
Aris Haryanto
Department of Biochemistry and Molecular Biology, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Jl. Fauna 2, Karangmalang, Yogyakarta, 55281, Indonesia
LiveDNA: 62.12752
Vera Wati
Division of Biotechnology, Animal Disease Investigation Center Wates, Jl. Yogya-Wates Km. 27, Wates, Daerah Istimewa, Yogyakarta, Indonesia
Suhendra Pakpahan
Department of Biochemistry and Molecular Biology, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Jl. Fauna 2, Karangmalang, Yogyakarta, 55281, Indonesia
Nastiti Wijayanti
Laboratory of Animal Physiology, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan, Yogyakarta, 55281, Indonesia
Asian Journal of Animal Sciences
Year: 2016 | Volume: 10 | Issue: 1 | Page No.: 39-48







ABSTRACT
Newcastle disease is an infectious disease in poultry caused by the Newcastle Disease Virus (NDV) which belongs to the family Paramyxoviridae. The NDV can infect various species of birds, both domestic and wild birds with highly morbidity and mortality. Based on the pathotype, NDV can be classified into velogenic (high), mesogenic (moderate) and lentogenic (low) pathotypes, however, clinical symptoms appear on infected birds are often similar that it is difficult to distinguish. In this study, molecular pathotyping of NDV has been done to ten virus isolates that isolated from various type of chicken which naturally infected by NDV using a combination method of RT-PCR and RFLP. It was then compared with quantification of Mean Death Time (MDT) value and DNA sequencing which is commonly method used for NDV pathotyping. The aim of this study was to develop a rapid, precise and efficient method for NDV pathotyping by combination method of RT-PCR amplification and RFLP using HinfI restriction enzyme. The results of NDV pathotyping showed that combination of RT-PCR amplification and RFLP using HinfI enzyme can be used well to differentiate the pathotype of virulent NDV versus avirulent NDV rapidly, accurately and efficiently. A total of ten NDV samples isolated from naturally NDV infected chickens showed that three samples are avirulent NDV, while seven other samples are virulent NDV. In compared with other pathotyping methods, namely MDT quantification and DNA sequencing, the molecular NDV pathotyping by RT-PCR and RFLP methods showed similar conclusion and consistent with highly compatibility.
PDF Abstract XML References Citation
Received: September 21, 2015;
Accepted: October 26, 2015;
Published: November 18, 2015
How to cite this article
Aris Haryanto, Vera Wati, Suhendra Pakpahan and Nastiti Wijayanti, 2016. Molecular Pathotyping of Newcastle Disease Virus from Naturally Infected Chickens by RT-PCR and RFLP Methods. Asian Journal of Animal Sciences, 10: 39-48.
DOI: 10.3923/ajas.2016.39.48
URL: https://scialert.net/abstract/?doi=ajas.2016.39.48
DOI: 10.3923/ajas.2016.39.48
URL: https://scialert.net/abstract/?doi=ajas.2016.39.48
INTRODUCTION
Newcastle Disease (ND) is an infectious viral disease of birds which are acute and easily transmitted. Newcastle disease is caused by a virus which infects various type of bird, both wild and domestic birds, especially chicken (Alexander, 2001). The ND causes high mortality in infected poultry with a mortality rate reaches 100%. It also caused economic loss, especially in the poultry industry, due to the trade restriction and embargo for affected countries by ND outbreak (Keusch et al., 2009).
Newcastle Disease Virus (NDV) is etiological agent of Newcastle disease. It belongs to the family Paramyxoviridae, genus Avulavirus, species of avian paramyxoviridae, serogroup avian paramyxovirus type-1 or APMV-1 (Alexander, 2003). Based on the phylogenetic distance, NDV can be classified into two classes, namely class 1 and class 2. Class 2 NDV are further subclassed into nine genotypes (Susta et al., 2011).
