
M.N. Sundararaman
Department of Animal Genetics and Breeding, Madras Veterinary College, Tamilnadu Veterinary and Animal Sciences University, Chennai-600 007, India
M.J. Edwin
Department of Animal Genetics and Breeding, Madras Veterinary College, Tamilnadu Veterinary and Animal Sciences University, Chennai-600 007, India
Asian Journal of Cell Biology
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 22-33







ABSTRACT
Artificial Insemination (AI) with frozen Boer goat semen could be a viable option for genetic upgradation of non-descript local goats. To maximize the utilization of Boer germplasm, minimizing the losses due to poor freezability and improving the quality of frozen semen are the areas to be addressed. A study was conducted to assess the deterioration in the quality of semen in terms of motility characteristics of spermatozoa during the processing of semen for cryopreservation using Computer Assisted Semen Analysis (CASA) technique. A highly significant decline in sperm motility, progressive motility, path velocity and progressive velocity was noticed during equilibration. Semen samples with acceptable freezability differed from samples not freezing well by not showing any significant decrease in velocity characteristics of the spermatozoa during glycerol-equilibration. The decline was significant in semen samples that froze poorly. Comparing the motility characteristics of spermatozoa during glycerol-equilibration in the good and poor samples, the progressive velocity in poor samples was significantly lower than that of good samples at 3 and 4 h of equilibration. The curvilinear velocity of spermatozoa was also significantly less in poor samples than good samples after 4 h. These results indicate that velocity of sperm motion plays a vital role in determining the post-thaw quality of frozen semen. Cryopreservation significantly affected the quality of frozen semen by lowering all the motion characteristics post-thawing. This study also revealed that the duration of exposure of spermatozoa to the cryoprotectant during equilibration is critical in cryopreservation of Boer cross goat semen. A shorter duration of 2 h of equilibration with glycerol at 5°C may be beneficial for Boer goat semen cryopreservation.
PDF Abstract XML References Citation
How to cite this article
M.N. Sundararaman and M.J. Edwin, 2008. Changes in Motility Characteristics of Goat Spermatozoa During Glycerol-Equilibration and the Relevance to Cryopreservation
. Asian Journal of Cell Biology, 3: 22-33.
DOI: 10.3923/ajcb.2008.22.33
URL: https://scialert.net/abstract/?doi=ajcb.2008.22.33
DOI: 10.3923/ajcb.2008.22.33
URL: https://scialert.net/abstract/?doi=ajcb.2008.22.33
INTRODUCTION
Artificial Insemination (AI) with frozen semen has been proved as the most potent method for rapid genetic improvement in domestic animals. Although the basic principle of cryopreservation is similar for spermatozoa of most mammalian species, the sperm from different species may react differently to freezing due to their difference in morphology and certain biochemical constituents. Therefore, a cryopreservation protocol developed for one species may not be ideal for sperm of other species.
The goat plays a vital role in the economy of the poor and marginal farmers of rural India. Not withstanding the fact that the goat population has shown a steady growth over the years very little structured effort has been taken for genetic improvement at farm level. In recent years, use of the South African Boer goat has been contemplated for the upgrading of the local non-descript goats in India especially on an experimental basis. Considering the high cost of importation of purebred Boer goats, AI using frozen semen is practiced to produce superior progeny and accelerate the upgrading of stock.
Studies on semen production potential and the freezability of Boer semen have indicated Boer spermatozoa to be successfully cryopreserved for use in A.I. (Tuli and Holtz, 1995; Sundararaman and Edwin, 2003). Thus to maximize the potential of frozen goat semen production of superior germplasm for wider application, it is essential, not only to minimize the losses due to poor freezability of samples but also to improve the quality of semen following freezing as well, as an effort of achieving better fertility results following AI.
At each stage of the cryopreservation cycle, which includes the entire process of semen collection, dilution, equilibration and freezing, the spermatozoa may lose the ability to fertilize normally (Watson, 1995).
It is therefore mandatory to optimize a cryopreservation cycle, which would cryopreserve the largest number of structurally and functionally normal spermatozoa. The equilibration of extended semen at 5°C is an essential step in many protocols for the cryopreservation of mammalian spermatozoa. Equilibration facilitates the cryoprotectant, glycerol present in the semen diluent to exert a beneficial effect in terms of minimizing the degree of structural damage to spermatozoa during deep freezing.
