium all through the incubation period. The pattern is characteristic of typical batch culture fermentation. The protein content and cellulase activity of the culture filtrates at different periods indicated a correlation. This was best depicted in Trichoderma sp. where the peak period for both protein and cellulase production was at 72 h of incubation (Fig. 9). The finding supports the trend previously reported for different microfungi (Nwodo-Chinedu et al., 2007; Chinedu et al., 2008). In the case of A. niger, the protein peak was at 120 h whereas cellulase activity peak was at 96 h (Fig. 8). Though the peak periods are very close, the difference seems to imply that the organism produces other proteins which may not have cellulolytic activity. Cellulose is known to also induce the production of other cell wall hydrolyzing enzymes such as xylanases in Aspergillus sp. (De Vries and Visser, 2001). In both organisms, the peak mycelium weight was reached before the cellulase activity peak (Fig. 10, 11). This seems to imply that the organisms continued to produce the cellulolytic enzymes even at the declining phase of the organisms’ growth. However, in each case, the cellulase activity peak was reached shortly after the peak mycelia weight and both continued to decrease afterwards.
The peak cellulase activity of the crude enzyme of A. niger and Trichoderma sp. was 0.077 and 0.108 unit mL-1 respectively. This is considered very significant. It compares with the cellulase activity peak of 0.10 unit mL-1 reported for a strain of Penicillium chrysogenum PCL 501 (Nwodo-Chinedu et al., 2007).
In conclusion, the isolates of A. niger and Trichoderma sp. cultivated in basal medium containing cellulose as sole carbon source have been shown to produce extracellular proteins with significant cellulase activity. These filamentous fungi can thus serve as potential tools in the industrial saccharification of cellulose. Fast growing fungi such as the strain of Trichoderma sp. isolated in the present study can act as biological control agent, inhibiting the growth of phyto-pathogenic fungi that could destroy a large variety of crops.
REFERENCES
- Abu, E.A., P.C. Onyenekwe, D.A. Ameh, A.S. Agbaji and S.A. Ado, 2000. Cellulase (EC 3.2.1.3) production from sorghum bran by Aspergillus niger SL1: An assessment of pre-treatment methods. Proceedings of the International Conference on Biotechnology: Commercialization and Food Security, October 21-23, 2000, Abuja, Nigeria, pp: 153-157.
- Baker, S., 2006. Aspergillus niger genomics: Past, present and into the future. Med. Mycol., 44: 17-21.
Direct Link - Chinedu, S.N., O.C. Nwinyi and V.I. Okochi, 2008. Growth and cellulose activity of wild-type Aspergillus niger ANL301 in different carbon sources. Can. J. Pure Applied Sci., 2: 357-362.
Direct Link - Cwdoza, R., M. Hermosa, J. Vizcaino, L. Sanz, E. Monte and S. Gutierrez, 2005. Secondary Metabolites Produced by Trichoderma and their Importance in the Biocontrol Process. In: Microorganisms for Industrial Enzymes Biocontrol, Mellado-Duran, E. and J.L. Barredo (Eds.). Research Sciencepost Publ., Kerala, India, pp: 207-228.
- De Vries, R.P. and J. Visser, 2001. Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiol. Mol. Biol. Rev., 65: 497-522.
CrossRefDirect Link - Howard, R.L., E. Abotsi, J.E.L. van Rensburg and S. Howard, 2003. Lignocellulose biotechnology: Issues of bioconversion and enzyme production: Review. Afr. J. Biotechnol., 2: 602-619.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Lynd, L.R., P.J. Weimer, W.H. van Zyl and I.S. Pretorius, 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev., 66: 506-577.
CrossRefPubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Nederhoff, E., 2001. Biological control of root diseases-especially with Trichoderma. Grower, 56: 24-25.
Direct Link - Nwodo-Chinedu, S., V.I. Okochi, H.A. Smith, U.A. Okafor, B.M. Onyegeme-Okerenta and O. Omidiji, 2007. Effect of carbon sources on cellulase (EC 3. 2. 1. 4) production by wild-type Penicillium chrysogenum PCL501. Afr. J. Biochem. Res., 1: 6-10.
Direct Link - Peciulyte, D., 2007. Isolation of cellulolytic fungi from waste paper gradual recycling materials. Ekologija, 53: 11-18.
Direct Link - Romanelli, R., C. Houston and S. Barnett, 1975. Studies on thermophilic cellulolytic fungi. Applied Microbiol., 30: 276-281.
Direct Link
christian Reply
please I need an extract of Trichoderma reesei
S.N. Chineedu
Unfortunately, we do not any strain of Trichoderma sp. in the lab presently.
Thanks.