
C.G. Ipsilandis
Not Available
B.N. Vafias
Not Available
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 1 | Page No.: 31-39







ABSTRACT
Breeding in maize (Zea mays L.) succeeded in improving tolerance to stresses and responsiveness to inputs, but failed to improve the potential yield per plant. Modern hybrids are highly productive, but their yield per unit area is dependent on high plant densities. Results of present study verified that modern hybrids require a narrow range of densities to give their maximum yield per unit area, since plant density affected significantly all kind of genetic materials. Results also showed that high densities are associated with high plant-to-plant variability and barrenness, affecting negatively the stability of hybrid performance. F2 and F1+F2 (mixture) genetic materials showed increasing yields, as plant density was increasing. This was due to high competition and the presence of heterotic plants. These findings pointed out the necessity for the development of maize hybrids with more stable performance across growing seasons. Density-independent hybrids can be effectively attained through selection in the absence of competition, aiming to improve the potential yield per plant. Very low densities, so that any plant-to-plant interference for growth resources use is eliminated, maximize mean yield per plant differentiation and optimise the phenotypic-genotypic correspondence, facilitating the identification of the superior entries. Evaluation under high plant densities may distinguish between different genetic materials, based on F2 yielding performance, to be used as starting materials in breeding programs. Even though the main goal is breeding for density-independent maize hybrids, since modern maize hybrids depend their yielding performance on higher plant densities, breeding for density tolerance may easily contribute to higher field yields.
PDF Abstract XML References Citation
How to cite this article
C.G. Ipsilandis and B.N. Vafias, 2005. Plant Density Effects on Grain Yield per Plant in Maize: Breeding Implications. Asian Journal of Plant Sciences, 4: 31-39.
DOI: 10.3923/ajps.2005.31.39
URL: https://scialert.net/abstract/?doi=ajps.2005.31.39
DOI: 10.3923/ajps.2005.31.39
URL: https://scialert.net/abstract/?doi=ajps.2005.31.39
INTRODUCTION
Since 1960 breeding in maize has contributed to improvement of hybrid yield per unit area at the annual rate of 1.1 quintals ha-1[1]. When hybrid grain yield was estimated at the typical plant density of 7.9 plants m-2. However, during the past 70 years, breeding had no effect on hybrid yield per plant improvement, as it was estimated under low stress conditions (1 plant m-2)[2]. This finding was of paramount importance showing that, in fact, higher grain yield productivity of modern maize hybrids resulted indirectly by improving a range of traits associated with tolerance to various biotic and abiotic stresses and by improving the efficiency of capture and use of resources[3-5]. The differential response to stress between older and newer hybrids has been shown for low night temperature during the grain-filling period[6], low soil moisture in the field[7,8], low soil N[9], weed interference[10] and high plant population density[11,12]. As a consequence of these findings and for further genetic improvement of maize hybrids in the future, Duvick[2] suggested that the best way to effect future gains in yielding ability may be to make further improvements in tolerance to high plant densities, in combination with improvements in potential yield per plant under low stress environments. Fasoula and Fasoula[13-15] emphasised the importance of low stress conditions (i.e., very low plant density, so that competition among plants is avoided) in optimising the effectiveness of selection for improved potential yield per plant, tolerance to stresses and responsiveness to inputs. The researchers stated that these three parameters constitute the key in the development of cultivars characterised by high and stable yield per unit area. The objective of the present study was to assess the plant density effects on yield per unit area of maize hybrids (that is the hybrid dependence on plant density) and the competition impact on plant-to-plant variability, as well as the following consequences on maize breeding strategy.
MATERIALS AND METHODS
From two commercial single-cross maize hybrids (named Rio Grande and Costanza) three kinds of genetic material (F1, F2 and F1+F2 mechanical mixture at the 1:1 weight/weight rate) were used in the study. They particular hybrids were chosen because they had been of the most widely used maize hybrids in Greece during the last decade, due to their high and stable productivity. During 1999 growing season the genetic materials were evaluated in two adjacent experiments (one for each hybrid) arranged in Split-plot Complete Block Design, under five different plant densities and four replications per density (with the subplots consisting of plant densities), conducted in Technological Education Institute farm of Larissa, Greece.