Glycerol, as a cryoprotectant in diluents has yielded successful results in the cryopreservation of ram (Salamon and Maxwell, 2000; Gil et al., 2003) and goat (Leboeuf et al., 2000; Sundararaman and Edwin, 2003; Peterson et al., 2006; Purdy, 2006) semen. However, glycerol is somewhat toxic to spermatozoa (Holt, 2000) and may induce osmotic damage (Purdy, 2006). The addition of glycerol by itself may cause certain structural damage and lowered motility of spermatozoa (Almquist and Wiggen, 1973; Colas, 1975; Watson, 1981; Hammerstedt et al., 1990; Bhosrekar et al., 1994; Das and Rajkonwar, 1994, 1996; Maxwell and Watson, 1996; Salamon and Maxwell, 2000; Sonmez and Dimirci, 2004).
The semen equilibration time at 5°C generally varies for cryopreservation of spermatozoa in different species. The period for each species has been standardized based on post-thaw motility as microscopically assessed under phase contrast (Edwin and Ulaganathan, 1988). Irrespective of the freezing protocol employed, variations have been observed between males regarding freezability and fertility of semen, so they could be classified as good freezers or bad freezers (Leboeuf et al., 2000). This variability is relatively independent of prior semen quality and the semen of certain individuals consistently freezes with less cryoinjury than that of others (Corteel et al., 1987). The fact that males can often be classified as good freezers or bad freezers implies that certain characteristics of membrane structure, which may be genetically determined, predispose towards survival under cryopreservation stress (Watson, 2000). There are instances in which certain semen samples within a species having good pre-freeze motility, still results in poor freezability. Differences in either ejaculation frequency, or in epididymal transit times and sperm mixing in the epididymis, provide a potential mechanism for variability in responses to subsequent temperature explaining why ejaculates within individuals can vary in their responses to cryopreservation (Watson, 1995). Further, it is of interest to explore whether any adverse changes that take place during exposure to the cryoprotectant during equilibration influence the freezability of spermatozoa.
Sperm motility, in general and characteristics of sperm motion in particular could be some of the indicators of the quality of spermatozoa. Commonly, by evaluating the proportion of progressive motile percent at different stages, the quality of semen is monitored. However, evaluation of characteristics of sperm motion may provide valuable information on why certain samples despite containing good proportion of progressive motile spermatozoa pre freezing poorly freezable.
Computer assisted semen analysis technique has been used to provide precise and accurate information on sperm motion characteristics. Therefore, a study was undertaken to analyze the changes in motility characteristics of Boer cross spermatozoa during equilibration at 5°C by Computer Assisted Semen Analysis (CASA) technique and its relevance in post-thaw survival of spermatozoa.
MATERIALS AND METHODS
Experimental Animals
Three Boer half-bred young males maintained under stall-fed conditions at Semen Bank, Madras Veterinary College, Chennai, India were used for the study. The growing males were provided with water and green fodder at-libitum, which included a kg of green lucerne/animal/day. In addition, 300 to 350 g of concentrate/animal/day was also fed. The research was undertaken during May and June 2004.
Semen Collection, Evaluation and Processing
The bucks (3-5 years of age) were trained to mount a teaser. Invariably one of the bucks was alternately used as a teaser. Semen was collected by means of the artificial vagina. Prior to semen collection, the goats were sexually aroused, by utilizing visual, olfactory and auditory stimuli. After arousal, 2 or 3 false mounts were allowed to ensure a good quality ejaculate. Semen collections were done twice a week. A total of 27 samples were collected. The samples were evaluated for ejaculate volume, sample colour, consistency, mass activity, sperm concentration and progressive motility by routine methods at room temperature (25-27°C). Nevertheless, the motility evaluation was done at 37°C. Semen samples having good quality with at least 70% of progressive motile spermatozoa were selected for freezing. Samples were extended in a Tris-egg yolk based diluent containing 20% egg yolk and 7% glycerol (Sivaselvam et al., 2000) in such a way that each insemination dose contained 1x106 motile spermatozoa.
