
Olusola O. Shonubi
Not Available
O.T. Okusanya
Not Available
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 6 | Page No.: 949-956







ABSTRACT
The dominance of Paspalum vaginatum in mangrove swamp at the University of Lagos, Nigeria, was investigated by determining its growth and physiological responses to various salinity levels. P. scrobiculatum which occurs further inland was included for comparison. Uniform cuttings of each species were subjected to salinity levels of 0-400 mM NaCl using artificial seawater in one-fifth strength Hoagland solution in a glasshouse for 28 days. The results showed that P. vaginatum behaved like a halophyte showing enhanced growth at low salinity and tolerating high salinity with no significant decrease in water content but with significant increase in chlorophyll content as salinity increased. On the other hand, P. scrobiculatum behaved like a non-halophyte with decreased growth and significant reduction in water content but with significant increase in chlorophyll content as salinity increased and is incapable of tolerating high salinity. The results indicate that P. vaginatum tolerated salinity by regulating the amount of salt in the shoot while retaining enough water for osmotic adjustment. P. scrobiculatum could neither regulate salt uptake, nor retain enough water for osmotic adjustment. It was concluded that these responses may be partly responsible for the ability of the species to inhabit their different environments.
PDF Abstract XML References Citation
How to cite this article
Olusola O. Shonubi and O.T. Okusanya, 2007. The Growth and Physiological Responses of Paspalum vaginatum S.W. and Paspalum scrobiculatum Linn. in Relation to Salinity
. Asian Journal of Plant Sciences, 6: 949-956.
DOI: 10.3923/ajps.2007.949.956
URL: https://scialert.net/abstract/?doi=ajps.2007.949.956
DOI: 10.3923/ajps.2007.949.956
URL: https://scialert.net/abstract/?doi=ajps.2007.949.956
INTRODUCTION
Field observation showed that the coastal flood plain around the University of Lagos is dominated by the monocotyledon, Paspalum vaginatum along with other salt marsh species like Avicenia nitida, Rhizophora racemosa and Drepanocarpus lunatus. The marsh is inundated at high tide thus the area is highly saline. Nwanko (1984) recorded salinities up to that of sea water during the dry season. We were thus interested in finding out how P. vaginatum is able to dominate the saline flood plain. Further inland, P. vaginatum is absent but there is the abundance of a closely related species, P. scrobiculatum. In view of this, we decided to include P. scrobiculatum in the investigation for comparison. P. vaginatum is a troublesome weed on rice lands in Sierra Leone and it has already been identified as a good fodder grass (Stanfield, 1970). The United States Golf Association (USGA) is considering using it on golf courses that are prone to salinity (Lee et al., 2002).
A major factor that we decided to test was the tolerance of the species to salinity. Greenway and Munns (1980) reported that the growth of many monocotyledonous halophytes is progressively inhibited with increasing salinity unlike the dicotyledonous halophytes where growth is often optimal in the presence of about 50% sea water. However, other grasses show slight increase in growth at low salinity (Blits and Gallagher, 1991; Short and Colmer, 1999; Lee et al., 2002). Flowers and Yeo (1988) also elucidated the physiological basis of salt tolerance in monocotyledonous halophytes. Lee et al. (2005) proposed criteria for assessing tolerance in Paspalum.
Our aim in this study was to test how the growth of the two species is affected by various levels of salinity and to determine their physiological responses to salinity. Relatively little information is available for temperate monocotyledonous halophytes, more so for tropical halophytes. This investigation will add to the meager literature on tropical monocotyledonous halophytes.
Paspalum vaginatum S.W. is a practically hairless perennial with creeping shoots. It has erect stems with closely overlapping leaf sheaths and green to dark green leaf blades. The inflorescence has two branches at the apex of the central stalk, occasionally with one lower down. It usually colonizes new areas by producing seeds in the rainy season but propagates vegetatively by stolons in well established stands. It can be found in southern Nigeria in states along tidal flats in the mangrove vegetation (Lowe, 1989).
Paspalum scrobiculatum Linn. is a more or less tufted, strangling perennial, approximately 60 cm high. It has variably hairy leaf blades and ribbon-like inflorescence with two to ten branches (Lowe, 1989). It is widespread and abundant throughout the wetter parts of Nigeria in moist and shady places.