As the major target of the study was the plant density impact on hybrid performance and not the two hybrids to be compared, it was preferred each hybrid to be evaluated in different experiments. Analysis of Variance was based both on the split-plot model and the plain RCB design. The unified RCB design analysis was chosen too, since we were interested in either the plant density impact on entry performance or the differences between the three kinds of genetic material of each hybrid (the density impact on entry differentiation). Plots consisted of two rows, 500 cm long and 75 cm wide. Distances between plants within rows were 30, 25, 20, 15 and 10 cm to obtain the respective densities of 44,444, 53,333, 66,666, 88,888 and 133,333 plants/ha. Plots were overplanted and thinned to desired stand at the seedling stage. Nitrogen and P fertiliser were applied at the rate of 120 and 60 kg ha-1, respectively at planting stage, while additional N (100 kg ha-1) was applied seven weeks after planting. Complete weed control was obtained by hand weeding. Trials were regularly irrigated to avoid drought stress.
Plants were harvested individually and after adjusting to 15.5% grain moisture, the mean yields per plant and entry ( ![]() ) were calculated. Plant-to-plant variability of yield per plant for a particular entry was estimated by the coefficient of variation (CV) of individuals (CV=σp /
) were calculated. Plant-to-plant variability of yield per plant for a particular entry was estimated by the coefficient of variation (CV) of individuals (CV=σp /![]() ), on the basis of mean value across replicates and the Q-Q plot method[16] verified the appropriateness of analysis of variance for CV values (p<0.001). According to Tollenaar and Wu[5], Fasoula and Fasoula[13-15], plant-to-plant variability consists of a measure of stand uniformity and therefore, it could be used as an index of stability. Present goal was to investigate how stand uniformity is influenced by the plant density changes. Additionally, barrenness was evaluated, as the percentage of plants per entry found to be unable to form any ear. Regression analysis was conducted for yield per unit area, yield per plant differentiation, CV values and percentage of barren plants response to plant density changes. Analysis of variance for quadratic regression[17], based on single plot values, substantiated the quadratic model in most cases, whereas in eight out of twenty four regressions the departure from linearity was not significant and was used instead (Fig. 1-6).
), on the basis of mean value across replicates and the Q-Q plot method[16] verified the appropriateness of analysis of variance for CV values (p<0.001). According to Tollenaar and Wu[5], Fasoula and Fasoula[13-15], plant-to-plant variability consists of a measure of stand uniformity and therefore, it could be used as an index of stability. Present goal was to investigate how stand uniformity is influenced by the plant density changes. Additionally, barrenness was evaluated, as the percentage of plants per entry found to be unable to form any ear. Regression analysis was conducted for yield per unit area, yield per plant differentiation, CV values and percentage of barren plants response to plant density changes. Analysis of variance for quadratic regression[17], based on single plot values, substantiated the quadratic model in most cases, whereas in eight out of twenty four regressions the departure from linearity was not significant and was used instead (Fig. 1-6).
The final (main) experimental design choice was based on preliminary data from a previously used experimental design. Preliminary experimentation involved a spilt-plot design similar to the one described above. The two above-mentioned hybrids were used and the three kinds of materials were formed.