Newcastle Disease Virus (NDV) replicates in the cytoplasm, is an enveloped virus with genetic material of single-stranded (ss) RNA, unsegmented and has a negative polarization. APMV-1 is the synonym of the NDV which has a varied genetic group (Alexander and Senne, 2008). The NDV has a genome size of 15.186 nt (Krishnamurthy and Samal, 1998). The NDV genome consists of 6 genes which encoding 6 major structural proteins, namely nucleoprotein (NP), phosphoprotein (P), matrix protein (M), fusion protein (F), haemagglutinin-neuraminidase (HN) and RNA-dependent RNA polymerase (L). It is conserved gene starts, gene stops and intergenic sequences similar to APMV-1 strains (Tirumurugaan et al., 2011). The F and HN genes are responsible for the infections process and viral pathogenicity (Lamb and Kolakofsky, 1996).
Viral envelope of NDV consists of two transmembrane glycoprotein HN and F proteins. These proteins are shaped spike, that play a role in virus attachment to host cell and they also play an important role in initiating of infection. The HN protein is responsible for the attachment of the virus particles that contain sialic acid on host cells receptors. The HN glycoprotein is the largest molecule that involves haemaglutination activity and neuraminidase (Romer-Oberdorfer et al., 2003). The F protein mediates the fusion of the virion envelope with the cellular membrane plasma. This protein is synthesized into protein precursor (Fo) and activated by proteolytic post-translation process of cleavage site region between amino acids No. 116-117 (Nagai, 1993).
Based on the pathogenicity, NDV is classified into three pathotypes, namely: Lentogenic or NDV with low virulence, mesogenic or NDV with moderate virulence and velogenic, NDV with highly virulence. Although, the NDV is divided into three pathotypes, but the observed clinical symptoms in infected animals are very similar, therefore, it is difficult to distinguish among pathotype only based on the clinical symptoms. Early common symptoms of ND are often found in the naturally infected chickens, such as respiratory disorder, drop in appetite, decreased egg production, as well as disturbances of nerve system. They have similarities with the early clinical symptoms of Avian Influenza (AI) disease (Tabbu, 2000).
There are some methods to determine the pathogenicity of NDV, determining of Index Cerebral Pathogenicity (ICPI) in Day Old Chick (DOC) and Intravenous Pathogenicity Index (IVPI) in 6 weeks old chickens (Tabbu, 2000). However, some scientists argue that ICPI and IVPI methods are not deemed an animal welfare and took a long time (Seal et al., 1995). Conventional NDV pathotyping can also be done by calculating the time spent of virus to kill the chicken embryo, is called by the quantification of Mean Death Time (MDT) value. Highly virulence NDV (velogenic) will kill the chicken embryo within less than 60 h, the moderate virulence NDV (mesogenic) will kill of chicken embryo between 60-90 h, low virulence NDV (lentogenic) takes more than 90 h to kill chicken embryo (Beard and Hanson, 1984). The modern method to distinguish NDV pathogenicity is by RT-PCR method followed by DNA sequencing. Although this method is more accurate but it costs a relatively high (Seal et al., 1995; Marin et al., 1996). Pathotyping method by RT-PCR amplification can be followed by digestion using a restriction enzyme to digest DNA fragment of RT-PCR product (Nanthakumar et al., 2000; Kou et al., 1999; Wehmann et al., 1997).
Development of molecular method to study pathogenesis of NDV have been done by study on the characterization of fusion (F) protein which is a major determinant of NDV pathogenicity. Molecular pathotyping for detection and differentiation of pigeon paramyxovirus serotype-1 (PPMV-1) isolates by RT-PCR and Restriction Enzymes Analysis (REA) have been previously reported by Naveen et al. (2013). The RT-PCR amplification followed by Restriction Fragment Length Polymorphism (RFLP) using restriction enzyme is one of rapid diagnostic method which can be used to determine of NDV pathotype (Heine and Trinidad, 2006; Haryanto et al. 2015). Therefore, the objective of this study was to develop a molecular pathotyping method for NDV that rapidly, accurately and efficiently by using a combination methods of RT-PCR amplification and RFLP using the restriction enzyme Hinfl. Then the results of NDV pathotyping by RT-PCR RFLP, MDT quantification and DNA sequencing were compared to analyze the consistency and compatibility of these NDV pathotyping methods. Thus, it will able to provide useful information in the preparation of mapping strategy, prevention, eradication and control of Newcastle disease spreading in poultry.