MATERIALS AND METHODS
The growth response of P. vaginatum and P. scrobiculatum to salinity was tested using uniform cuttings from each species as the seeds do not germinate easily. Each cutting contained three nodes with no tillers. The cuttings were planted in acid washed sand medium in 8.8 cm diameter and 8 cm deep plastic pots. They were allowed to establish for one week during which time they were watered with one-fifth strength Hoagland’s solution. (Hoagland and Arnon, 1950).
After one week, the plants were treated with saline solutions at concentrations of 0, 50, 100, 200, 300 and 400 mM NaCl. The saline solutions were prepared by mixing different proportions of Artificial Sea Water (ASW) with one-fifth strength Hoagland solution. The pH of the solutions was adjusted to 6.5 with dilute nitric acid. Each treatment had ten replicates.
The plants at high salinities were brought up to their salinities by step-wise increment of 50 mM at 2 day interval. Watering was done by dipping the sand medium in the appropriate solution for about 15 min every other day. The solutions were changed weekly and the sand medium leached weekly to prevent salinity buildup.
The experiment was carried out in 1999, in the greenhouse of the University of Sussex, England with 12 h photoperiod. The greenhouse has a photo flux of between 400 and 1000 μmoles m-2 sec-1 in the range of Photosynthetic Active Radiation (PAR). Solar lighting was from high-pressure sodium lamps (GEC Son T 400W, Complex plant care Ltd.). Daytime temperature ranged between 23 and 25°C and night temperature was between 15 and 18°C. Relative humidity was 58% during the day and 62% at night.
At the end of the experiment, total fresh weights and chlorophyll contents from sub-samples were determined, and then the roots and shoots were dried for 24 h in a fan oven (Townsend and Mercer Ltd., Croydon) at 70°C, weighed and analyzed for mineral content. Chlorophyll was estimated using the method of Arnon (1949). Water content was calculated as difference between fresh and dry weights of the plants.
Extraction in 100 mM acetic acid was used for the mineral content analysis. Mineral cations namely sodium, potassium, calcium and magnesium were estimated using a Pye Unicam Atomic Absorption Spectrophotometer model SP9 with computer attachment.
Chloride was estimated with the EIL chloride electrode used in conjunction with a vibron electrometer. The ion contents in the dead and young leaves, culms and roots of each species were determined separately.
Data obtained from dry weight values were used to calculate the Leaf Weight Ratio (LWR), Stem Weight Ratio (SWR), Root Weight Ratio (RWR) and shoot root ratio (S:R). The University of Sussex computer and statistical package Minitab were used for the analysis of the data.
RESULTS
Analysis of variance (ANOVA) of the effect of salinity on mean total dry weight showed that there was a significant effect of salinity on the growth of each species. (p<1% for P. vaginatum and p<0.1% for P. scrobiculatum). In P. vaginatum, there was a slight increase in dry weight (growth) up to 100 mM salinity, thereafter, there was a significant decrease as salinity increased (p<1% for 200 mM and p<0.1% for 300 mM and 400 mM). In P. scrobiculatum, there was a decrease in dry weight as salinity increased. Above 50 mM salinity, the decrease was significant (p<0.1%) (Fig. 1).
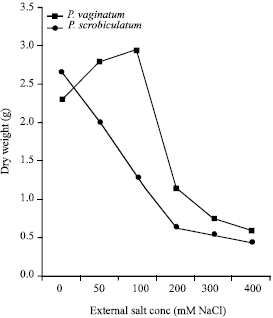 | |
| Fig. 1: | The dry weight of Paspalum vaginatum and P. scrobiculatum in relation to salinity |
| Table 1: | Effect of salinity on proportion of matter in the leaves (LWR), stem (SWR) and roots (RWR) of Paspalum vaginatum and P. scrobiculatum. Mean of ten replicates (±SE) |
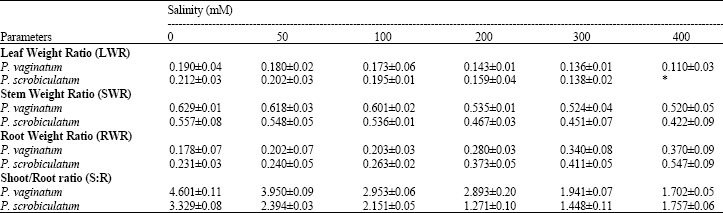 | |
| *: Leaves dead | |
| Table 2: | Effect of salinity on chlorophyll concentration, magnesium concentration and water content of Paspalum vaginatum and P. scrobiculatum |
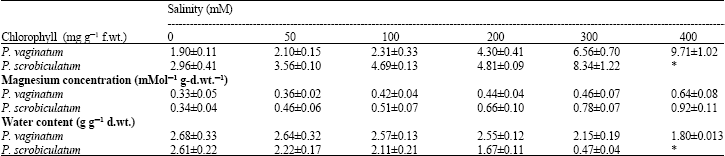 | |
| *: Leaves dead | |
The proportion of dry matter in leaves, stem and roots expressed in relation to the total dry weight (LWR, SWR and RWR) are shown in Table 1. In both species, LWR decreased with increasing salinity, but P. vaginatum had lower values than P. scrobiculatum where no leaves survived at 400 mM salinity. The SWR values in both species showed a decrease with increasing salinity. The values in P. vaginatum were higher than in P. scrobiculatum at all treatments. In both species, there was a gradual increase in RWR values as salinity increased. P. vaginatum however had lower values than P. scrobiculatum. The S:R values in both species decreased as salinity increased, but with the values in P. vaginatum being higher than in P. scrobiculatum (Table 1).