| Table 1: | Mean yield per plant in grams (0), coefficient of variation of individual plant yields (CV) and percentage barren plants at five plant densities of the F1, F1+F2 mixture and F2 of the hybrids Rio Grande and Costanza (main experiment) |
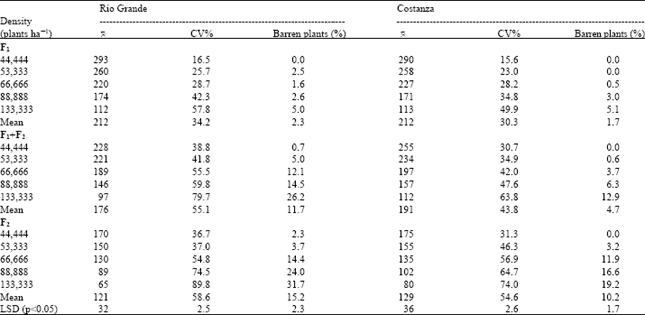 | |
| Table 2: | Mean yield per plant in grams (0), coefficient of variation of individual plant yields (CV) and percentage barren plants at four plant densities of the F1, F1+F2 mixture and F2 of the hybrids Rio Grande and Costanza (preliminary data). |
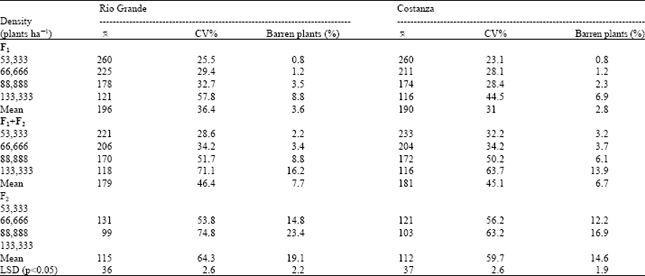 | |
The only difference was the use of four plant densities (53,333, 66,666, 88,888 and 133,333 plants ha-1), plus the lack of some data for F2 generation. Main experimental data are presented in Table 1 and preliminary data are presented in Table 2 to indicate the appropriateness of the experimentation and similarity of data. This preliminary experimentation was considered necessary to define the proper main experimental design and to obtain more data for comparisons in different years. Discussion of data was based on the main experiment (with five levels of the plant density factor).
F1, F2 and their mixture were used to compare the influence of plant density on genetic materials with different level of heterozygosity, gene action and uniformity. F1 is considered uniform, whereas F2 and the mixture are non-uniform genetic materials and this implies competition effects according to Fasoulas[18]. Uniformity is a significant parameter for breeding strategies and agronomy performance. In this study agronomy implications were ignored and breeding strategies were analysed.
RESULTS
Data from either F1 or F2, as well as of their mixture, of both hybrids Rio Grande and Costanza, show that yield per plant decreased with increasing plant density (Table 1 and 2), an expected impact due to increased competition among plants at higher densities[19]. After adjustment of yield per plant to yield per unit area, an important consequence arises. Yield potential per unit area of the two hybrids was found to be dependent on plant density, as Fig. 1 (main experiment) and Fig. 2 (preliminary data) demonstrate.
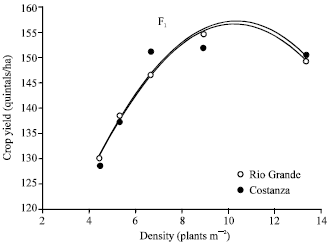 | |
| Fig. 1: | Relationship between plant density and maize yield per unit area found curvilinear. The equation of line is: y=-0.76x2+15.5x+76.6 for Rio Grande (R2=0.999) and y=-0.77x2+15.8x+75.3 for Costanza (R2=0.92). The two curves reveal hybrid yield dependence on plant density in maize (main experiment) |
In general, hybrid Costanza, showed more stable yielding performance across densities (level between 66,666 and 133,333 plants ha-1). Graphs of the yield per unit area data indicate that the yields of the two hybrids tended to level off at the same density of 10 plants m-2, while any plant population, either lower or higher than 100,000 plants ha-1, results in grain yield loss.
Even though the F1 yielding performance across densities was similar for the two hybrids, beginning from the same level of yield and ending at the same level of yield (Fig. 1), the F2 yielding performance behaved differently (Fig. 3).
