
Renu Bhardwaj
Department of Botanical and Environmental Sciences, Guru Nanak Dev University, Amritsar-143005 Punjab, India
Nitika Arora
Department of Botanical and Environmental Sciences, Guru Nanak Dev University, Amritsar-143005 Punjab, India
Priyanka Sharma
Department of Botanical and Environmental Sciences, Guru Nanak Dev University, Amritsar-143005 Punjab, India
Hardesh Kumar Arora
Department of Botanical and Environmental Sciences, Guru Nanak Dev University, Amritsar-143005 Punjab, India
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 5 | Page No.: 765-772







ABSTRACT
The effects of 28-homobrassinolide (28-homoBL) on seedling growth, lipid peroxidation and antioxidative enzyme activities under Nickel (Ni) stress in the seedlings of Zea mays L. (var. Partap-1) were studied. The surface sterilized seeds of Zea mays were given treatments of different concentrations of 28-homoBL (10-4, 10-6 and 10-8 mM) and heavy metal Ni (0.5, 1.0, 1.5 and 2.0 mM) in combination for 7 days. The activities of superoxide dismutase (EC 1.15.1.1) catalase (EC 1.11.1.6), ascorbate peroxidase (EC 1.11.1.11), guaiacol peroxidase (EC 1.11.1.7) and glutathione reductase (EC 1.6.4.2) of 7-days old seedlings were analyzed. It was observed that 28-homoBL reduced the toxicity of heavy metal on seedling growth considerably and also influenced protein content. Lipid peroxidation level was significantly increased under heavy metal treatments alone but lowered with 28-homoBL applications revealing less oxidative damage. 28-homoBL treatments expressed anti-stress activity by regulating the activities of antioxidative enzymes. It was observed that activities of all antioxidative enzymes increased except for superoxide dismutase as compared to control.
PDF Abstract XML References Citation
How to cite this article
Renu Bhardwaj, Nitika Arora, Priyanka Sharma and Hardesh Kumar Arora, 2007. Effects of 28-homobrassinolide on Seedling Growth, Lipid Peroxidation and Antioxidative Enzyme Activities under Nickel Stress in Seedlings of Zea mays L. Asian Journal of Plant Sciences, 6: 765-772.
DOI: 10.3923/ajps.2007.765.772
URL: https://scialert.net/abstract/?doi=ajps.2007.765.772
DOI: 10.3923/ajps.2007.765.772
URL: https://scialert.net/abstract/?doi=ajps.2007.765.772
INTRODUCTION
Brassinosteroids (BRs) are recently explored group of phytohormones, which have structures similar to animal steroid hormones and are distributed throughout the plant kingdom (Clouse and Sasse, 1998; Mandava, 1988; Rao et al., 2002; Bhardwaj et al., 2006). They play an essential role in plant growth and development and have been implicated in many physiological responses, including cell expansion, vascular differentiation, reproductive development, seed germination and promotion of root growth (Khripach et al., 2000; Cao et al., 2005). These phytohormones also promote plant tropisms by modulating polar auxin transport (Xu, 2006). Alongwith growth promoting effects, they are also reported to confer resistance to plants against various abiotic/biotic stresses like heat, drought, heavy metals, infection, pesticides, salt and even viruses (Dhaubhadel et al., 1999, 2002; Krishna, 2003; Wachsman et al., 2000, 2002, 2004; Upreti and Murti, 2004).
