
Mehmet Ali Avci
Not Available
Ercan Ceyhan
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 1 | Page No.: 1-4







ABSTRACT
A line x tester analysis was carried out in pea to determine the genetic interaction in the expression of various pod characters including pod yield. Seven varieties were selected, four of them were designated as lines Sprinter, Manuel, Carina and Bolero and three of them were called as testers, B1, B6 and B12 line. Combining ability studies were conducted on these parents along with their F1 hybrids for the variables pod yield, hundred seed weight, pods per plant, pod width, pod length and seeds per pod. While Sprinter, Manuel and B6 were best general combiners among the parents, Sprinter x B12, Manuel x B6 and Carina x B1 were the best crosses for pod yield. Correlation studies showed that the pod yield was significant positive correlated with pods per plant and hundred seed weight. The highest direct effect was exhibited by pods per plant, indirect effects, especially through the seeds per pod in pea.
PDF Abstract XML References Citation
How to cite this article
Mehmet Ali Avci and Ercan Ceyhan, 2006. Correlations and Genetic Analysis of Pod Characteristics in Pea (Pisum sativum L.). Asian Journal of Plant Sciences, 5: 1-4.
DOI: 10.3923/ajps.2006.1.4
URL: https://scialert.net/abstract/?doi=ajps.2006.1.4
DOI: 10.3923/ajps.2006.1.4
URL: https://scialert.net/abstract/?doi=ajps.2006.1.4
INTRODUCTION
Pea (Pisum sativum L.) also referred as dry pea to distinguish it from the vegetable type, is one of the most important grain legumes occupying an area of more than 7 m ha in the World. The significant pea growing nations are China, France, Australia and India. The total pea sown areas, seed yield and production in Turkey is 110,000 ha, 2.3 t ha-1 and 250 000 t, respectively[1].
Breeding for superior varieties requires selection of parents capable of transmitting their desirable qualities. A rational approach for breeding is to select parents based on their combining ability rather than on visual observation of their traits. Studies on combining ability provide useful information for the selection of parents to include in a hybridization programme by defining the parent of gene action in the expression of quantitative traits[2,3]. General combining ability provides an indication of importance of genes which have largely additive effects while specific combining ability indicates the importance of non-additive effects.
One of the problems in breeding plant genotypes is knowledge of relationships between pod yield and pod characters. To increase the pod yield, study of direct and indirect effects of pod traits provides the basis for its successful breeding programme and hence the problem of pod yield increase can be more effectively tackled on the basis of performance of pod yield traits and selection for closely related characters[4]. Ceyhan[5] reported high genotypic and phenotypic variances for pod yield, pods per plant and hundred seed weight. He also reported positive correlation of all the pod traits with pod yield. Use of simple correlation analysis could not fully explain the relationships among the characters. Therefore, the path coefficient analysis has been used by many researches for a more and complete determination of impact of independent variable on depend one. The path coefficient analysis helps the breeder to explain direct and indirect effects and hence has extensively been used in breeding work in different crops species by various researchers[6-8]. The objectives of this study were to determine the general and specific ability, relationship among pod yield and pod traits in pea as a good source of plant protein.
MATERIALS AND METHODS
Twelve F1 progenies obtained from a line x tester mating design were evaluated along with the seven parents in a randomized complete block design with three replications, to understand the genetics basis and combining ability of some important pod quality traits. The six varieties selected were all dwarf types, including a wide range of pod traits. B1, B6 and B12 lines were well adapted to local conditions. Carina, Sprinter, Cosmos and Manuel demonstrated that high yields can be obtained by growing spring in Central Anatolian.
Seeds of F1 hybrids and parental lines were sown in a randomized complete block design with three replications on 18 March 2002. Each plot consisted of 12 F1 or parent plants on a single 2 m row which were 50 cm apart. Plant spacing was 10 cm apart. Total annual precipitation was 159.5 mm, which was more than in long years (139.3 mm) of the site. The experimental crops were irrigated two times (at the initiation of flowering and at the pod filling stage). Weeds were removed manually. The soil at the region had a pH = 8.03 and soil phosphorous, potassium, iron, zinc, calcium and organic matter were 55.9, 17.9 kg ha-1, 14.74, 0.32 ppm and 37.6, 2.25%, respectively. Plants were grown without fertilization and harvested on 10 July 2002.
