
Eltahir S. Ali
Department of Crop Science, Universiti Putra Malaysia, 43400- UPM, Serdang, Selangor, Malaysia
Ghizan B. Saleh
Department of Crop Science, Universiti Putra Malaysia, 43400- UPM, Serdang, Selangor, Malaysia
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 1 | Page No.: 65-70







ABSTRACT
Two cycles of phenotypic mass selection for ear length were conducted on two sweet corn (Zea mays L. saccharata) populations. In both populations, viz. the Manis Madu derived (M) and the population derived from its cross with Hybrid SSC 240 (MS), the realised cumulative response to selection was generally much lower than the predicted response. In the M populations, the predicted response from individual cycles was higher in the second cycle (M C2) (21.52%) than it was in the first (M C1) (19.31%), while the cumulative predicted response in M C2 was 45.94%. In the MS populations, a similar trend was seen, where the predicted response from individual cycles was 23.61% in MS C2 compared to 16.79% in MS C1, while the cumulative predicted response was 43.21% in MS C2. The two populations responded differently to the two cycles of selection, where, in the Manis Madu derived population, average realised response was attained only in the first cycle of selection (4.08%), while negative average response was observed in the second (–0.65%). The opposite was observed in the cross population, as average realised responses were –0.69 and 2.08% in MS C1 and MS C2, respectively. The average cumulative realized response was higher in M C2 (3.40%) than it was in MS C2 (1.38%). In the populations generated from Manis Madu, high broad-sense heritability estimates were revealed by plant height (89.1%), kernel total soluble solids (TSS) (86.2%), ear height (67.9%) and number of kernels/row (55.0%). Moderate estimates were shown by ear length (42.3%), fresh ear yield (36.8%) and ear weight (31.5%). In the cross populations, the highest broad-sense heritability estimate was revealed by number of ears/ha (99.8%), while low to moderate estimates were given by ear length (39.0%), days to maturity (27.9%) and days to tasseling (25.1%). In the improved populations, M C2 and MS C2, plant height revealed high positive correlations with most of the traits investigated. Positive correlations were also observed among ear traits. It was concluded that phenotypic mass selection was found efficient for improvement of the two sweet corn populations, but more cycles would be required for more pronounced results.
PDF Abstract XML References Citation
How to cite this article
Eltahir S. Ali and Ghizan B. Saleh, 2003. Response of Two Cycles of Phenotypic Mass Selection and Heritability on Two Tropical
Sweet Corn (Zea mays L. saccharata) Populations. Asian Journal of Plant Sciences, 2: 65-70.
DOI: 10.3923/ajps.2003.65.70
URL: https://scialert.net/abstract/?doi=ajps.2003.65.70
DOI: 10.3923/ajps.2003.65.70
URL: https://scialert.net/abstract/?doi=ajps.2003.65.70
INTRODUCTION
Mass selection was proven effective for the improvement of certain traits in many crop species, but consistent results were not usually realized until the method was improved and modified by Gardner (1961). The quite prominent modification suggested was the grading of the selection block by dividing it into smaller sub-units (grids) to minimize environmental effects. The improved technique and positive results reported renewed interest in the use of mass selection for the improvement of maize and other crop species. Lonnquist (1967) who applied five generations of mass selection for increased productivity in corn using correlated characters to prolificacy, found that the gain in yield per cycle of 6.25% approximated that made when direct selection for weight of grain per plant was used.
Subandi (1990) found that, after 10 cycles of mass selection for prolificacy in the BC10 maize composite, grain yield was increased by 2.6% per cycle. In a shruken-2 sweet corn population, mass selection was used to improve emergence in warm and cold germination. Cold germination increased from 47% in cycle 0 to 64% in cycle 10, whereas warm germination increased from 46% in cycle 0 to 77% in cycle 10 (Chang, 1990). Maita and Coors (1996) reported that, after twenty cycles of biparental mass selection for prolificacy in the open-pollinated maize population Golden Glow, ears per plant increased by 3.3% per cycle, from an average of 0.98 ears per plant for C0 to 1.63 ears per plant for C20.