Equilibration of the extended semen was carried out for 4 h in a refrigerated cabinet (5°C). A small portion of the extended semen sample was further prepared for the Computer Assisted Semen Analyses (CASA), by adjusting the sperm concentration to 20x106 mL-1 using Tris buffer. The samples under glycerol-equilibration were evaluated for motility using CASA technique at 0, 1, 2, 3 and 4 h of incubation at 5°C.
| Table 1: | Analysis set-up for HT-IVOS Version 10.9 used to evaluate goat spermatozoa |
 | |
CASA Analysis
CASA analysis was done using Hamilton-Thorne integrated visual optical system (HT-IVOS) (Version 10.9). The analysis set-up used to evaluate the goat semen is set out in Table 1 with the chamber temperature being set to 37°C. Using a micro-pipette, 4 μL of the prepared semen sample was loaded on the Makler counting chamber (Self-medical Insts Ltd.) and a cover glass was placed on the droplet. Six microscopic fields were analyzed for each semen sample.
Equilibrated semen samples were packaged into French straws (0.25 mL) for cryopreservation. After 24 h of storage at -196°C, the cryopreserved semen samples were thawed in a water bath at 37°C for 30 sec. The CASA analyses of the post-thaw samples were performed by repeating the procedure adopted for the samples pre-freezing.
Sperm Motility (SM): spermatozoa that are moving at or above minimum speed as determined by values defined under set-up (%); Progressive Motility (PSM): spermatozoa moving with path velocity at or above 25 μm sec-1 and straightness at or above 70%; path velocity (VAP): the average velocity of the smoothened cell path (μm sec-1); progressive velocity (VSL): The average velocity measured in a straight line from the beginning to the end of track (μm sec-1); Curvilinear velocity (VCL): The average velocity measured over the actual point-to-point track followed by the cell (μm sec-1); lateral amplitude of head (ALH): corresponds to the mean width of the head oscillation as the sperm swim (μm); Beat Cross Frequency (BCF): frequency with which the sperm track crosses the sperm path in either direction (Hz); straightness (STR): measures the departure of the cell path from a straight line. It is the average value of ratio of VSL/VAP (%); linearity (LIN): measures the departure of the cell track from a straight line. It is the average value of ratio of VSL/VCL (%), were the sperm motility characteristics analyzed by CASA technique.
Statistical Analyses
Based on post-thaw progressive motility, samples recording 40% or more progressive motility were classified as Good and those with less motility as Poor freezability samples. In general, the post thaw progressive motility of goat semen varies from 36.1% (Deka and Rao, 1987) to 46% (Aboagla and Terada, 2004). Tuli and Holtz (1995) reported the motility of Boer goat semen post thaw as 29, 35, 39 and 45% at different seasons. Furthermore, in a pilot study on fertility in goats, conducted by this department using cryopreserved semen with 40% and above motility post thaw, an acceptable kidding rate was achieved on cervical insemination. Therefore, semen samples with 40% and above progressive motility were classified as Good samples in this study. The results presented as mean ± SE, were analyzed by a one-way analysis of variance using microstat software (Ecosof Inc., 1984, Balitimore, USA). Statistical significance was set at p≤0.05 and p≤0.01. The means were tested as per Duncan (1995) for significance.
RESULTS AND DISCUSSION
Glycerol-Equilibration on Sperm Motility Characteristics
A gradual and significant decline in sperm motility and progressive motility of spermatozoa were recorded during the glycerol-equilibration period of from 0 to 4 h (Fig. 1). Regarding the velocity parameters, glycerol-equilibration resulted in a highly significant reduction of path velocity and progressive velocity (Fig. 2). However, the consistent decrease in curvilinear velocity was not statistically significant. The inconsistent variation in mean values for lateral amplitude of head beat cross frequency, straightness and linearity did not show any significance.
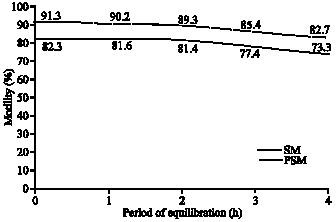 | |
| Fig. 1: | Sperm motility following glycerol-equilibration in Boer cross goat semen |
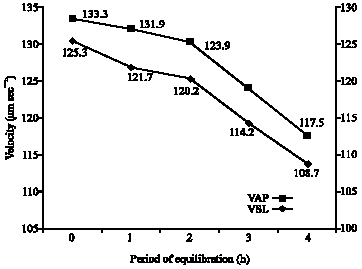 | |
| Fig. 2: | Sperm velocity following glycerol-equilibration in Boer cross goats |
Glycerol-Equilibration on Sperm Motility Characteristics-Good and Poor Freezable Samples (Pre-Freeze Changes)
Both in good and poor freezability samples, the lowering of proportion of total sperm motility and progressive sperm motility during equilibration were highly significant (Table 2). However, in both classes of semen samples, the decrease in SM and PSM was not significant up to 2 h of glycerol-equilibration but highly significant thereafter. A gradual non-significant decline in all the velocity characteristics, namely path velocity, progressive velocity and curvilinear velocity in good freezability samples were observed for the 4 periods of equilibration, respectively. Whereas, in poor freezability semen samples, the reduction in the velocity parameters was not significant only up to 2 h of equilibration, but the decrease was significant thereafter for path velocity, progressive velocity and curvilinear velocity, respectively. For other motility characteristics the variation exhibited was inconsistent and not significant.