Chlorophyll concentration increased with increasing salinity in both species, but P. vaginatum had lower values than P. scrobiculatum at all salinity levels. The decrease in water content as salinity increased was insignificant in P. vaginatum and the values were higher than in P. scrobiculatum where there was significant decrease as salinity increased (Table 2).
Mineral element content:
Sodium ion concentration: Analysis of variance (ANOVA) showed that there was a significant effect of salinity on sodium concentration in the whole plant of P. vaginatum (p<0.1%) as its total concentration increased with increase in salinity. The amount found in the root was much higher than that in either the stem or the fresh leaves, but it was lower than in the shoot (stem + leaves). The amount found in the dead leaves increased with increasing salinity and is much higher than those found in the fresh green leaves (Table 3).
ANOVA showed that there was a significant effect of salinity on the sodium concentration in the whole plant of P. scrobiculatum (p<0.1%). The accumulation of sodium in P. scrobiculatum followed the same pattern as that found in P. vaginatum, in that the concentration in the whole plant increased with increase in salinity. It is noteworthy that the total amounts in P. scrobiculatum at higher salinities (200-400 mM) were significantly higher than in P. vaginatum despite the smaller dry matter of P. scrobiculatum (Table 3).
The amounts of sodium ions in the roots of P. scrobiculatum were higher than those found in the stem, which were also higher than those found in the leaves except at 0 mM (Table 3). P. scrobiculatum also had a higher amount of sodium in the shoot than in the root. The amounts in the dead leaves increased with increasing salinity and were much lower than those in the fresh leaves, stem and root. Again, it is noteworthy that at virtually all salinities, P. scrobiculatum had higher sodium content values than P. vaginatum.
| Table 3: | Concentration of sodium and chloride ions in the plant of Paspalum vaginatum and P. scrobiculatum |
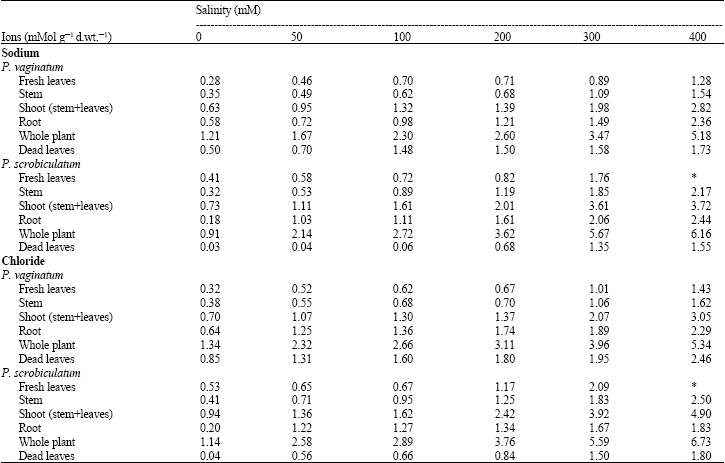 | |
| *: Leaves dead | |
Chloride ion content: The total ion concentration of chloride among treatments in the whole plant of P. vaginatum was significantly different (p<0.1%), with the values increasing as salinity increased (Table 3). Like the sodium ions, the concentration of chloride ions in the root was higher than the amount in the stem, which was also higher than that in the fresh leaves. The amounts in the dead leaves increased with increased salinity and they were higher than those in the fresh green leaves (Table 3).