Though heavy metals are essential as micronutrients for plants, but at higher concentration they are toxic (Dalton et al., 1988). Ni as an essential element has a role in several metabolic processes of plants (Brown et al., 1987; Welch, 1995; Bai et al., 2006; Rahman et al., 2005). However excess of Ni is indicated by a decrease in growth and development, rate of photosynthesis and an induction of oxidative damage due to the production of Reactive Oxygen Species (ROS) such as superoxide radical (O2• ¯), hydrogen peroxide (H2O2), hydroxyl radical (OH•) and alkoxyl radical (RO•) in various plant species (Marschner, 1995). These ROS have the capacity to initiate lipid peroxidation and degrade proteins, lipids and nucleic acids (Costa et al., 2002; Lu et al., 2005). These ROS are removed by enzymatic antioxidant system consisting of Superoxide Dismutase (SOD), Guaiacol Peroxidase (POD), Ascorbate Peroxidase (APOX), Catalase (CAT) and Glutathione Reductase (GR) (Asada and Takahashi, 1987; Salin, 1988). BRs help to overcome the stress by regulating the activities of these antioxidative enzymes (Ozdemir et al., 2004). The present study was therefore aimed to investigate the effects of 28-homobrassinolide (28-homoBL) on seedling growth, lipid peroxidation and antioxidative enzyme activities under the heavy metal (Ni) stress in the seedlings of Zea mays L. (var. Partap-1).
| Table 1: | F-ratio of two–way ANOVA of 28-homoBL treatments for morphological parameters (Shoot and root length, fresh weight of shoots and roots) |
 | |
| (* Indicate statistically significant differences from control at p<0.05) | |
| Table 2: | F-ratio of two–way ANOVA of 28-homoBL treatments for biochemical parameters (Protein content, MDA content, SOD, CAT, POD, APOX and GR) |
 | |
| (* Indicate statistically significant differences from control at p<0.05) | |
MATERIALS AND METHODS
Seeds of maize (Zea mays L. var. Partap-1) were obtained from Punjab Agriculture University, Ludhiana, Punjab, India. The experiment was conducted at Department of Botanical and Environmental Sciences, GNDU, Amritsar, Punjab, India in Dec 2006. Uniform sized seeds of maize were surface sterilized with 0.05% mercuric chloride for 5 min followed by three rinses in sterile distilled water. The surface sterilized seeds were germinated in Whatman No.1 filter paper lined glass Petriplates (10 cm diameter, 8 seeds per Petriplate) containing different concentrations of Ni (0.5, 1.0, 1.5 and 2.0 mM) alone, 28-homoBL (10-4,10-6 and 10-8 mM) alone and Ni (0.5, 1.0, 1.5 and 2.0 mM) supplemented with 28-homoBL (10-4,10-6 and 10-8 mM). The Ni was given in the form of NiSO4.7H2O. Each Petriplate was supplied with 4 mL of test solution on first day and 2 mL of test solution on alternate days, up to day 7. Control seedlings were supplied with distilled water. Each treatment was replicated 5 times. The experiment was conducted under controlled conditions (20°C ± 5°C, 16 h photoperiod). On 7th day, seedlings were harvested and shoots and roots of seedlings were separated. Seedling growth in terms of length and fresh weight was recorded. Thirty seedlings per treatment were used for measurement of morphological parameters (root/shoot length and root/shoot fresh weight). Experiment was repeated twice.
Lipid peroxidation was determined as the content of malondialdehyde (MDA) using the thiobarbituric acid (TBA) reaction as described by Heath and Packer (1968). For estimation of antioxidative enzyme activities and protein content, 1 g leaves of 7 day old seedlings were homogenized in 3 mL of 100 mM potassium phosphate buffer (pH = 7). The homogenate was centrifuged at 4°C for 20 min at 15,000 g. The supernatant was used for assays of the activities of SOD, POD, CAT, GR and APOX. The activity of SOD was determined by monitoring its ability to inhibit photochemical reduction of nitrobluetetrazolium (NBT) at 540 nm (Kono, 1978). POD activity was determined according to Putter (1974). CAT activity was determined by following the initial rate of disappearance of H2O2 at 240 nm (Aebi, 1983). The activities of APOX and GR were measured by the method of Nakano and Asada (1981); Carlberg and Mannervik (1975) respectively. Protein content was determined following the method of Lowry et al. (1951). The data were processed by two-way Analysis of Variance (ANOVA) and comparisons with p values <0.05 were considered significantly different. Standard error due to replicates was calculated (Table 1 and 2).