Pod yield (g) data were collected at the dry pod yield on five plants in the each plot. Pods number per plant was measured on 5 randomly plants per plot taken at maturity. Pod length (mm) were measured to closest centimeter using a meter scale. Pod width (mm) was measured from the widest point of the leaflet using a vernier caliper. Number of seeds per pod was obtained from number seeds of a pod. Hundred seed weight (g) was expressed as a mean of four separate 20 seed weights, multiplied by 5.
Regarding the statistical analysis, data recorded on parents and the F1 hybrids were analyzed together as suggested by Singh and Chaudhary[9]. The combining ability analysis was done following Kempthonre[10]. Narrow sense heritabilities were calculated for each character by using the Falconer’s[11] methods. Correlation coefficients and the path analysis was conducted following the procedure developed by Wright[12] and applied by Dewey and Lu[13].
RESULTS AND DISCUSSION
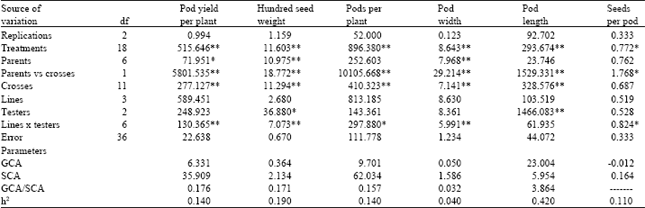
Mean sum of squares of parents were highly significant for almost all characters except for pod length and seeds per pod. Variation due to crosses showed significant differences for all characters. Parent x crosses were significant for all traits. Variation due to testers showed significant difference for hundred seed weight and pod length while lines no differed for all traits. The interaction between line and tester was significant all characters except pod length (Table 1).
Table 2 also reveals the fact that the ratio of variance of GCA and SCA was much less than unity for all characters which indicate the predominant role of non-additive gene action in the inheritance of most of the traits in pea. Sharma et al.[3] and Ceyhan[5] observed nature of dominance and non-additive genes for pod yield and pod characters. Kumar et al.[2] showed additive gene action in the inheritance of pod length, while the other characteristics investigated controlled mainly by non-additive gene action. This study confirms the latter view.
Narrow sense heritability values varied among traits (Table 1). Of all traits studied, pod length had the greatest heritability for narrow sense heritability. The narrow sense heritabilities of all studied traits except pod length were relatively high suggesting that selection of these traits in selection of these traits in early generations would not be possible. Similar conclusions were obtained Kumar et al.[2], Sharma et al.[3] and Ceyhan[5].
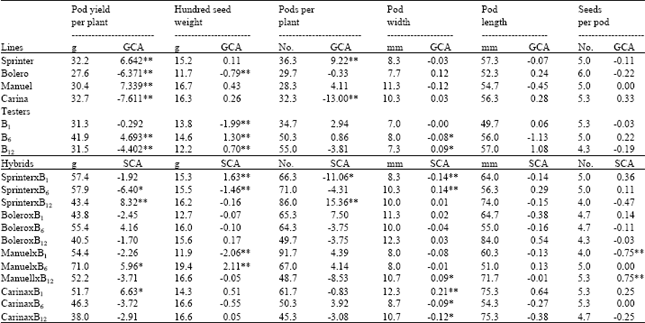
Hybrid performance was generally better than parental performance for all characters (Table 2). This result was in agreement with Kumar et al.[2], Ceyhan and Önder[4], Ceyhan[5] Amurrio et al.[14] and Santalla et al.[15].
The estimated GCA effects of parents (Table 2) revealed considerable differences among the parents. The parents that proved to be good general combiners on the basis of their desirable GCA effects were Sprinter for pod yield and pods per plant and Manuel for pod yield. B6 exhibited positively significant GCA effects for pod yield and hundred seed weight. B12 expressed significant GCA in hundred seed weight and pod.
| Table 1: | Analysis of variance for pod yield and pod traits of the parents and their F1 generation of pea |
 | |
| *: p<0.05, **: p<0.01 | |
| Table 2: | Mean pod yield and pod traits and general combining ability and specific combining in pea |
 | |
| *: p<0.05, **: p<0.01 | |
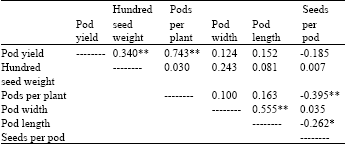
| Table 3: | Correlations coefficients among pod yield and pod traits in pea crosses |
 | |
| *: p<0.05, **: p<0.01 | |
However, among the parents the highest positive effect for pod yield was exhibited by Sprinter, Manuel and B6 hence they should be considered as the best female and male combiners.