| Table 2: | Means (±SE) for motility characteristics of Boer cross goat spermatozoa during glycerol-equilibration |
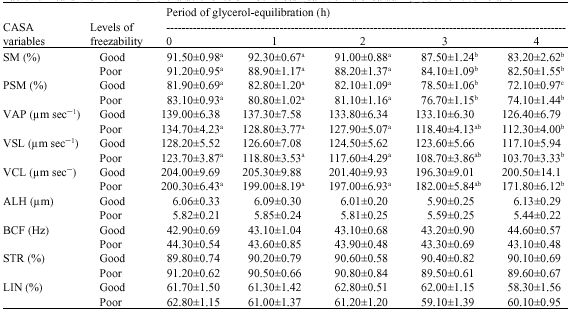 | |
| Means bearing different superscripts in a row differ significantly for Good (P≤0.01) and Poor (P≤0.05) freezability samples, No. of good samples = 10, No. of poor samples = 17, SE: Standard Error | |
 | |
| Fig. 3: | Progressive velocity of Boer cross goat sperm following glycerol-equilibration |
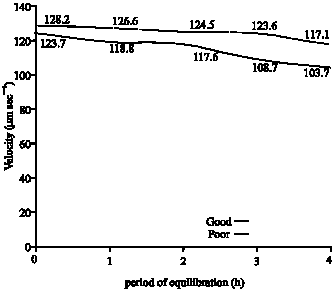
The mean values recorded for progressive velocity of spermatozoa during equilibration for good freezability samples were significantly higher at 3 and 4 h than those for poor freezability semen samples (Fig. 3). The mean values obtained for curvilinear velocity after 4 h of equilibration for good samples were also significantly higher than those recorded for poor freezability samples (Fig. 4).
Effect of Freezing on Sperm Motility Characteristics (Post-Thaw Changes)
Freezing had a significant effect on total motility, progressive motility, path velocity, progressive velocity, curvilinear velocity, beat cross frequency, straightness, linearity and lateral amplitude of head of goat spermatozoa (Table 3). The good freezability samples recorded a highly significant decrease in total motility and progressive motility and also a significant reduction in all the velocity parameters and beat cross frequency. In contrast, semen with poor freezability showed a highly significant decline, not only in total motility and progressive motility, but also other parameters like the three velocity parameters and linearity besides a significant reduction in beat cross frequency and straightness.
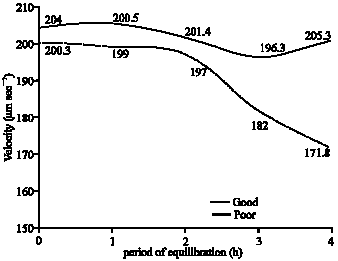 | |
| Fig. 4: | Curvilinear velocity of Boer cross goat sperm following glycerol-equilibration |
| Table 3: | Means (± SE) for sperm movement characteristics in Boer cross goats at the pre-freeze and post-thaw stages of cryopreservation cycle (n = 27) |
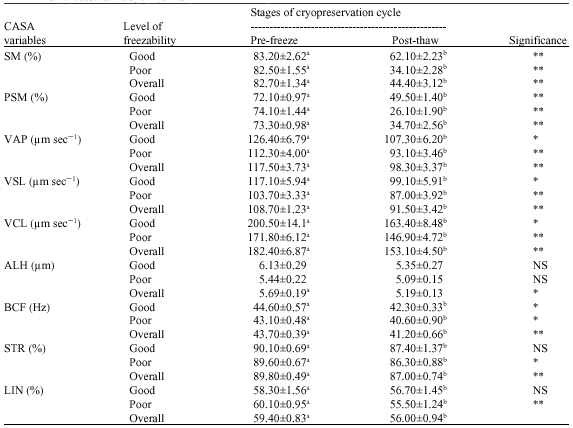 | |
| **Means bearing different superscripts in a row differ significantly, ** = Highly significant (p≤0.01), * = Significant (p≤0.05), NS =Not significant, SE: Standard Error, N: Number of samples | |
In this trial, during equilibration, not only did the motility, but also the velocity characteristics of the spermatozoa, viz., path velocity and progressive velocity show a decrease. The decline in sperm motility during equilibration may be due to formation of peroxides from free radicals. This oxidative stress causes damage to biomolecules and cellular components (Halliwell, 1991). The sperm membrane contains a high number of unsaturated fatty acids, which are predisposed to damage due to peroxidation, which destroys the structural integrity of plasma membrane leading to loss in motility (Aitken et al., 1989; Jones and Mann, 1977b; Salamon and Maxwell, 2000). Further, the endogenous phospholipids of damaged spermatozoa in ejaculates undergo peroxidation and the Reactive Oxygen Species (ROS) produced may be toxic to the normal spermatozoa (Jones and Mann, 1977a; Nichi et al., 2006), which, perhaps also contributes to altering the speed of sperm movement. Although semen contains substantial quantities of antioxidants to neutralize the peroxide formation (Lewis et al., 1997), the endogenous antioxidative property of semen may be inadequate during dilution and storage Maxwell and Salamon (1993), Colas (1975), Watson (1981) and Maxwell and Watson (1996) also reported the toxicity of cryoprotective agent glycerol in semen diluents, to bring about a reduction in sperm motility and to alter the acrosome integrity by interfering with the permeability of the sperm membrane.