RESULTS
The effect of 28-homoBL on seedling growth under Ni stress expressed as length and fresh weight of shoots and roots, are shown in Fig. 1. Growth of seedlings decreased with the increase of concentration of Ni as compared to control. However supplementation of Ni solution with 28-homoBL considerably reduced the inhibitory effect of Ni on seedling growth. The root length (18.07 cm) of seedlings treated with 10-6 mM 28-homoBL supplemented with 0.5 mM Ni solution was maximum as compared to control (seedlings treated with Ni alone) (12.57 cm). Similarly the shoot length of seedlings decreased as the concentration of metal increased and this decrease was maximum (2.96 cm) in case of seedlings treated with 2 mM of Ni. Maximum increase in shoot length (7.1 cm) was observed in case of seedlings treated with 10-8 mM 28-homoBL supplemented 0.5 mM of Ni solution. Similar trends were observed for fresh weight of root and shoot of seedlings. Soluble protein content of seedlings increased significantly in all treatments of 28-homoBL as compared to control (Fig. 2).
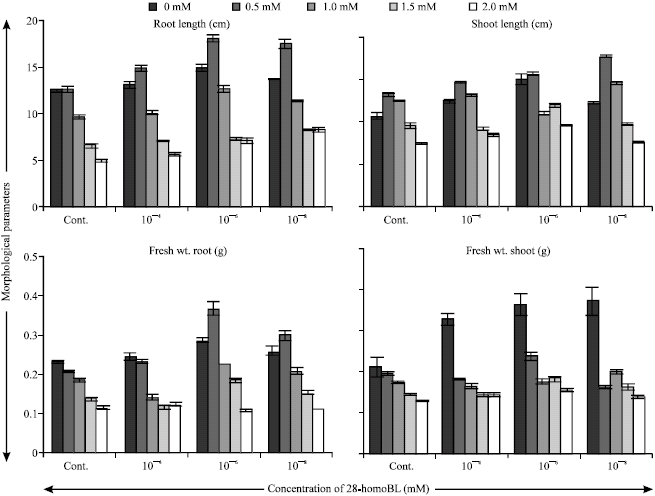 | |
| Fig. 1: | Effect of 28-homoBL on morphological parameters of 7-days old Zea mays seedlings under Ni stress. Bar represents the SE (n = 30) |
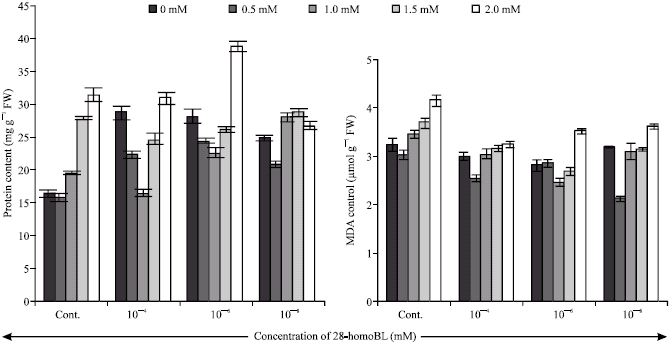 | |
| Fig. 2: | Effect of 28-homoBL on protein and malondialdehyde (MDA) content of 7-days old Zea mays seedlings under Ni stress. Bar represents the SE (n = 5) |
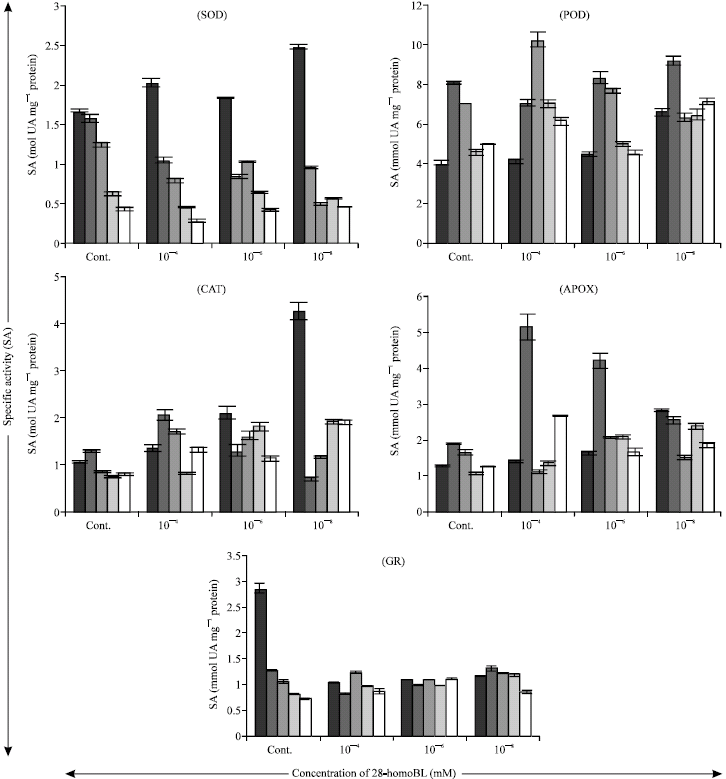 | |
| Fig. 3: | Effect of 28-homoBL on Specific Activities (SA) of antioxidative enzymes (superoxide dismutase (SOD), guaiacol peroxidase (POD), catalase (CAT), ascorbate peroxidase (APOX) and glutathione reductase (GR)) of 7-days old Zea mays seedlings under Ni stress. Bar represents the SE (n = 5) |