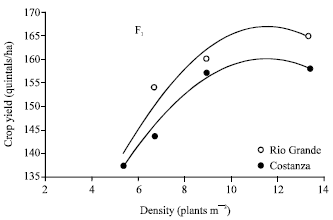 | |
| Fig. 2: | Relationship between plant density and maize yield per unit area (preliminary data), indicating hybrid yield dependence on plant density |
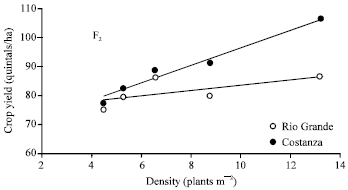 | |
| Fig. 3: | Relationship between plant density and F2 maize yield per unit area (main experiment). The equation of line is: y=0.0869x+74.878 for Rio Grande (R2=0.4) and y=0.3031x+66.252 for Costanza (R2=0.953) |
The F2s of the two hybrids began from the same level of yield, but as plant density was increasing, the yielding performance of Costanza’s F2 was increasing at a higher rate than Rio Grande’s one.
Yield per plant differentiation between the three kinds of material was inversely associated with plant density, (Fig. 4). At the lower plant density of 4.4 plants/m2 the yield per plant differences between F 1 and F 1+F 2 mixtures were 65 g for Rio Grande and 35 g for Costanza, with the respective differences at the higher density of 13.3 plants m-2 being 15 and 1 g (Table 1). Yield per plant differences between F1 and F2 at the lower and the higher density respectively were 123 and 47 g for Rio Grande and 115 and 33 g for Costanza.
Coefficients of variation of individual plant yields were higher in F1+F2 mixtures and F2s, compared with the CV values in F1s (Table 1 and 2), an expected impact as the F1s are assumed to be genetically homogeneous (based on blue-print specifications). Regarding the plant density influence on stand uniformity, it was found that increased plant-to-plant variability accompanied increased plant density (Fig. 5). Compared with the CV values at the lower plant density of 4.4 plants m-2, CV values at the higher plant density of 13.3 plants m-2 were over threefold at the F1s and over twofold at F1+F2 mixtures and F2s (Table 1).
The effects of genetic homogeneity and plant density on the percentage of barren plants were similar with those observed in case of CV values. Compared with the average percentage of barren plants at the two homogeneous F1s, the corresponding values at F1+F2 mixtures and F2s were over four and six times higher, respectively (Table 1). Figure 6 show that barrenness increases as plant density increases.
DISCUSSION
Grain yield per unit area: The dependence of maize hybrid yield per unit area on plant density shown by the curvilinear response of Fig. 1, was also depicted by previously reported data[1,20-22]. The very high optimum plant density of 10 plants m-2 (Fig. 1), being in agreement with that found by Tetio-Kagho and Gardner[20], evidences previous reports that modern hybrids, compared with the older ones, require higher densities to give their maximum yield per unit area. Preliminary data exhibited an even higher optimum plant density (Fig. 2). Differences in grain yield between older and newer maize hybrids were shown to be a function of plant population density[11]. Breeding in maize succeeded in developing modern hybrids with highly improved yield per unit area, through improvement in tolerance to stresses, including high plant densities[3-5,8,23]. On the contrary, breeding had no contribution to hybrid yield per plant improvement[2]. The necessity of higher plant densities for optimal productivity of modern maize hybrids led that selection under higher plant densities was a means to improve grain yield of maize[24]. As a consequence of selection under high plant densities, with no effect on yield per plant improvement, modern hybrids were rendered highly dependent on optimum plant density even though they exhibit a higher optimum population density[15]. Farmers have always favoured cultivars which are characterised by stable performance across the growing seasons. From this point of view, density-independence should be considered in the development of maize hybrids, as a means of a stability parameter incorporation in modern hybrids, because, according to Fasoula and Fasoula[15], the density-dependent cultivars are accompanied by a number of disadvantages; i.e., increased density by cultivar interaction, seeding cost and increased risk in case of adverse conditions during the growing season.