There was a significant effect of salinity on chloride concentration in the whole plant of P. scrobiculatum (p<0.1%). The amount in the whole plant and plant parts also followed the same patterns as those of sodium, increasing with increased salinity (Table 3). Its concentrations in the dead leaves were lower than those in the fresh green leaves, stem and root at all salinity levels, much like the sodium ions (Table 3).
The chloride values in the fresh leaves and stem and consequently the shoot, were higher at all salinities in P. scrobiculatum than in P. vaginatum, but the reverse was the case in the root (Table 3). Thus for the whole plant in both species, there was not much difference in the sodium and chloride ion concentrations at low salinity (0-100 mM), but at higher salinities, P. scrobiculatum accumulated significantly greater amount of salt than P. vaginatum (Table 3).
Potassium ion content: ANOVA of the data on the effect of salinity on potassium ion concentration in the whole plant of P. vaginatum showed that there was a significant effect between treatments (p<5%). Its concentrations in the whole plant, the shoot, stem and root of P. vaginatum decreased as salinity increased. The values however increased in the fresh and dead leaves with increasing salinity (Table 4).
In P. scrobiculatum, there was also a significant effect on the amounts of potassium ion in the whole plant at each treatment (p<0.1%). The concentration of potassium ion in the fresh leaves, stem, root and consequently the whole plant of P. scrobiculatum decreased with increasing salinity (Table 4). Its concentration in the dead leaves however increased with increasing salinity. Comparing both species, the values in P. vaginatum were generally higher than those in P. scrobiculatum at all salinities (Table 4).
Na:K: Sodium to potassium ratio increased with increasing salinity in both species. At all salinity levels, P. scrobiculatum had higher values than P. vaginatum especially at high salinities where the values were more than double those in P. vaginatum (Table 4).
| Table 4: | Potassium ion content and Na:K ratios in the plant parts of Paspalum vaginatum and P. scrobiculatum |
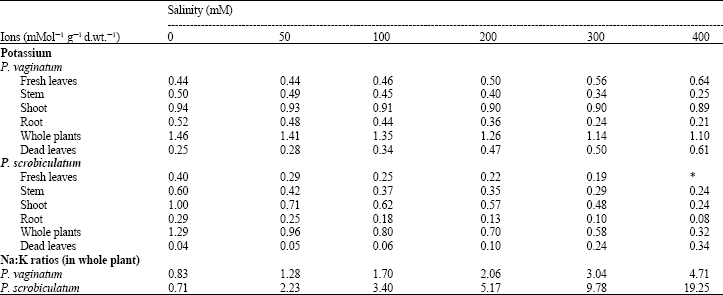 | |
| *: Leaves dead | |
Magnesium ion concentration: Like the other ions, the concentrations of magnesium in the whole plants of both species were affected by salinity (p<0.1). Its concentrations in both species increased with increasing salinity, but the values for P. scrobiculatum were significantly higher than for P. vaginatum (Table 2).
DISCUSSION
Growth as measured by dry weight of the more extreme dicotyledonous halophytes is typically stimulated by salinity, being optimal at low to moderate concentrations (50-250 mM) of NaCl (Okusanya, 1979; Okusanya and Ungar, 1984; Short and Colmer, 1999; Debez et al., 2004). Some halophytes like the Atripex species (Chenopodiaceae), (Longstreth and Nobel, 1979) and Plantago maritima (Plantaginaceae) (Erdei and Kuiper, 1979) do not show any increase in growth in response to increasing salinity. The results of this experiment showed that P. vaginatum behaved like a true halophyte showing slight increase in dry weight at low salinity while P scrobiculatum behaved like a non-halophyte with decreased dry weight as salinity increased (Fig. 1). The presence of P. vaginatum in saline mangrove environment shows a relationship to its ability to grow well at low salinity and to survive at high salinity as found out in this experiment. The converse seems to be true for P. scrobiculatum.
For the monocotyledonous halophytes, growth is not generally stimulated by saline conditions, but if it is, the stimulation is mild. (Munns et al., 1983; Blits and Gallagher, 1991; Lee et al., 2002). This is seen to be true in this experiment for the monocotyledonous P. vaginatum, where there was a slight increase in growth at 50-100 mM NaCl (Fig. 1).
The better performance of P. scrobiculatum than P. vaginatum in the control set up, (0 mM) (Fig. 1) might be explained in terms of their natural habitat. Since P. scrobiculatum does not live naturally in saline soil as it grows further inland than P. vaginatum, the non-saline culture medium is a more natural condition. Unlike P. vaginatum which may require low salinity for maximum growth as reported for some other halophytes. (Okusanya, 1980; Flowers, 1985; Matoh et al., 1988; Yeo and Flowers, 1990).