Lopez-Reynoso and Hallauer (1998) reported that divergent mass selection was effective when selection for long ears was carried out, as measurement for the trait was improved by 1.4% per cycle. Biasutti et al. (2000), reported that grain yield increased significantly with the application of all selection criteria used, which include ear length, prolificacy, silking interval and harvest index. For ear length specifically, an increase of 2.8% was reported after four cycles of selection were performed.
Hansen et al. (1977), Becker (1984) and Fernandez and Miller (1985) reported that, heritability could be calculated from variance components of the ANOVA based on the method used to partition the genetic variance and its components in the calculations. Daniel and Batjay (1975), who estimated broad-sense heritability (h2B) from variance components in a diallel cross involving 11 sweet corn inbred lines, found high estimates for ear and plant height and number of kernel rows/ear.
The study was conducted to investigate the response of two cycles of phenotypic mass selection for ear length on two improved populations of sweet corn. In addition, broad-sense heritability for important traits and general phenotypic correlations among traits were also estimated.
MATERIALS AND METHODS
The research work was conducted at Field 2 and Share Farm, Universiti Putra Malaysia, Serdang, Selangor, Malaysia during May 1999 to June 2001. Two sweet corn base populations were used to initiate selection, viz. a Malaysian open-pollinated composite variety, Manis Madu (designated as M C0) and an intermated F2 segregating population derived from the cross between Manis Madu and the imported hybrid variety, SSC 240 (designated as MS C0). Populations M C1 and M C2 were derived from the first and second cycles, respectively, of phenotypic mass selection performed on M C0, while MS C1 and MS C2 were populations from the same cycles of selection, respectively, performed on MS C0. All populations carried the shrunken-2 gene conferring sweetness of the kernel in its recessive form (sh2).
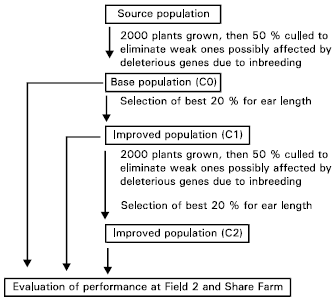 | |
| Fig. 1: | The phenotypic mass selection and evaluation procedures undertaken on the two sweet corn populations |
The phenotypic mass selection procedure employed, was a modification of that by Bletsos and Goulas (1999) and is illustrated in Fig. 1. The selection procedure was applied to both base populations for two successive cycles. Experimental design used was randomized complete block design, with three replications. Each plot measured 5 x 3.75 m2 and the planting density used in the plots was 0.75 x 0.25 m2. Two to three seeds were sown per point manually, but later thinned to a single plant per point two weeks after planting. A plot comprised of five rows, each measuring 5 m long, planted with twenty plants. Normal agronomic practices were employed in all experiments. Data analysis was first carried out separately for the two locations, followed by combined analysis, after tests of homogeneity of error variances were performed on the data from the two locations.
The predicted and realized responses to selection were estimated and compared. The predicted response to selection was estimated using the equation used by Simmonds (1979) and Knight (1979), as follows:
R = I h2 σ |
where:
| R | = | Predicted response to selection |
| I | = | Intensity of selection |
| h2 | = | Heritability of trait |
| σ | = | Phenotypic standard deviation of the parental population |
The intensity of selection (I) was estimated by the following equation, as described by Becker (1975):
I = 1.13 + 0.73 log 1/k
where: k = the selected fraction of the population
Means were presented with their standard errors, for the purpose of making comparison. Heritability estimates and their standard deviations were also shown, as they constitute the basic components of the predicted gain equation. The cumulative response was estimated as a percentage of the original population (C0), for both cycles. The response to selection from one cycle to another was based on the performance of the respective parental population.
Broad-sense heritability was estimated using the formula described by Singh and Chaudhary (1977) and Becker (1984). While simple phenotypic correlations among traits investigated were determined according to the formula described by Gomez and Gomez (1984).