MATERIALS AND METHODS
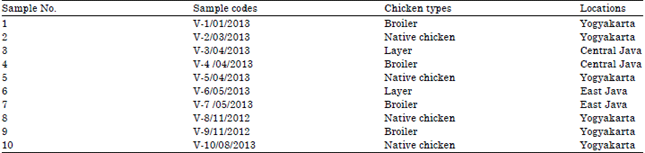
NDV isolates: A total of ten NDV isolates were collected and isolated from naturally infected chicken in various type (broiler, layer, native chicken) derived from some regions in the incidence of Newcastle disease cases in Yogyakarta and Central Java in Indonesia during the period 2012-2013. Samples were collected from cloacal swabs and organs of naturally infected chickens. They were inoculated and propagated in allantoic fluid of embryonated chicken eggs which Specific Antibody Negative (SAN) aged 11 days. Subsequently, it was quantification the MDT value, RNA extraction, RT-PCR amplification, DNA electrophoresis, RFLP by Hinfl enzyme and DNA sequencing.
RNA extraction: Extraction of viral RNA were performed using the High Pure Viral Nucleic Acid Kit (cat. No. 11-858-874-001) according to standard procedures of ROCHE. A total of 200 μL of viral suspension in allantoic fluid was extracted to obtain as many as 50 μL of viral RNA. This NDV RNA to be used as a template for RT-PCR amplification.
RT-PCR amplification: The RT-PCR amplification was performed by SuperScriptTM III One-Step RT-PCR with Platinum kit (cat. No. 12574-026) from Invitrogen. Oligonucleotide primers were used to amplify parts of the NDV fusion (F) gene was designed based on researcher (Kant et al., 1997), as presented in Table 1.
Process of RT-PCR amplification began with the reverse transcriptase reaction at 50°C temperature for 30 min, followed by a process of denaturation at 94°C for 2 min, annealing at 53°C for 30 sec, extension at 68°C for 1 min. This amplification process was ended with the final extension at 68°C for 5 min. The PCR amplification was run in a total of 40 cycles. Then, the RT-PCR products were visualized by electrophoresis on 2% agarose gel stained with fluorosafe 0.5 μg mL–1. The DNA fragments were visualized on a UV Transilluminator at a wavelength of 260 nm in a dark room.
| Table 1: | Sequence of oligonucleotide primers for RT-PCR amplification |
Restriction Fragment Length Polymorphism (RFLP): Hinfl enzyme was used for RFLP in this study, This enzyme was isolated from Haemophilus influenzae Rf; which produced by Thermo scientific in the concentration of 10 U μL–1. Component for RFLP was RT-PCR products as many 2 mL, 6.5 mL dH2O, 10X buffer R as many 1 μL and enzyme HinfI 10 U mL–1 as much as 0.5 mL. The mixture is then incubated at 37°C temperature for 2 h. After the electrophoresed on 3% agarose gel stained with fluorosafe 0.5 μg mL–1. DNA fragments were visualized on a UV Transilluminator appears at a wavelength of 260 nm in a dark room.
DNA sequencing: The DNA sequencing of RT-PCR products to determine the nucleotide sequences was performed in Genetika Science Indonesia Co. by using chain termination method (Sanger method). The DNA sequencing products were then multiple-aligned with F protein encoding genes of NDV database in GenBank using Molecular Evolution Genetics Analysis (MEGA) Program 5.10 version. Results of DNA sequencing were then also to determine the virulent and avirulent NDV. Finally they were compared with the results of RFLP by HinfI enzyme and it was analyzed by using CLC Sequences Viewer 6.8.1 version.
RESULTS AND DISCUSSION
A total of ten samples isolates of NDV that used in this study were isolated from naturally infected chickens. They were collected from various type of chickens during the period of 2012-2013. The list of samples, sample code, the type of chicken and the origin of samples were presented in Table 2.