In the present study, all the growth data suggested that P. scrobiculatum was more sensitive to salinity than P. vaginatum, especially at high salinities. At low salinity, P. scrobiculatum had less than half the dry weight of P. vaginatum. By the end of the experiment, the leaves of P. scrobiculatum were all dead at the highest salinity while those of P. vaginatum were still alive, albeit rolled up at the edge, a condition which promotes greater water-use efficiency in halotolerant grasses (Blits and Gallagher, 1991). This shows that P. vaginatum is better able to tolerate higher salinity than P. scrobiculatum. This is probably another reflection of their ecological habitats. The salinity of the Lagos Lagoon which floods the marsh where P. vaginatum grows, can reach nearly that of sea water during the dry season (Nwankwo, 1984). P. scrobiculatum grows further inland where there is little or no salinity.
The increased Root Weight Ratio (RWR) exhibited by both species (Table 1) is a response to salinity stress. (Okusanya, 1979; Okusanya and Ungar, 1984; Donovan and Gallagher, 1985). Increased RWR in plants results in an effort to absorb more nutrients and water. The greater RWR in P. scrobiculatum than in P. vaginatum indicated that it was under greater salinity stress.
The increased chlorophyll content with increased salinity probably indicated that both species were responding to salinity stress by increasing chlorophyll content with a view to making more matter. P. scrobiculatum again appears to be under greater stress by making more chlorophyll (Table 2). While the results for P. vaginatum agreed with those reported by Blits and Gallagher (1991), Ashraf and Bhatti (2000), Murillo-Amador et al. (2002) and El-Hendaw et al. (2005) for salt tolerant ecotypes and crops, the results for P. scrobiculatum were in contrast to the salt sensitive ecotypes and crops reported.
Both species appear to respond and adapt to salinity by lowering their osmotic potential through the absorption of ions. Reduction in water potential was achieved by an increase in the internal concentration of NaCl. However, a mechanism for tolerating salinity is to regulate the amount of salt uptake. Species must accumulate sufficient ions in their leaves for osmotic adjustment, while avoiding toxic effects of these ions (Gorham, 1995; Munns, 2002). Table 3 indicated that P. scrobiculatum accumulated more sodium and chloride ions in the shoot than P. vaginatum. In the root, the reverse was the case. Thus, it appears that P. scrobiculatum was unable to regulate salt uptake, a process that may result in build up of ions to toxic level. The death of leaves at the highest concentration in P. scrobiculatum and the high concentration of ions in such leaves support this assertion.
Munns (2002) and El-Hendaw et al. (2005) reported low sodium contents in the leaves of salt tolerant species and ecotypes. P. vaginatum has lower sodium content so it is able to regulate the uptake of ions into its shoot, keeping relatively large amount in the roots for osmotic adjustment. The high salt concentration in the older (dead) leaves in P. vaginatum (Table 3) seems to indicate that the species may also be removing excess salt through the older leaves while maintaining favorable levels in the fresh leaves. These processes make P. vaginatum a more salt tolerant species than P. scrobiculatum and it can thrive well in the saline environment. Munns (2002) concluded that most glycophytes have a poor ability to exclude salt and it concentrates to toxic levels in the transpiring leaves.
In some halophytes, ions principally sodium and chloride, can amount to between 30 and 50% of dry weight generating Na:K ratios in excess of 10. (Flowers et al., 1986). Higher Na:K ratios were observed in P. scrobiculatum than P. vaginatum (Table 4). This is an indication of sodium stress which further confirmed that P. scrobiculatum was unable to restrict the entry of sodium thus making it a non-halophyte. Hannon and Barber (1972), Flowers and Yeo (1988) and Al-Karaki (2000) observed low Na:K ratios in other halophytic grasses as was observed in P. vaginatum. The result is also in line with Albert and Kinzel (1973) concept of physiotypes in grasses and related groups with respect to salt tolerance. They reported that such physiotypes utilize potassium and sugar for osmotic adjustment, thus they have high concentration of potassium ion and low concentration of sodium ion resulting in low Na:K ratio as in P. vaginatum (Table 3, 4).
The consistently lower values for the accumulated ions in P. vaginatum than in P. scrobiculatum (Table 3) agrees with the observations that halotolerant ecotypes of Juncus and Festuca accumulated less NaCl than did the salt sensitive ecotypes (Rozema, 1976; Rozema et al., 1978). The more salt-tolerant Agrostis stolonifera and Festuca rubra from salt marshes had lower sodium contents than their less salt tolerant counterparts from non-saline areas (Tiku and Snaydon, 1971). The exclusion of harmful ions from the shoots contributes to salt tolerance (Munns, 1993).