Results
Response to selection: In M C2 population, the predicted cumulative response was 45.94% (Table 1). The predicted response from individual cycles was higher in M C2 (21.52%) than that in M C1 (19.31%). In MS C2 population, the same trend was revealed, as the predicted cumulative response was 43.21%, with the predicted response from individual cycles was higher in MS C2 (23.61%) compared to that in MS C1 (16.79%). The standard deviation value for ear length measurements in the improved population after the two cycles of selection was higher in the cross population (2.40) compared to that in Manis Madu derived population (2.05) (Table 1).
Results on the realized response to selection for ear length estimated on the improved sweet corn populations of Manis Madu (M) and its cross with SSC 240 (MS), evaluated at the two locations (Table 2). For the populations derived from Manis Madu, at Field 2, the realised response to selection from individual cycles was 5.80% in M C1, but was negative in M C2 (-1.37%). The realised cumulative response to selection in M C2 over M C0 was therefore, 4.35%. At Share Farm, the realised response to selection from individual cycles was 1.92 and 1.26% respectively, for M C1 and M C2 and cumulative realised response was 3.21% in M C2 over M C0. The average realised response to selection from individual cycles were 4.08 and –0.65%, respectively for M C1 and M C2 and the average realised cumulative response in M C2 was 3.40%.
For the crossed populations, from evaluation at Field 2, the realised response from individual cycles was 0.71 and 0.70 respectively, for MS C1 and MS C2, while the cumulative realised response in MS C2 was 1.42%. At Share Farm, MS C1 showed a negative a realised response to selection of –1.34%, while MS C2 showed a realised response of 2.72%. The cumulative realised response in MS C2 was 1.34%. Average realised response to selection was –0.69 and 2.08% respectively, for MS C1 and MS C2, while the average cumulative response in MS C2 was 1.38%. It was generally observed that in both populations, the realised responses to selection were much lower than the predicted responses.
Broad-sense heritability: In the Manis Madu derived populations, high broad-sense heritability estimates were revealed by plant height (89.1%), total soluble solids (86.2%), ear height (67.9%) and number of kernels/row (55.0%) (Table 3). All other plant traits showed low broad-sense heritability estimates, except for ear diameter, number of ears/ha and number of kernel rows/ear, which showed either zero or negative values for the estimates. For the Manis Madu x SSC 240 derived populations, highest broad-sense heritability was revealed by number of ears/ha (99.8%), while moderate estimates were given by ear length (39.0%), days to maturity (27.9%) and days to tasseling (25.1%). Low heritability was shown by number of kernels/row (4.1%). Other traits gave zero or negative estimates of broad-sense heritability (Table 3).
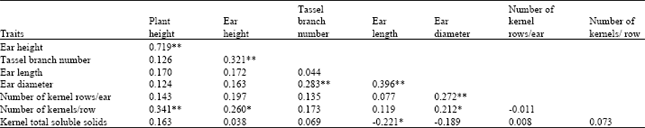
Simple phenotypic correlations within improved populations: Simple phenotypic correlations among important plant traits measured on the two improved populations, M C2 and MS C2 were computed for data from the two locations combined (Table 4). Ear length was positively correlated (P ≤ 0.01) with ear diameter (r =0.396), but negatively correlated (P ≤ 0.05) with kernel TSS (r = –0.221). Plant height was positively correlated with ear height (r = 0.719) and number of kernels/row (r = 0.341) (P ≤ 0.01). Ear height also showed positive correlation with tassel branch number (r = 0.321) (P ≤ 0.01) and number of kernels/row (r = 0.260) (P ≤ 0.05). Tassel branch number was positively correlated (at p ≤ 0.01) with ear diameter (r = 0.283), while ear diameter was positively correlated with number of kernel rows/ear (r = 0.272) (P ≤ 0.01) and number of kernels/row (r = 0.212) (P ≤ 0.05).