In this experiment, all 27 samples that were subjected to cryopreservation showed 70% or more of progressive sperm motility after glycerol-equilibration for 4 h before freezing. Of these, however, only 10 samples (classified as Good freezability samples) exhibited satisfactory results after freezing for A.I. The evaluation on glycerol-equilibration in good and poor freezable semen samples has revealed that both classes of samples showed similar trends for changes in total motility and progressive motility besides changes in certain other motility characteristics namely, lateral amplitude of head, beat cross frequency, straightness and linearity. However the decrease in speed of spermatozoa in terms of path velocity, progressive velocity and curvilinear velocity was significant only in samples that froze poorly. Furthermore, on comparing the movement characteristics of spermatozoa between good and poor freezable samples during glycerolation, it was found that the progressive speed of spermatozoa in the inferior samples was significantly lower than that of the samples that froze satisfactorily following 3 and 4 h of glycerol-equilibration (Fig. 3). In addition, the curvilinear velocity of spermatozoa was also significantly less in the weaker samples at 4 h of glycerolation (Fig. 4). These findings were suggestive that it could possibly be the speed of sperm motility at pre-freeze stage (after 4 h of glycerol-equilibration), which could have predominantly influenced the quality of frozen spermatozoa. Van Duijn Jr. (1962) has reported that more than the initial velocity of spermatozoa, the rates of velocity-decrease and the number of spermatozoa moving normally are more important with respect to motility characteristics. Therefore, the rate of velocity retardation of spermatozoa could be considered as an indicator for predicting livability and quality of sperm (Chakrabarti, 1993).
On evaluation of effect of freezing, it was observed that, all the motility characteristics of spermatozoa were significantly influenced by freezing and thawing, which reiterates earlier findings in Boer cross and Barbari goats (Sundararaman and Edwin, 2005). Even among good and poor freezable samples there was not much of difference in sperm behavior due to freezing and thawing. All the sperm motility and velocity parameters were significantly reduced in the post-thawing semen. Changes in the osmotic pressure during semen processing for cryopreservation critically affect the spermatozoa. This may be the most important deterrent to sperm survival during cryopreservation (Watson, 1995). Furthermore, membrane destabilization can occur when the sperm plasma membrane undergoes a phase transition from the liquid crystalline phase to the gel phase due to a decrease in temperature (Barrea-Compean et al., 2005). The irreversible changes in the sperm membrane induced by lipid phase transitions during cooling warming may possibly affect the movement characteristics of spermatozoa during semen processing for cryopreservation (Holt and North, 1984; Deleeuw et al., 1990). In addition, frozen-thawed sperm are more vulnerable to oxidative stress due to peroxidation than sperm in freshly diluted semen (Neild et al., 2005). As semen is diluted many fold in the extender it reduces the total antioxidant concentration in the medium and cells (Kumar and Das, 2005). Many sperm are killed during cryopreservation. Thus, it is likely that cryopreserved sperm cells are posed to more ROS concentration and therefore many of the surviving cells post-thaw exhibit as if they are capacitated or acrosome reacted (Bailey et al., 2000). The overall effects of these events may adversely affect quality of post thawing semen.