As a component of chlorophyll, the increased magnesium concentration as salinity increased in both species (Table 3) correlates well with the increased concentration in chlorophyll. The higher values in P. scrobiculatum than in P. vaginatum may be a response to greater salinity stress. This result is in contrast to other results (Glenn et al., 1996; Khan et al., 2000; Debez et al., 2004) but it showed that the species may not be under magnesium deficiency.
The insignificant decrease in water content as salinity increased and the higher water content in P. vaginatum is probably another reason that makes it more salt tolerant than P. scrobiculatum since it would have more water to dilute the relatively low level content of absorbed ions in its shoot, thus keeping osmotic concentration to tolerable level. Debez et al. (2004) also reported no significant effect of salinity on leaf hydration but a significant increase in leaf succulence. These two processes may enhance better salt tolerance in P. vaginatum.
In conclusion, P. vaginatum is tolerant of salinity adopting a variety of methods including increased osmotic potential, keeping adequate water content in the shoot and regulating the amount of salt in the shoot. Other methods may include the ability to take up potassium and the removal of excess salt through the old leaves. Thus its halophytic nature may partly account for its dominance of the coastal saline mangrove around the University of Lagos and similar habitat. On the other hand, P. scrobiculatum behaved like a non-halophyte being intolerant of salinity largely because it cannot regulate the amount of salt in its shoot. It thus inhabits non-saline areas further inland from the mangrove. The results of other experiments will be published later.
ACKNOWLEDGMENTS
We thank the authority of the University of Lagos for permission to carry out this project. Dr. Shonubi would like to thank the University of Sussex, Brighton, U.K for providing space and material through a British Council Chevening Scholarship. Many thanks to Prof. T.J. Flowers, Mr. Femi Ogundele, Mrs. Sue Scanzillo and Ms. A. Kloczko for their help.
REFERENCES
- Al-Karaki, G.N., 2000. Growth, water use efficiency and sodium and potassium acquisition by tomato cultivars grown under salt stress. J. Plant. Nutr., 23: 1-8.
Direct Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Blits, K.C. and J.L. Gallagher, 1991. Morphological and physiological responses to increased salinity in marsh and dune ecotypes of Sporobolus virginicus (L.) Kunth. Oecology, 87: 330-335.
Direct Link - Debez, A., K.B. Hamed, C. Grignon and C. Abdelly, 2004. Salinity effects on germination, growth and seed production of the halophyte Cakile maritima. Plant Soil, 262: 179-189.
Direct Link - El-Hendawy, S.E., Y. Hu and U. Schmidhalter, 2005. Growth, ion content, gas exchange and water relations of wheat genotypes differing in salt tolerances. Aust. J. Agric. Res., 56: 123-134.
Direct Link - Glenn, E.R., Pfister, J.J. Brown, T.L. Thompson and J. O`leary, 1996. Na and K accumulation and salt tolerance of Atriplex canescens (Chenopodiaceae) genotypes. Am. J. Bot., 83: 997-1005.
Direct Link - Greenway, H. and R. Munns, 1980. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol., 31: 149-190.
CrossRefDirect Link - Khan, M.A., A.U. Irwin and A.M. Showalter, 2000. Effects of salinity on growth, water relations and ion accumulation of the subtropical perennial halophyte, Atriplex griffithii var. stocksii. Ann. Bot., 85: 225-232.
CrossRefDirect Link - Lee, G., R.N. Carrow and R.R. Duncan, 2005. Criteria for assessing salinity tolerance of the halophytic turfgrass seashore paspalum. Crop Sci., 45: 251-258.
Direct Link - Munns, R., 1993. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell Environ., 16: 15-24.
CrossRefDirect Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Murillo-Amador, B., E. Troyo-Dieuez, R. Lopezaguilar, A. Lopez-Cortes, C.L. Tinoco-Ojanguren, H.G. Jones and C. Kaya, 2002. Matching physiological traits and ion concentrations associated with salt stress in cowpea genotypes. Aust. J. Agric. Res., 53: 1243-1255.
Direct Link - Tiku, B.L. and R.W. Snaydon, 1971. Salinity tolerance within the grass species, Agrostis stolonifera L. Plant Soil, 35: 421-431.
CrossRefDirect Link