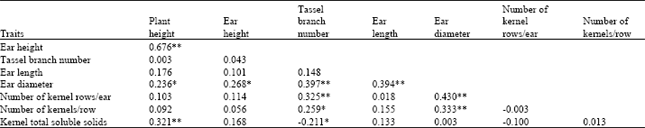
Ear length was positively correlated (P ≤ 0.01) with ear diameter (r = 0.394), while plant height was positively correlated with ear height (r = 0.676), kernel TSS (r = 0.321) (P ≤ 0.01) and ear diameter (r = 0.236) (P ≤ 0.05) (Table 5). Ear height showed positive correlations with ear diameter (r = 0.268) (P ≤ 0.05). Tassel branch number showed positive correlations with ear diameter (r = 0.397), number of kernel rows/ear (r = 0.325) (P ≤ 0.01) and number of kernels/row (r = 0.259) (P ≤ 0.05). Tassel branch number was however, negatively correlated (P ≤ 0.05) with kernel TSS content (r = –0.211). Ear diameter revealed positive correlation with number of kernel rows/ear (r = 0.420) and number of kernels/row (r = 0.333) (P ≤ 0.01).
Discussion
Response to selection: As expected, in both populations, the cumulative predicted responses to selection for ear length were comparatively higher than those from the individual cycles. This implies that selection for ear length had contributed to some improvement in its magnitude in the improved populations. The predicted increase was very gradual, from the first cycle and eventually the second cycle. Comparing the standard deviation values between the improved populations, the improved population, MS C1 was more variable than the improved population, M C1 (Table 1). This was expected because combination of the two parental populations in the cross had introduced new introgression of dissimilar genes in the new population.
| Table 1: | Estimates of predicted response to selection for ear length measured on improved sweet corn populations derived from Manis Madu (M) and the cross of Manis Madu x SSC 240 (MS), after two cycles of phenotypic mass selection |
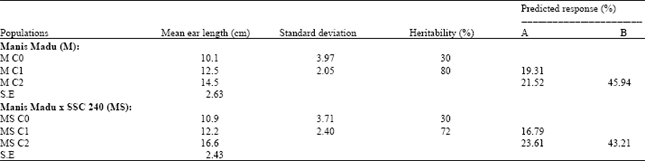 | |
| Table 2: | Mean values and realized response to selection for ear length measured on improved sweet corn populations derived from Manis Madu (M) and the cross of Manis Madu x SSC 240 (MS), after two cycles of phenotypic mass selection, evaluated at Field 2 and Share Farm |
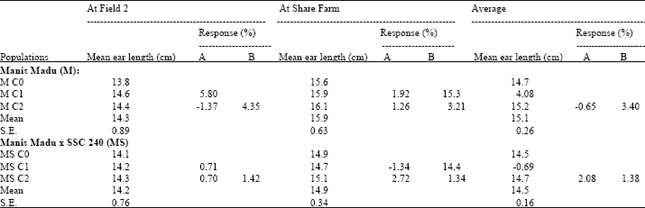 | |
| Table 3: | Genetic variances (σ2G), phenotypic variances (σ2P) and broad-sense heritability estimates (h2B) for traits measured on Manis Madu and the cross Manis Madu x SSC 240 (MS C2) populations |
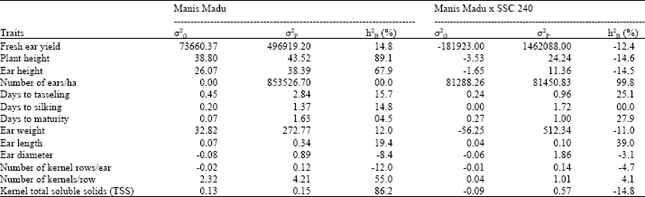 | |
| Table 4: | Simple phenotypic correlations among traits measured on improved sweet corn population M C2, at the two locations combined |
 | |
| Table 5: | Simple phenotypic correlations among traits measured on improved sweet corn population MS C2, evaluated at the two locations combined |
 | |
On realised response, the improvements after the two cycles of selection were also gradual in both populations involved, although were much lower than the predicted responses. The two populations responded differently to the two cycles of selection.
For the Manis Madu populations, realised response from the second cycle of selection was negative at Field 2, because M C1 revealed higher ear length than M C2, despite the fact that ear length values for both improved populations were higher than that of the original population, M C0 (Table 2). These contributed to positive overall average response to the first cycle, but negative response to the second. In the crossed population, ear length of MS C1 at Share Farm was lower than that of MS C0 (but the difference was not significant), resulting in a negative response in MS C1. As a result therefore, the average response to selection over the two locations was negative in the first cycle for the cross population (MS C1). The MS C2 population revealed reasonable improvements at both locations.