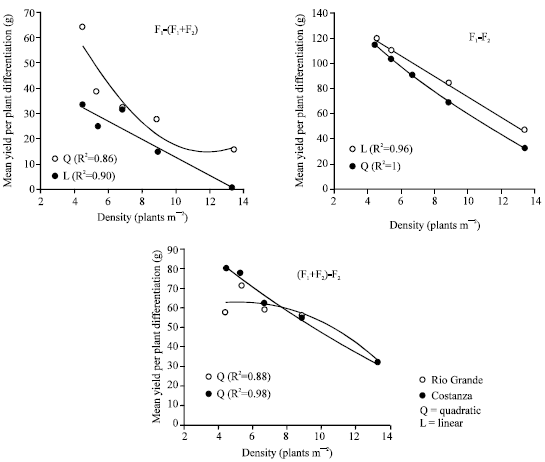 | |
| Fig. 4: | Relationship between the plant density and the mean yield per plant differentiation between F1, F1+F2 mixture and F2 of the maize hybrids Rio Grande and Constanza. In general, lower plant densities are associated with greater differentiation, facilitating the identification of superior entries |
Yield differentiation among entries: Inverse association between entries’ differentiation and plant density, similar with that of Fig. 4, showed data presented by Tokatlidis et al.[25], Fery and Janick[26], Traka-Mavrona[27] regarding maize hybrid yield, tomato (Lycopercicon esculentum Mill.) variety yield and carrot (Daucus carota L.) quality traits, respectively. The impact of plant density on yield differentiation should be considered in the degree of competition under which evaluation for selection of entries should be applied. The decreased differentiation with increasing plant density indicates that evaluation and selection in dense stand would impair response to selection. On the contrary, the increased differentiation at the lower densities would facilitate the identification of the superior entries. When plant density is very low, as to exclude plant-to-plant interference with the equal use of growth resources (i.e, in the absence of competition), the maximised phenotypic differentiation will maximise response to selection[13].
Stand uniformity: The plant density influence on plant-to-plant variability, either in homogeneous F1s, or in heterogeneous F1+F2 mixtures and F2s (Fig. 5), clearly demonstrate that lower densities contribute to greater stand uniformity. Increased plant-to-plant uniformity for yield per plant of maize hybrids, with decreasing the plant density was also reported by Tollenaar and Wu[5], Vafias et al.[28]. Similar impact of plant densities on plant-to-plant uniformity have been found for other maize hybrid traits like dry weight and plant size[29,30] and kernel number per ear[22]. Plant size variability is directly related to neighbourhood competition in which the growth of an individual depends on the number, size and proximity of neighbours[31]. Generally, there is a negative relationship between plant density and plant-to-plant uniformity[5,13,32]. Stand uniformity deserves consideration concerning the selection effectiveness in a breeding program and the productiveness of a field crop. Coefficient of variation represents two kinds of effects, as a consequence of genetic and environmental variance. Therefore, the increased CV values of a given entry at the higher densities result from higher environmental effects. This is clearly seen in case of the F1s of hybrids Rio Grande and Costanza, which are assumed to be monogenotypic, meaning that genetic variance had no effects in their CV values.