The protein content was remarkably higher (38.79 mg g-1 FW) in the seedlings treated with 10-6 mM of 28-homoBL supplemented with 2.0 mM of Ni as compared to control (31.48 mg g-1 FW). The concentration of MDA (Fig. 2) increased under heavy metal stress but decreased with 28-homoBL applications. Minimum content of MDA (2.12 μmol g-1 FW) was observed in 10-8 mM 28-homoBL supplemented 0.5 mM of Ni solution as compared to control (3.02 μmol g-1 FW).
SOD activity got decreased by the treatment of Ni in maize seedlings (Fig. 3). Decreased activity of SOD was not alleviated by any of the concentration of 28-homoBL. On the other hand the CAT activity of 28-homoBL treated seedlings was increased in comparison to control. CAT showed maximum activity (2.044 mol UA mg-1 protein) in case of seedlings treated with 10-4 mM 28-homoBL supplemented 0.5 mM of Ni solution (Fig. 3). The activity of POD also increased under the influence of 28-homoBL as compared to control. Maximum activity (10.23 m mol-1 UA mg protein) of POD was observed in seedlings treated with 10-4 mM 28-homoBL supplemented 1.0 mM of Ni solution (Fig. 3). Similarly APOX and GR activities were also enhanced by applications of different concentrations of 28-homoBL. Maximum enhanced activity of APOX (5.137 m mol-1 UA mg protein) and GR (1.305 m mol-1 UA mg protein) was observed in case of seedlings, treated with 10-4 mM and 10-8 mM 28-homoBL supplemented 0.5 mM of Ni solution, respectively (Fig. 3).
DISCUSSION
The present study shows that application of 28-homoBL improved the seedling growth, increased protein content, decreased the MDA content and increased activities of CAT, POD, APOX and GR except of SOD under Ni stress.