Broad-sense heritability: Broad-sense heritability estimated for plant traits measured on the populations of Manis Madu (M) showed that plant height, kernel TTS, ear height and number of kernels/row were highly heritable (Table 3). Ear length, which was used as the criterion for selection, had shown moderate broad-sense heritability estimate, indicating that the trait is an appropriate choice as criterion for selection. Ear diameter, number of kernel rows/ear and number of ears produced per hectare, which showed negative or zero heritability estimates were highly influenced by the environmental factors and therefore, not heritable.
Results on broad-sense heritability for plant traits measured on the crossed populations, Manis Madu x SSC 240 (MS), showed that the number of ears produced per hectare was most heritable. As in Manis Madu populations, ear length was also moderately heritable in the crossed populations. The flowering traits were also moderately heritable. The other plant traits were also observed as non-heritable, due to the high influence of the environmental factors. The influence was more highly manifested in the crossed populations than it was in Manis Madu derived populations, indicating that the cross was more sensitive to environmental changes. This might be attributed to the low adaptability of the hybrid parent involved in the cross, SSC 240 to the local environment. These results are in agreement with those obtained by Hansen et al. (1977), Rafii et al. (1994) and Fountain and Hallauer (1996), where yield and its components showed lower estimates of h2B than did other growth characters like plant height, ear height and flowering traits.
Simple phenotypic correlations within improved populations: In both populations, very high positive correlations were revealed among most ear traits, especially ear length, ear diameter and number of kernels/row. In M C2, significant positive correlations were found between plant height and ear height and to some extent between these two traits and the ear traits, including ear length, ear diameter, number of kernel rows/ear and number of kernels/row. In MS C2, the correlations between plant height, ear height and ear traits were less manifested.
Both ear length and ear diameter are important quality traits for marketability of sweet corn, as big and long ears are preferred. These traits should therefore, be given proper attention in selection programmes, which would be helpful to breeders in utilizing these improved populations. Ear length, which was taken as the criterion for selection in this study, had shown high correlation with ear diameter in both populations. This trait should therefore, be continuously used as the criterion in further selections involving these selected populations. This would enable breeders to obtain populations with the best ear diameter, while selecting for longer ears, thus increasing yield.
In conclusion, phenotypic mass selection for ear length was reasonably effective for improvement of the sweet corn populations, although positive average realised response was achieved only in the first cycle of selection for Manis Madu population and in the second cycle for the cross population. More pronounced improvement could be made if selection on the improved populations is to be continued beyond the two cycles. Other plant traits such as ear diameter and number of kernel rows/ear could also be considered as criteria in further selection programmes on these populations.
REFERENCES
- Fernandez, G.C.J. and J.C. Miller Jr., 1985. Estimation of heritability by parent-offspring regression. Theor. Appl. Genet., 70: 650-654.
CrossRefDirect Link - Gardner, C.O., 1961. An evaluation of effects of mass selection and seed irradiation with thermal neutrons on yield of corn. Crop Sci., 1: 241-245.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., New York, ISBN: 9780471870920, Pages: 704.
Direct Link - Lopez-Reynoso, J.J. and A.R. Hallauer, 1998. Twenty-seven cycles of divergent mass selection for ear length in maize. Crop Sci., 38: 1099-1107.
Direct Link - Maita, R. and J.G. Coors, 1996. Twenty cycles of biparental mass selection for prolificacy in the open-pollinated maize population Golden Glow. Crop Sci., 36: 1527-1532.
Direct Link - Rafii, M.Y., G. Saleh and T.C. Yap, 1994. Response to simple and full-sib reciprocal recurrent selection in sweet corn varieties Bakti-1 and Manis Madu. J. Malaysian Applied Biol., 22: 173-180.
Direct Link - Singh, R.K. and B.D. Chaudhary, 1977. Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publisher, New Delhi, India, ISBN: 8176633070, Pages: 318.
Direct Link