Identification of NDV began from the inoculation and propagation of virus samples originating from organs and cloacal swabs of naturally infected chickens by NDV into SAN embryonated chicken eggs aged 11 days. The RT-PCR amplification of F protein encoding gene was performed using specific primers corresponding conducted by researcher (Kant et al., 1997). The RT-PCR amplification generated DNA fragment in size of 363 bp. Electrophoresis of RT-PCR products of tenth NDV isolates and positive control of the live ND vaccine (LaSota) as well as a negative control presented in Fig. 1.
Figure 1 showed that the lane M contains marker DNA ladder 100 bp, lane 1-10 sequentially contains sample No. 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10, lane (+) is a positive control of NDV live vaccines (LaSota) and lane (-) is the negative control, namely RT-PCR reagent mixed with RNase-free water. Electrophoresis of RT-PCR products was performed on 2% agarose. In Fig. 1 it is indicated that all the samples were naturally infected by NDV which generates DNA fragments in size of 363 bp, only the last lane containing negative controls which did not seem to generate DNA fragment. This RT-PCR products are similar which study of RT-PCR and RFLP which previously reported by Kant et al. (1997), who used some organs from NDV infected turkey.
| Table 2: | List of NDV isolates which collected from naturally infected chickens |
 | |
| NDV: Newcastle disease virus | |
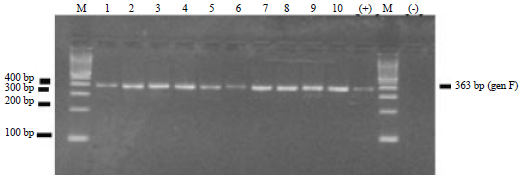 | |
| Fig. 1: | Electrophoresis of RT-PCR product from tenth NDV isolates in size of 363 bp. Lane M: Marker DNA ladder, Lane (+): Positive control, Lane (-): Negative control, Lane 1-10: RT-PCR products |
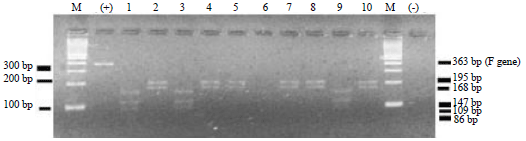 | |
| Fig. 2: | Electrophoresis of RT-PCR products which digested by Hinfl enzyme. Lane M: Marker DNA ladder, Lane (+): Positive control of live NDV vaccine which did not digested by Hinfl enzyme. Lane 1, 3 and 9: RT-PCR products of NDV which digested by Hinfl enzyme into 3 DNA fragments. Lane 2, 4, 5, 6, 7, 8 and 10: RT-PCR products of NDV which digested by Hinfl enzyme into 3 DNA fragments, Lane (-): Negative control |
The RT-PCR of NDV isolates to amplify F gene has been also performed by Haryanto et al. (2013), however, they amplified F gene of NDV directly from field specimens without virus inoculation and propagation process into embryonated chicken eggs.
Then the DNA fragments of RT-PCR products were digested by RLFP using Hinfl restriction enzymes. To determine the NDV pathotype which infects chicken, the pattern of DNA bands after digested by Hinfl were analyze by electrophoresis in 3% agarose gel. The enzymatic digestion by RLFP refers to the method that has been reported by Kou et al. (1999). The DNA fragments formed on 3% agarose gel will be truncated to follow a fixed pattern for virulent and avirulent NDV. The RFLP products from ten NDV isolates were presented more detail in Fig. 2.
The DNA digestion product using Hinfl on tenth NDV isolates showed 2 digestion patterns of DNA fragment. The first digestion pattern shown on NDV isolate No. 1, 3 and 9, while the second digestion pattern shown on sample No. 2, 4, 5, 6, 7, 8 and 10. The NDV isolate No. 1, 3 and 9 generated two DNA fragments in size of 168 and 195 bp respectively, while NDV isolates No. 2, 4, 5, 6, 7, 8 and 10 produced four DNA fragments in size of 21, 86, 109 and 147 bp, respectively. Restriction digest using Hinfl enzyme could distinguish the virulence of NDV with a fixed digestion pattern, because this enzyme digest nucleotide sequence as follow 5'-G/ANTC-3'. Pathotyping to differentiate virulent and avirulent NDV have been done by Kant et al. (1997), who took samples from some organs, such as brain, trachea, spleen and lung from NDV infected turkey.