Plant growth via cell elongation and cell division requires the coordination of several processes, some of them appears to be influenced by BRs. Plasticity of the cell wall is increased when proton extrusion by H+- ATPases acidifies the apoplast, thereby activating the cell wall loosening enzymes. It increases the synthesis of new cell wall and membrane materials. BRs have been found to increase the ATPase activity in Azuki bean epicotyls and maize roots, leading to proton extrusion and induced cell wall relaxation (Cerana et al., 1983, 1984; Haubrick and Assmann, 2006). Earlier Sasse et al. (1995) also reported the ability of 24-epiBL (24-epiBL) to stimulate seed germination of Eucalyptus camaldulensis under saline conditions. Similarly Anuradha and Rao (2001, 2003) reported that 24-epiBL and 28-homoBL alleviated the inhibition of germination and seedling growth; and prevented the photosynthetic pigment loss in rice induced by salinity. Our earlier studies had also shown that 24-epiBL treatments (presowing) improved the shoot emergence and plant biomass production in Brassica juncea seedlings and plants under heavy metal stress (Cu, Zn, Mn, Co and Ni). 24-EpiBL has also been found to reduce the heavy metal uptake and accumulation in B. juncea seedlings and plants. The mechanism involved for reducing toxicity may be the chelation of the metal ions by the ligands. Such ligands include organic acids, amino acids, peptides or polypeptides (Sharma and Bhardwaj, 2006, 2007).
Improvement of seedling growth may also be due to increase in protein content and reduction of lipid peroxidation. Earlier studies also report that 28-homoBL and 24-epiBL induced denovo polypeptide synthesis in wheat leaves under heat shock stress (Kulaeva et al., 1991). Similarly Bajguz (2000) also found that BRs increased DNA, RNA and protein contents of Chlorella vulgaris as the number of cells increased in medium. Sam et al. (2001) studied the ultra structure of tomato leaf discs treated with BB6 (brassinosteroids analogue with spirostanic structure as active ingredient) under high temperature. BB6 treatment increased the rate of production of heat shock proteins, which protected mRNA from stress-induced degradation which further increased the protein production. As membrane destruction results from ROS induced oxidative damage (McCord, 2000), the 28-homoBL treated seedlings might be scavenging ROS more effectively than the seedlings treated with metal alone. The observations are in consistence with the results of Ozdemir et al. (2004), who found that lipid peroxidation level induced by NaCl was significantly lower in rice seedlings when treated with 24-epiBL as compared to control.
Environmental stresses often cause membrane damage, decrease hydrolytic enzyme activity and increased lipid peroxidation level. It may stimulate formation of ROS, such as H2O2, O2• ¯ and OH• radicals. To neutralize the toxicity of ROS, plants have enzymatic (e.g. superoxide dismutase, guaiacol peroxidase, ascorbate peroxidase, catalase and glutathione reductase) and non-enzymatic (e.g. ascorbate, glutathione, tocopherols and proline) defence system to operate (Mittler, 2002; Schutzendubel and Polle, 2002; Arora et al., 2002). Among ROS, superoxide radical (O2•¯) is dismutated by SOD into H2O2 and is further scavenged by CAT and various peroxidases. APOX and GR also play a key role by reducing H2O2 to water through the ascorbate-glutathione cycle (Noctor and Foyer, 1998). It is widely accepted that ROS are responsible for various stress-induced damages to macromolecules and ultimately to cellular structures (Halliwell and Gutteridge, 1999). Consequently, the role of antioxidative enzymes, such as SOD, CAT, POD, GR and APOX become very important. The level of CAT, POD, APOX and GR is increased by the application of 28-homoBL to overcome the stress generated by Ni and to boost the resistance capacity of plants (Fig. 3). This data is consistent with the earlier studies that exogenous BRs treatment is effective in stressful rather than in optimal conditions (Sasse, 1997). The effect of 24-epiBL and MH5 (polyhydroxylated spirostanic analogue of brassinosteroids) was analyzed by Mazorra et al. (2002) on catalase, peroxidase and superoxide dismutase activity in tomato leaf discs at 25-40°C. Brs altered the activity of these enzymes, suggesting a possible role of 24-epiBL and MH5 in the reduction of cell damage produced by heat stress. Tomato plants treated with 24-epiBL were more tolerant to high temperature than untreated plants. Further Nunez et al. (2003) also noted higher activity of antioxidative enzymes in rice, grown under salinity and supplemented with BRs. 28-homoBL also ameliorated the cadmium toxicity in Brassica juncea plants by increasing the activities of peroxidase, catalase and superoxide dismutase (Hayat et al., 2007). So the difference in alteration of antioxidative enzyme activities may suggest that 28-homoBL treated seedlings were less affected by Ni than the untreated seedlings.