The SCA effects (Table 2) clearly revealed that it would not be possible to isolate crosses where all traits are in the most desirable combinations. Also, it appeared that desirable SCA effects of the cross combinations were not necessarily depended on the level of GCA effects of parents involved. The SCA estimates for pod length showed no combination in desirable direction. However the SCA estimates of the crosses Sprinter x B12, Carina x B1 and Manuel x B6 were highly significant for pod yield. Similarly, Sprinter x B1 and Manuel x B6 showed significant positive SCA effects for hundred seed weight, Sprinter x B12, for pods per plant, Sprinter x B6, Manuel x B12 and Carina x B1 for pod width while Manuel x B12 had significant positive SCA effects for seeds per pod. This suggests that on the basis of general combining ability studies it would be difficult to make definite breeding plans as the high pod yielding combination was obtained from parents which did not show significant GCA effect in desirable direction. However, the high SCA effects of the crosses Sprinter x B12, Carina x B1 and Manuel x B6 further confirm the predominance of non-additive gene actions in pea. Hence, it is suggested that in pea emphasis should be given to specific crosses followed by selection in progenies rather than pursuing GCA by mass selection. General and specific combining ability has previously been shown in pea to be the major contributing factor for pod yield[2-5].
Correlation coefficients were determined between pod yield and other variables. Pod yield was significantly positive correlated with hundred seed weight and seeds per pod (Table 3). Relationships between pod length and pod width were significant positively correlated. Seeds per pod correlated significant negatively with pods per plant and pod length. Ceyhan[5] found positive correlation between pod yield with hundred seed weight, pod width, pod length, pods per plant and seeds per pod in pea. These results shown that, for high pod yield should be moderately with hundred seeds weight and seeds per pod.
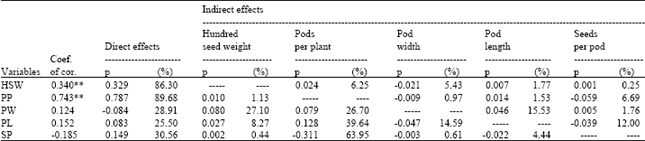
The highest direct positive effects on pod yield were exhibited by pods per plant Relation between pod yield and pods per plant was positive and significant, with a direct effect of 89.68% and indirect effects of 6.69%, especially through the seeds per pod (Table 4). The direct effects of hundred seed weight on pod yield were also positive and significant. These relations for hybrids were further studied using breeding programs.
| Table 4: | Path coefficient analysis between pod yield and other variables examined among different pea genotypes |
 | |
| *: p<0.05, **: p<0.01, HSW: Hundred seed weight, PP: Pods per plant, PW: Pod width, PL: Pod length, SP: Seeds per pod | |
Selection in a breeding program based pods per plant was 89.68% as effective as selection for pod yield directly. When selection for pod yield was based alone on hundred seed weight, genetic advance was 86.30%. Pod length had lowest direct effect on pod yield.
By comparing the correlation coefficients values of six independent variables against the pod yield, significant differences became evident. pods per plant and hundred seed weight had highly significant association with pod yield. Pods per plant and hundred seed weight also were only characters that exhibited the highest direct effect on pod yield. Therefore, both these traits seem to be a good selection criteria to improve pod yield in pea.
REFERENCES
- Ali, N., F. Javidfar, J.Y. Elmira and M.Y. Mirza, 2003. Relationship among yield components and selection criteria for yield improvement in winter rapeseed (Brassica napus L.). Pak. J. Bot., 35: 167-174.
Direct Link - Dewey, D.R. and K.H. Lu, 1959. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J., 51: 515-518.
CrossRefDirect Link