Pathotyping study to differentiate the virulence of NDV have been carried out by Mase and Kanehira (2012) using Restriction Endonuclease Analysis (REA), which digested the DNA fragments from RT-PCR products using Hin1I and Apa1 enzymes. Other pathotyping study have been conducted by Olav et al. (2005). They reported that not only F gene that plays a role in the virulence of NDV, but the HN protein also plays a role in the NDV virulence. The RFLP method for pathotyping of NDV have been also conducted by Kou et al. (1999), but they used NDV isolated from chickens and owls in Taiwan by using restriction enzymes HinfI, BstOI and RsaI.
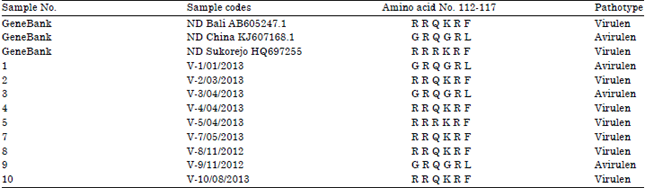
According to Alexander (2003) and Lee et al. (2009), pathogenicity of NDV determined by the base amino acid sequence in the cleavage site area of F protein, especially amino acids sequence No. 112-117. Restriction digest of DNA fragment in F gene by RFLP does not always digest precisely the DNA fragment at the cleavage site region. In this study, Hinfl enzyme digested the DNA fragments on their specific restriction site with a fixed digestion pattern (Berg et al., 2006). Then the each digestion pattern was separated by DNA electrophoresis techniques. They were compared and analyzed based on their difference digestion pattern (Fatchiyah et al., 2011).
On the same NDV isolates, the results of RT-PCR and RFLP were then compared with the result of DNA sequencing. Nucleotide sequence of RT-PCR products were analyzed by multiple alignment. They were compared with the nucleotide sequence of NDV F gene from the GenBank database by using a Molecular Evolution Genetics Analysis (MEGA) program 5.10. version. Results of DNA sequencing in cleavage site area that has been converted to amino acid sequence No. 112-117 was presented in Fig. 3.
In Fig. 3, sample No. 1, 3 and 9 shown that the cleavage site area in the F protein of NDV, amino acid No. 117 is leucine (L). Therefore, the virus can be categorized as avirulent NDV.
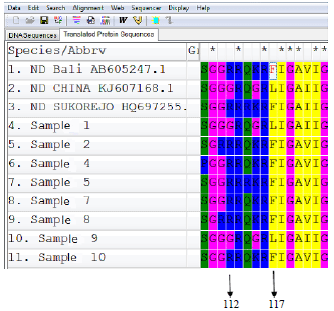 | |
| Fig. 3: | Amino acid sequence No. 112-117 in cleavage site area of NDV F protein |
Sample No. 2, 4, 5, 6, 7, 8 and 10 were categorized as virulent NDV, because the amino acid No. 117 in cleavage site area is phenylalanine (F) as stated by Alexander (1991, 2003), Mase and Kanehira (2012) and Lee et al. (2009). These results are similar with study which reported previously by Haryanto et al. (2015), that determined NDV pathotype by RT-PCR and REA methods, however they used another primers design and others restriction enzymes, namely BamHI and ApaI restriction enzymes.
In addition based on the type of amino acid No. 117 in cleavage site area of NDV F protein, the virulence of NDV viruses can be determined by the presence of multiple basic amino acids in the cleavage site region (amino acid No. 112-117). The more basic amino acids arginine (R) and lysine (K), then the NDV was more virulent (Oberdorfer and Werner, 1998; Ahmadi et al., 2013). (Sequence of amino acid in cleavage site region (No. 112-117) and pathotype of tested NDV isolates were presented in Table 3.