The present study indicated glycerol-equilibration to significantly affect sperm motility and velocity or speed characteristics in Boer goat semen. In the freezing of goat semen, the quality of sperm motility in terms of speed is important in determining the post-thawing survivability of the spermatozoa in terms of progressive motility rather than the proportion of motile spermatozoa alone. Further, the velocity measurements of sperm motility are useful in predicting the fertility of semen (Holt et al., 1985; Budsworth et al., 1988; Aitken, 1990; Sarma et al., 1996; Tardiff et al., 1997; Kirk et al., 2005). Therefore more emphasis has to be given to sperm velocity in semen processing and the quality assessment of post-thaw spermatozoa. Rapidly moving post-thaw sperm seem to have a high probability of crossing the cervical barrier during AI by cervical deposition. And this approach may be more appropriate for small ruminants (Eppleston and Maxwell, 1993). The glycerol-equilibration period in the processing of semen may be optimized based not only on the evaluation of sperm motility but also on the assessment of the velocity measurements using the CASA technique. This enables rapid, precise and objective evaluation of sperm movement (Budsworth et al., 1988; Amann, 1989; Suttiyotin and Thwaites, 1992; Yeung et al., 1992). Thus the loss of precious germplasm due to poor quality of post-thaw semen can be minimized and the quality of cryopreserved spermatozoa in terms of fertility can be enhanced.
CONCLUSION
The current understanding with regard to differences in cryosurvival between good and poor semen freezers is based solely on the ability of spermatozoa to withstand the drastic changes that occur during the actual freezing process and exposure to liquid nitrogen vapour. However, the present study has shown that the challenges that the spermatozoa face starts much earlier in the freezing protocol, viz., glycerol-equilibration. Glycerol-equilibration adversely influences the movement of goat sperm and the duration of exposure to the cryoprotectant becomes critical. A clear-cut difference, which starts at equilibration can ultimately determine the degree of post-thaw survival of spermatozoa of good and poor freezability semen samples. This study also shows the need to revisit cryopreservation protocols existing with emphasis on sperm movement for the different species, as the sensitivity of sperm to toxic effect of cryoprotectant vary. A short duration equilibration of 2 h with glycerol may be beneficial for goat semen. The application of the CASA technique in evaluation of the movement characteristics of spermatozoa, especially the speed and adoption of flexibility in devising cryopreservation protocols accordingly, would offer scope for potential improvements in quantity and quality of semen frozen in goats.
REFERENCES
- Aboagla, E.M.E. and T. Terada, 2004. Effects of egg yolk during the freezing step of cryopreservation on the viability of goat spermatozoa. Theriogenology, 62: 1160-1172.
CrossRefPubMedDirect Link - Aitken, R.J., J.S. Clarkson and S. Fishel, 1989. Generation of reactive oxygen species, lipid peroxidation and human sperm function. Biol. Reprod., 41: 183-187.
Direct Link - Bailey, J.L., J.F. Bilaodean and N. Cormier, 2000. Semen cryopreservation in domestic animals: A damaging capacitating phenomenon. J. Androl., 20: 1-7.
PubMed - De leeuw, F.E., H.C. Chen, B. Colenbrander and A.J. Verkleij, 1990. Cold-induced ultrastructural changes in bull and boar sperm plasma membranes. Cryobiology, 27: 171-183.
PubMed - Gil, J., N. Lundeheim, L. Soderquist and H. Rodriguez-Martinez, 2003. Influence of extender, temperature and addition of glycerol on post-thaw sperm parameters in ram semen. Theriogenology, 59: 1241-1255.
CrossRefPubMedDirect Link - Halliwell, B., 1991. Reactive oxygen species in living systems: Source, biochemistry and role in human disease. Am. J. Med., 91: S14-S22.
CrossRefPubMedDirect Link - Hammerstedt, R.H., J.K. Graham and J.P. Nolan, 1990. Cryopreservation of mammalian sperm: What we ask them to survive. J. Androl., 11: 73-88.