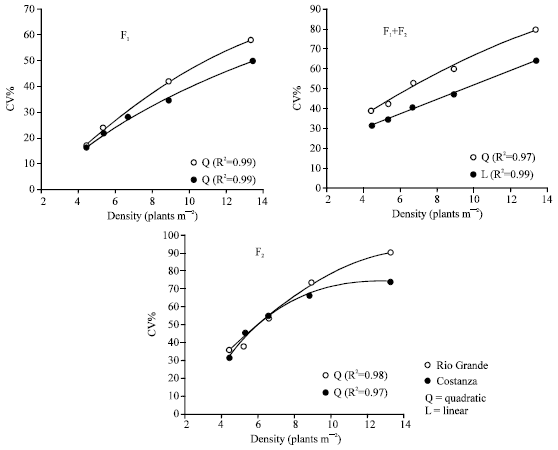 | |
| Fig. 5: | Relationship between the plant density and the CV of individual plant yield, in the F1, F1+F2 mixture and F2 of the maize hybrids Rio Grande and Constanza. Lower plant densities are associated with improved stand uniformity, an influence expected to be a crucial factor for high and stable yield per unit area |
Consequently, the increased CV values at the higher densities indicate that, as the plant density increases, phenotypic and genotypic correspondence decreases, rendering the individual plant selection more difficult and thus less effective. On the other hand, stand uniformity in a field crop plays an important role in resource use efficiency, adequate for maximum yield per unit area. Resource use efficiency is inversely related to plant-to-plant variability, because in non-uniform stands of field crops bigger or taller plants have a competitive advantage over the smaller or shorter ones[5]. There is a negative relationship between yield per unit area and plant-to-plant variability and thus stand uniformity is essential for maximum productivity[5,13,32,33].
Barrenness: Results showed that increased barrenness accompanies higher plant densities and similar were the findings by Vafias et al.[19,28] and Hashemi-Dezfouli and Herbert[21] who reported linear relationship between the number of barren plants and the plant population. The reduction in grain yield per plant and the increase in plant-to-plant variability with increased density may partly be attributed to an increase in barrenness. Therefore, barrenness seems to have a decisive contribution to reduced stand uniformity with increased density and to yield per unit area reduction at the very high densities. Although stand uniformity and stress tolerance are highly associated and easy to achieve, density-tolerant maize hybrids exhibit higher plant-to-plant variability at plant densities close to or higher than their optimum densities[5]. So, even in case of density-tolerant hybrids, greater variability at higher densities for time of pollen shed and silking could cause a greater asynchrony between them, that according to Hashemi-Dezfouli and Herbert[21] leads to increased number of barren plants and therefore, to the change in total grain yield.
Implications in breeding: Maize hybrid dependence on high densities and the enhanced plant-to-plant variability at high densities, render necessary to consider two important points: (a) the goal of developing density-independent hybrids in parallel with density-tolerant ones and (b) the conditions under which selection is optimised to be more effective in the aim to achieve this goal.
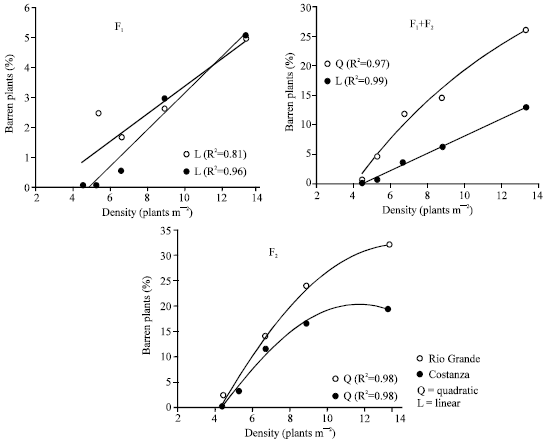 | |
| Fig. 6: | Relationship between the plant density and the percentage of barren plants, in the F1, F1+F2 mixture and F2 of the maize hybrids Rio Grande and Constanza. Lower densities result in reduced number of barren plants, followed by greater stand uniformity |
A cultivar has the potential to provide high and stable yield per unit area at a wide range of densities, if it has the genetic potential of high yield per plant at the lower densities, so that the lack of plants is compensated by individual plant potential for higher yield. The importance of potential yield per plant as a component of the yield per unit area potential and as it is expressed under very low stress conditions, has been emphasised by Fasoulas[18], Yan and Wallace[34], Duvick[2], Janick[32], Fasoula and Fasoula[13-15]. Breeding in maize failed to succeed in improving potential yield per plant because it has never focused on this target. Direct improvement of potential yield per plant can be achieved by selection at very low densities, so that competition among plants is minimised, because planting at very low density reduces stress to a minimum and allows maximum grain production per plant[13,15,18]. Selection in the absence of competition is also imposed by the decreased entry differentiation and the enhanced environmental effect on phenotypic expression under competition (Fig. 4 and 5). Strong plant-to-plant interference associated with growth under competition always reduces the range of genotypic expression, thus reducing the efficiency of phenotypic selection[15,18]. Additionally, a negative relationship between competitive ability and yield in pure stands has been widely reported[35,37].