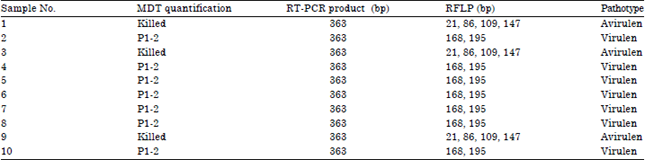
Conventional method for NDV pathotyping was also be done by quantification the Mean Death Time (MDT) value in the tenth tested NDV isolates. Recapitulation of MDT quantification then compared with molecular pathotyping by RT-PCR and RFLP methods (Table 4).
In Table 4, results of MDT quantification for NDV isolates No. 1, 3, 9 shown that for killing embryo in SAN chicken eggs required more than 90 h, so that, the embryos killed intentionally. The NDV isolates No. 2, 4, 5, 6, 7, 8 and 10, virus can killed the embryo in SAN chicken eggs during first passage (P-1) with MDT less than 60 h. Based on the MDT calculation sample No. 1, 3 and 9 can be categorized as avirulent NDV, while sample No. 2, 4, 5, 6, 7, 8 and 10 are virulent NDV. The comparison of MDT quantification and molecular pathotyping by RT-PCR and RFLP showed similar conclusion and consistent with highly compatibility.
| Table 3: | Amino acid sequence and pathotype determination of tested NDV isolates |
 | |
| Table 4: | Data of NDV pathotyping based on MDT quantification and RT-PCR and RFLP |
 | |
MDT: Mean death time, RT-PCR: Reverse transcription polymerase chain reaction, RFLP: Restriction fragment length polymorphism, NDV: Newcastle disease virus | |
CONCLUSION
The overall results of this work showed that the RT-PCR and RFLP methods can be used to determine the NDV pathogenicity rapidly, accurately and efficiently. A total of ten NDV samples isolated from naturally infected chickens showed that the three samples are avirulent NDV, while seven other samples are virulent NDV. In compared with other pathotyping methods, MDT calculation and DNA sequencing, the molecular pathotyping by RT-PCR and RFLP methods for all ten NDV isolates from various type of chickens showed similar conclusion and consistent with a highly compatibility.
ACKNOWLEDGMENTS
We thank to the head office of Animal Disease Investigation Center (ADIC) in Wates, Yogyakarta and the Head of Department of Biochemistry, Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia for the virus isolates, laboratorium facilities and technical supporting to finish this study. This study was supported by Competency Research Grant, DP2M DIKTI, Ministry of Higher Education and Research of Republic Indonesia, academic year 2014-2015 which awarded to Aris Haryanto.
REFERENCES
- Ahmadi, E., M. Ahmadi, S.A. Pourbakhsh and A. Talebi, 2013. Detection and differentiation of newcastle disease virus strains affecting commercial poultry in Northwest of Iran using RT-PCR. Int. J. Vet. Sci., 2: 138-142.
Direct Link - Haryanto, A., M. Purwaningrum, S, Verawati, S.H. Irianingsih and N. Wijayanti, 2015. Pathotyping of local isolates Newcastle disease virus from field specimens by RT-PCR and restriction endonuclease analysis. Procedia Chem., 14: 85-90.
CrossRefDirect Link - Haryanto, A., D. Kristiawan, S.H. Irianingsih and D.W. Yudianingtyas, 2013. [Ampification of fusion protein encoding gene of newcastle disease virus from field specimens by onestep RT-PCR method]. Jurnal Veteriner, 14: 387-393, (In Indonesian).
Direct Link - Kant, A., G. Koch, D.J. Van Roozelaar, F. Balk and A. Ter Huurne, 1997. Differentiation of virulent and non‐virulent strains of Newcastle disease virus within 24 hours by polymerase chain reaction. Avian Pathol., 26: 837-849.
CrossRefDirect Link - Krishnamurthy, S. and S.K. Samal, 1998. Nucleotide sequences of the trailer, nucleocapsid protein gene and intergenic regions of Newcastle disease virus strain Beaudette C and completion of the entire genome sequence. J. Gen. Virol., 79: 2419-2424.