CONCLUSION
The present study suggests that though Ni is essential for normal plant growth and metabolism but elevated concentration may results in growth inhibition and toxicity symptoms. However, 28-homoBL overcomes the inhibitory effects of Ni stress. Previous studies on stress protective properties of Brs indicate that exogenous brassinosteroids can act efficiently in plants as immuno-modulators when applied at appropriate dose and at the correct stage of plant development. The present piece of work further strengthens the antistress activity of BRs and this makes BRs a suitable candidate for their application in agriculture, especially under stressed conditions.
REFERENCES
- Anuradha, S. and S.S.R. Rao, 2001. Effect of brassinosteroids on salinity stress induced inhibition of germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul., 33: 151-153.
CrossRefDirect Link - Anuradha, S. and S.R. Rao, 2003. Application of brassinosteroids to rice seeds (Oryza sativa L.) reduced the impact of salt stress on growth, prevented photosynthetic pigment loss and increased nitrate reductase activity. Plant Growth Regul., 40: 29-32.
Direct Link - Arora, A., R.K. Sairam and G.C. Srivastava, 2002. Oxidative stress and antioxidative system in plants. Curr. Sci., 82: 1227-1238.
Direct Link - Bai, C., C.C. Reilly and B.W. Wood, 2006. Nickel deficiency disrupts metabolism of ureides, amino acids and organic acids of young pecan foliage. Plant Physiol., 140: 433-443.
CrossRefPubMedDirect Link - Bajguz, A., 2000. Effect of brassinosteroids on nucleic acids and protein content in cultured cells of Chlorella vulgaris. Plant Physiol. Biochem., 38: 209-215.
CrossRefDirect Link - Brown, P.H., R.M. Welch and E.E. Cary, 1987. Nickel: A micronutrient essential for higher plants. Plant Physiol., 85: 801-803.
Direct Link - Cao, S., Q. Xu, Y. Cao, K. Qian and K. An et al., 2005. Loss-of-function mutations in DET2 gene lead to an enhanced resistance to oxidative stress in Arabidopsis. Physiol. Plant, 123: 57-66.
CrossRefDirect Link - Clouse, S.D. and J.M. Sasse, 1998. Brassinosteroids: Essential regulators of plant growth and development. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49: 427-451.
CrossRefDirect Link - Costa, M., K. Salnikow, J.E. Sutherland, L. Broday, W. Peng, Q. Zhang and T. Kluz, 2002. The role of oxidative stress in nickel and chromate genotoxicity. Mol. Cell. Biochem., 234: 265-275.
Direct Link - Dalton, D.A., S.A. Russell and H.J. Evans, 1988. Nickel as a micronutrient element for plants. Biofactors, 1: 11-16.
PubMed - Dhaubhadel, S., S. Chaudhary, K.F. Dobinsen and P. Krishna, 1999. Treatment with 24-epibrassinolide, a brassinosteroid, increases the basic thermotolerance of Brassica napus and tomato seedlings. Plant Mol. Biol., 42: 333-342.
Direct Link - Dhaubhadel, S., K.S. Browning, D.R. Gallie and P. Krishna, 2002. Brassinosteroid functions to protect the translational machinery and heat shock protein synthesis following thermal stress. Plant J., 29: 681-691.
PubMedDirect Link - Haubrick, L.L. and S.M. Assmann, 2006. Brassinosteroids and plant function: Some clues, more puzzles. Plant Cell Environ., 29: 446-457.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Khripach, V.A., V.N. Zhabinskii and A.E. de Groot, 2000. Twenty years of brassinosteroids: Steroidal hormones warrant better crops for the XXI century. Ann. Bot., 86: 441-447.
CrossRefDirect Link - Kono, Y., 1978. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys., 186: 189-195.
CrossRefPubMedDirect Link - Krishna, P., 2003. Brassinosteroid-mediated stress responses. J. Plant Growth Regul., 22: 289-297.