PubMed - Holt, W.V. and R.D. North, 1984. Partially irreversible cold-induced lipid phase transitions in mammalian sperm plasma membrane domains: Freeze-fracture study. J. Exp. Zoo., 230: 473-483.
PubMed - Holt, W.V., H.D.M. Moore and S.G. Hiller, 1985. Computer-assisted measurement of sperm swimming speed in human semen: Correlation of results with in vitro fertilization assays. Fert. Steril., 44: 112-119.
PubMed - Holt, W.V., 2000. Basic aspects of frozen storage of semen. Anim. Reprod. Sci., 62: 3-22.
CrossRefPubMedDirect Link - Jones, R. and T. Mann, 1977. Toxicity of exogenous fatty acid peroxides towards spermatozoa. J. Reprod. Fert., 50: 255-260.
PubMed - Kirk, E.S., E.L. Squires and J.K. Graham, 2005. Comparison of in vitro laboratory analyses with the fertility of cryopreserved stallion spermatozoa. Theriogenology, 64: 1422-1439.
CrossRefPubMedDirect Link - Leboeuf, B., B. Restall and S. Salamon, 2000. Production and storage of goat semen for artificial insemination. Anim. Reprod. Sci., 62: 113-141.
Direct Link - Lewis, S.E.M., E.S.L. Sterling, L.S. Young and W. Thompson, 1997. Comparison of individual antioxidants of sperm and seminal plasma in fertile and infertile men. Fert. Steril., 67: 142-147.
CrossRefPubMedDirect Link - Maxwell, W.M. and S. Salamon, 1993. Liquid storage of ram semen: A review. Reprod. Fert. Dev., 5: 613-638.
PubMedDirect Link - Maxwell, W.M.C. and P.F. Watson, 1996. Recent progress in the preservation of ram semen. Anim. Reprod. Sci., 42: 55-65.
CrossRef - Neild, D.M., J.P. Brouwers, B. Colenbrander, A. Aguero and B.M. Gadella, 2005. Lipid peroxide formation in relation to membrane stability of fresh and frozen-thawed stallion spermatozoa. Mol. Reprod. Dev., 72: 230-238.
CrossRefPubMedDirect Link - Nichi, M., I.G.F. Goovaerts, C.N.M. Cortada, V.H. Barnabe, J.B.P. De Clercq and P.E.J. Bols, 2007. Roles of lipid peroxidation and cytoplasmic droplets on in vitro fertilization capacity of sperm collected from bovine epididymides stored at 4 and 34°C. Theriogenology, 67: 334-340.
CrossRefDirect Link - Purdy, P.H., 2006. A review on goat sperm cryopreservation. Small Rumin. Res., 63: 215-225.
CrossRefDirect Link - Salamon, S. and W.M.C. Maxwell, 2000. Storage of ram semen. Anim. Reprod. Sci., 62: 77-111.
PubMedDirect Link - Sonmez, M. and E. Demirci, 2004. The effect of ascorbic acid and the freezability of ram semen diluted with extenders containing different proportions of glycerol. Turk. J. Anim. Sci., 28: 893-899.
Direct Link - Sundararaman, M.N. and M.J. Edwin, 2003. Semen production traits and freezability of spermatozoa of boer grade goats. Indian. J. Anim. Reprod., 24: 109-112.
Direct Link - Tardiff, A.L., P.B. Farrell, V. Trouern-Trend and R.H. Foote, 1997. Computer assisted sperm analysis for assessing initial semen quality and changes during storage at 5°C. J. Dairy Sci., 80: 1606-1612.
CrossRef - Tuli, R.K. and W. Holtz, 1995. Effect of season on the freezability of Boer goat semen in the Northern temperate zone. Theriogenology, 43: 1359-1363.
CrossRefDirect Link - Van Duijn Jr., C., 1962. Velocity characteristics and numbers of bull spermatozoa in relation to ageing, determined by photo-electric methods. J. Reprod. Fert., 4: 277-290.
CrossRef - Watson, P.F., 1995. Recent developments and concepts in the cryopreservation of spermatozoa and the assessment of their post-thawing function. Reprod. Fertil. Dev., 7: 871-891.
CrossRefPubMedDirect Link - Yeung, C.H., G. Oberiander and T.G. Cooper, 1992. Characterisation of the motility of maturing rat spermatozoa by computer-aided objective measurement. J. Reprod. Fert., 96: 427-441.
CrossRef