Data presented by Tokatlidis et al.[25] showed that recycling of a hybrid, aiming to improve its potential yield per plant via self-pollination and selection at the very low density of 0.74 plants/m2 and on the basis of higher single plant yield and lower CV of individual plant yields, led to recycled hybrids which exhibited less density dependence than their original hybrid. In another recent study, Tokatlidis[38] emphasised the beneficial effect of the improved potential yield per plant of maize hybrids on their yield per unit area potential and the range of optimum plant density. Fasoula and Fasoula[15] thoroughly explained the causes of tolerance to higher and lower plant densities and the ways of overcoming the density barriers. This was made possible through partitioning of yield per unit area into three components, all assessed in the absence of competition. According to Fasoula and Fasoula[15], the role of the three components of the yield per unit area in developing density-independent cultivars is the following: The first component (i.e., yield per plant), determined by the progeny mean, extends the lower limit of the optimum plant density range. The second component (i.e., tolerance to stresses), determined by the progeny standardised mean (the reciprocal value of CV), extends the upper limit of the optimum plant density range. The third component (i.e., the responsiveness to inputs), determined by the progeny standardised selection differential, enables the exploitation of favourable environments. Tollenaar and Wu[5] claimed that increased stress tolerance, combined with increased stand uniformity under stress conditions, will probably continue to provide the highest potential for yield improvement in maize in next decades. Even though selection differential is maximised under higher plant densities in breeder’s field, strong plant-to-plant interference associated with growth under competition always reduces the range of genotypic expression, thus reducing the efficiency of phenotypic selection[15]. Even more, decreased differentiation with increasing plant density and reduced phenotypic and genotypic correspondence under competition, also reduces genotypic expression rendering individual plant selection insecure and less effective. The maximised selection differential under higher plant densities could distinguish among genotypes only in late breeding stages where phenotype represents better the genotype. However, the development of density-independent hybrids does not exclude density tolerance, which is incorporated into genotype by the above-mentioned second component of yield per unit area and easy to achieve.
Data of this study showed that the F2 yielding performance of the two maize hybrids used was different in high densities. F2 yield differences found greater than F1 ones for hybrids Costanza and Rio Grande, so that evaluation based on F2 yielding performance under high plant densities may distinguish between different genetic materials. Based on F2s’ performance many F1s could be evaluated and selected as starting populations for second cycle maize breeding schemes[39].
Maize breeders should consider the development of hybrids, which may give their maximum yield per unit area at a wide range of densities, in such a way that modern hybrids will be characterised by high yield per unit area and stable performance. This goal can be achieved through improvement of potential yield per plant and tolerance to stresses, so that the lower and the upper limit of the optimum plant density will be extended. Effective selection for higher yield per plant is feasible under non-competition conditions, so that the undesirable environmental effects on genotypic and phenotypic correspondence are eliminated. Under non-competition tolerance to stresses and consequently tolerance to high densities, may be attained by selection of entries, which exhibit the lowest sensitivity to environmental effects, i.e., those with the lowest CV of individual plant yields.
Improved yield per plant and tolerance to stresses, in combination with responsiveness to inputs may contribute to higher and stable yield per unit area, due to improved resource use efficiency. Since modern maize hybrids depend their yielding performance on higher plant densities, breeding for density tolerance or density independence may easily contribute to higher field yields. Evaluation under high plant densities may distinguish between different genetic materials, based on F2 yielding performance, to be used as starting populations in breeding programs.