CrossRefDirect Link - Kulaeva, O.N., E.A. Burkhanova, A.B. Fedina, V.A. Khokhlova, G.A. Bokebayeva, H.M. Vorbradt and G. Adam, 1991. Effect of Brassinosterods on Protein Synthesis and Plant-Cell Ultrastructure Under Stress Conditions. In: Brassinosteroids-Chemistry, Bioactivity and Applications, Culter, H.G., T. Yokota and G. Adam (Eds.), ACS Symposium Series, American Chemical Society, Washington, pp: 141-155.
- Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Lu, H., X. Shi, M. Costa and C. Huang, 2005. Carcinogenic effect of nickel compounds. Mol. Cell. Biochem., 279: 45-67.
CrossRefPubMedDirect Link - Mazorra, L.M., M. Nunez, M. Hechavarria, F. Coll and M.J. Sanchez-Blanco, 2002. Infulence of brassinosteroids on antioxidant enzymes activity in tomato under different temperatures. Biol. Plant, 45: 593-596.
Direct Link - McCord, J.M., 2000. The evolution of free radicals and oxidative stress. Am. J. Med., 108: 652-659.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Nunez, M., P. Mazzafera, L.M. Mazorra, W.J. Siqueira and M.A.T. Zullo, 2003. Influence of brassinosteroid analogue on antioxidant enzymes in rice grown in culuture medium with NaCl. Biol. Plant., 47: 67-70.
CrossRefDirect Link - Ozdemir, F., M. Bor, T. Demiral and I. Turkan, 2004. Effects of 24-epiBL on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul., 42: 203-211.
CrossRefDirect Link - Rahman, H., S. Sabreen, S. Alam and S. Kawai, 2005. Effects of nickel on growth and composition of metal micronutrients in barley plants grown in nutrient solution. J. Plant Nutr., 28: 393-404.
CrossRefDirect Link - Rao, S.S.R., B.V. Vardhini, E. Sujatha and S. Anuradha, 2002. Brassinosteriods. A new class of phytohormones. Curr. Sci., 82: 1239-1245.
Direct Link - Salin, M.L., 1988. Toxic oxygen species and protective systems of the chloroplast. Physiol. Plant., 72: 681-689.
CrossRefDirect Link - Sam, O., M.C. Nunez, M.C. Ruiz-Sanchez, J. Dell`Amico, V. Falcon, M.C. Dela Rosa and J. Seoane, 2001. Effect of a brassinosteroid analogue and high temperature stress on leaf ultrastructure of Lycopersicon esculentum. Biol. Plant, 44: 213-218.
Direct Link - Sasse, J.M., 1997. Recent progress in brassinosteroid research. Physiol. Plant., 100: 697-701.
Direct Link - Schutzendubel, A. and A. Polle, 2002. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot., 53: 1351-1365.
CrossRefPubMedDirect Link - Sharma, P. and R. Bhardwaj, 2007. Effect of 24-Epibrassinolide on seed germination, seedling growth and heavy metals uptake in Brassica juncea L. Gen. Applied Plant Physiol., 33: 59-73.
Direct Link - Sharma, P. and R. Bhardwaj, 2007. Effects of 24-epibrassinolide on growth and metal uptake in Brassica juncea L. under copper metal stress. Acta Physiologiae Plantarum, 29: 259-263.
CrossRefDirect Link - Wachsman, M.B., E.M. Lopez, J.A. Ramirez, L.R. Galagovsky and C.E. Coto, 2000. Antiviral effect of brassinosteroid against herpes virus and arena viruses. Anti. Chem. Chemother., 11: 71-77.
PubMedDirect Link - Welch, R.M. and L. Shuman, 1995. Micronutrient nutrition of plants. Crit. Rev. Plant Sci., 14: 49-82.
CrossRefDirect Link - Mandava, N.B., 1988. Plant growth-promoting brassinosteroids. Annu. Rev. Plant Physiol. Plant Mol. Biol., 39: 23-52.
CrossRefDirect Link