CrossRefPubMedDirect Link - Lee, E.K., W.J. Jeon, J.H. Kwon, C.B. Yang and K.S. Choi, 2009. Molecular epidemiological investigation of Newcastle disease virus from domestic ducks in Korea. Vet. Microbiol., 134: 241-248.
CrossRefDirect Link - Marin, M.C., P. Villegas, J.D. Bennett and B.S. Seal, 1996. Virus characterization and sequence of the fusion protein gene cleavage site of recent newcastle disease virus field isolates from the Southeastern United States and Puerto Rico. Avian Dis., 40: 382-390.
CrossRefDirect Link - Mase, M. and K. Kanehira, 2012. Simple differentiation of avirulent and virulent strains of avian paramyxovirus serotype-1 (Newcastle disease virus) by PCR and restriction endonuclease analysis in Japan. J. Vet. Med. Sci., 74: 1661-1664.
CrossRefDirect Link - Nagai, Y., 1993. Protease-dependent virus tropism and pathogenicity. Trends. Microbiol., 1: 81-87.
CrossRefPubMedDirect Link - Nanthakumar, T., R.S. Kataria, A.K. Tiwari, G. Buchaiah and J.M. Kataria, 2000. Pathotyping of Newcastle disease viruses by RT-PCR and restriction enzyme analysis. Vet. Res. Commun., 24: 275-286.
CrossRefPubMedDirect Link - Naveen, K.A., S.D. Singh, J.M. Kataria, R. Barathidasan and K. Dhama, 2013. Detection and differentiation of pigeon paramyxovirus serotype-1 (PPMV-1) isolates by RT-PCR and restriction enzyme analysis. Trop. Anim. Health Prod., 45: 1231-1236.
CrossRefPubMedDirect Link - Oberdorfer, A. and O. Werner, 1998. Newcastle disease virus: Detection and characterization by PCR of recent German isolates differing in pathogenicity. Avian Pathol., 27: 237-243.
CrossRefPubMedDirect Link - Olav, S., O.S. Deleeuw, G.L. Hantog, N. Ravenshost and B.P. Peeters, 2005. Virulence of Newcastle disease virus is determined by the cleavage site of the fusion protein and by both the stem region and globular head of the haemagglutinin-neuraminidase protein. J. Gen. Virol., 86: 1759-1769.
CrossRefDirect Link - Romer-Oberdorfer, A., O. Werner, J. Veits, T. Mebatsion and T.C. Mettenleiter, 2003. Contribution of the length of the HN protein and the sequence of the F protein cleavage site to Newcastle disease virus pathogenicity. J. General Virol., 84: 3121-3129.
CrossRefDirect Link - Seal, B.S., D.J. King and J.D. Bennett, 1995. Characterization of Newcastle disease virus isolates by reverse transcription PCR coupled to direct nucleotide sequencing and development of sequence database for pathotype prediction and molecular epidemiological analysis. J. Clin. Microbiol., 33: 2624-2630.
Direct Link - Susta, L., P.J. Miller, C.L. Afonso and C.C. Brown, 2011. Clinicopathological characterization in poultry of three strains of Newcastle disease virus isolated from recent outbreaks. Vet. Pathol., 48: 349-360.
CrossRefDirect Link - Kou, Y.T., L.L. Chueh and C.H. Wang, 1999. Restriction fragment length polymorphism analysis of the f gene of newcastle disease viruses isolated from chickens and an owl in Taiwan. J. Vet. Med. Sci., 61: 1191-1195.
CrossRefDirect Link - Tirumurugaan, K.G., S. Kapgate, M.K. Vinupriya, K. Vijayarani, K. Kumanan and S. Elankumaran, 2011. Genotypic and pathotypic characterization of newcastle disease virus from India. PLoS ONE, Vol. 6.
CrossRefDirect Link - Wehmann, E., J. Herczeg, A. Ballagi-Pordany and B. Lomniczi, 1997. Rapid identification of Newcastle disease virus vaccine strains LaSota and B-1 by restriction site analysis of their matrix gene. Vaccine, 15: 1430-1433.
CrossRefDirect Link