ACKNOWLEDGMENTS
Authors are grateful to Professor C. Goulas of the University of Thessaly, Greece, co-writer in references Vafias et al.[19,28] for supporting experimentation and Dr. I.S. Tokatlidis of the University of Thrace, Greece, who supported part of the discussion.
REFERENCES
- Tollenaar, M. and J. Wu, 1999. Yield improvement in temperate maize is attributable to greater stress tolerance. Crop Sci., 39: 1597-1604.
CrossRefDirect Link - Dwyer, L.M. and M. Tollenaar, 1989. Genetic improvement in photosynthetic response of hybrid maize cultivars, 1959 to 1988. Can. J. Plant Sci., 69: 81-91.
Direct Link - Nissanka, S.P., M.A. Dixon and M. Tollenaar, 1997. Canopy gas exchange response to moisture stress in old and new maize hybrids. Crop. Sci., 37: 172-181.
Direct Link - McCullough, D.E., P.H. Cirardin, M. Mihajlovic, A. Aguilera and M. Tollenaar, 1994. Influence of N supply on development and dry matter accumulation of an old and a new maize hybrid. Can. J. Plant Sci., 74: 471-477.
Direct Link - Tollenaar, M., A. Aguilera and S.P. Nissanka, 1997. Grain yield is reduced more by weed interference in an old than in a new maize hybrid. Agron. J., 89: 239-246.
CrossRefDirect Link - Duvick, D.N., 1984. Genetic Contributions to Yield Gains of U.S. Hybrid Maize, 1930 to 1980. In: Genetic Contributions to Yield Gains of Five Major Crop Plants, Fehr, W.R. (Ed.). ASA and CSSA, Madison, WI., pp: 15-47.
CrossRefDirect Link - Fasoula, D.A. and V.A. Fasoula, 1997. Competitive ability and plant breeding. Plant Breed. Rev., 14: 89-138.
Direct Link - Fasoula, D.A. and V.A. Fasoula, 1997. Gene action and plant breeding. Plant Breed. Rev., 15: 315-374.
Direct Link - Fasoula, V.A. and D.A. Fasoula, 1999. Honeycomb breeding: Principles and applications. Plant Breed. Rev., 18: 177-250.
Direct Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Tetio-Kagho, F. and F.P. Gardner, 1988. Response of maize to plant population density. II. Reproductive development, yield and yield adjustments. Agron. J., 80: 935-940.
CrossRef - Hashemi-Dezfouli, A. and S.J. Herbert, 1992. Intensifying plant density response of corn with artificial shade. Agron. J., 84: 547-551.
CrossRefDirect Link - Troyer, A.F. and R.W. Rosenbrook, 1983. Utility of higher plant densities for corn performance testing. Crop Sci., 23: 863-867.
Direct Link - Tokatlidis, I.S., M. Koutsika-Sotiriou and A.C. Fasoulas, 2001. The development of density-independent hybrids in maize. Maydica, 46: 21-25.
Direct Link - Edmeades, G.O. and T.B. Daynard, 1979. The relationship between final yield and photosynthesis at flowering in individual maize plants. Can. J. Plant Sci., 59: 585-601.
Direct Link - Bonan, G.B., 1991. Density effects on the size of annual plant populations: An indication of neighbourhood competition. Ann. Bot., 68: 341-347.
Direct Link - Kenneth, A.G. and R.S. Helms, 1996. Seeding rate effects on rough rice yield, head rice and total milled rice. Agron. J., 88: 82-84.
Direct Link - Yan, W. and D.H. Wallace, 1995. Breeding for negatively associated traits. Plant Breed. Rev., 13: 141-177.
CrossRefDirect Link - Thomas, J.B., G.B. Schaalje and M.N. Grant, 1994. Height, competition and yield potential in winter wheat. Euphytica, 74: 9-17.
Direct Link - Tokatlidis, I.S., 2002. The effect of improved potential yield per plant on crop yield potential and optimum plant population in maize hybrids. J. Agric. Sci., 137: 299-305.
Direct Link