
Amri Ismail
Faculte des Sciences de Bizerte, Bizerte, 7021, Tunisie
Hamrouni Lamia
Laboratoire d`Ecologie et d`Amelioration Sylvo-Pastorale, Institut National de Recherches en Genie Rural Eaux et Forets, P.B. 10, 2080 Ariana, Tunisie
Hanana Mohsen
Centre de Biotechnologie de Borj-Cedria, BP 901, Hammam-lif 2050, Tunisie
Jamoussi Bassem
Institut Superieur d�Education et de Formation Continue, Tunis, Tunisie
Asian Journal of Applied Sciences
Year: 2011 | Volume: 4 | Issue: 8 | Page No.: 771-779







ABSTRACT
In this study, the chemical composition of the essential oils isolated from leaves of Juniperus oxycedrus L. subsp. macrocarpa were analyzed and their phytotoxic effects were evaluated. All together, 41 compounds were identified, accounting for 96% of the total oil. The monoterpenes fraction amounted to 61% of the oil and was characterized by a high proportion of monoterpene hydrocarbons (55%), amongst which the most abundant was α-pinene (42%) and sesquiterpenoids (29%). Regarding the herbicidal activity, the oil completely inhibited seed germination and seedling growth of three common weeds: Phalaris paradoxa, Trifolium campestre and Lolium rigidum and the activity by direct contact was more effective than by fumigation. The findings of the present study suggest a possible use of these essential oils as natural herbicides.
PDF Abstract XML References Citation
Received: July 31, 2011;
Accepted: October 13, 2011;
Published: November 01, 2011
How to cite this article
Amri Ismail, Hamrouni Lamia, Hanana Mohsen and Jamoussi Bassem, 2011. Chemical Composition of Juniperus oxycedrus L. subsp macrocarpa Essential Oil and Study of Their Herbicidal Effects on Germination and Seedling Growth of Weeds. Asian Journal of Applied Sciences, 4: 771-779.
DOI: 10.3923/ajaps.2011.771.779
URL: https://scialert.net/abstract/?doi=ajaps.2011.771.779
DOI: 10.3923/ajaps.2011.771.779
URL: https://scialert.net/abstract/?doi=ajaps.2011.771.779
INTRODUCTION
Secondary metabolites are diverse compounds with a variety of functions, playing an important role in plant-plant, plant-insect and plant-pathogen interactions (Koudou et al., 2008; Shonouda et al., 2008; Razavi, 2011). Several phytochemicals, extracted from various botanical sources, have been reported to have detrimental effects on weeds, bacteria, fungi and insects (Ghasemi et al., 2007; Nour et al., 2009). Plant secondary metabolites in general have been recognized as an important natural source for weeds management. Several studies have documented that plant extracts and volatile oils inhibit and delay seed germination and inhibit seedling growth of many weeds and cultivated crops (Salam and Kato-Noguchi, 2010; Kordali and Atabeyoglu, 2006). The studies of allelopathic interactions between plants are one of the most interesting strategies for herbicide discovery. The use of secondary metabolites implicated in allelopathic interactions as sources for new agrochemical models could satisfy the requirements for crop protection and weeds management (Singh et al., 2003; Kolahi et al., 2009). These studies are important in view of the environmental and human health concerns of chemical weed control and increasing resistance in weeds caused by synthetic herbicides (Buriro et al., 2003); thus, there is a need to search for environmentally safer compounds.
In the literature, the herbicidal effects of essential oils from Lamiaceae, Anacardiaceae, Verbenaceae, Rutaceae, Asteraceae, Myrtaceae and other family against weeds have been previously reported (Kordali and Atabeyoglu, 2006; Kil et al., 2000; De Feo et al., 2002; Angelini et al., 2003; Kordali et al., 2008; Verdeguer et al., 2009) but nothing was yet reported on the phytotoxic effects of members of the Cupressaceae family like Juniperus oxycedrus L. subsp. macrocarpa which is a small tree growing in stony places of Mediterranean and Near East countries. J. oxycedrus is one of the most appreciated plants in folk medicine for its essential oil richness and its plethora of biologically active compounds. The leaves of J. oxycedrus are commonly used as a spicy and aromatic flavouring for soups. They are also widely used in folk medicine to treat various disease treatments, such as hyperglycemia, obesity, tuberculosis, bronchitis and pneumonia (de Medina et al., 1994). There are many reports on the chemical composition of the oils from Juniperus species (Adams et al., 1999; Milos and Radonic, 2000; Salido et al., 2002; Loizzo et al., 2007). Most of these reports indicate that α-pinene, manoyl oxide and z-caryophyllene are the main constituents of these oils. The chemical composition of Tunisian J. oxycedrus have been reported (Medini et al., 2010), however, there are no reports available on the phytotoxic potential of J. oxycedrus essential oils. Therefore, the aims of this study were to determine the chemical composition of essential oils extracted from the aerial parts of Juniperus oxycedrus subsp. macrocarpa, to assess their herbicidal activity via air and direct contact on germination and seedling growth of three different weed species and finally, to relate the biological effects observed to the chemical composition.
MATERIALS AND METHODS
Plant material: Fresh leaves of Juniperus oxycedrus subsp. macrocarpa were collected in October 2009 from an experimental field (Seliana, Tunisia) of the national Institute for Research in rural Engineering, water and forest. The experimental site is located in the makther region at an altitude of 865 m, where an upper semi arid climate prevails. The specimen of the plant was submitted to the herbarium division of the Institute and identification was performed in the Laboratory of Genetic.
Isolation of the essential oils: One hundred grams of air-dried (10 days at 25°C) and finely grounded raw materials were submitted to hydro distillation for 5 h with 500 mL distilled water using a Clevenger type apparatus. The oil obtained was collected and dried over anhydrous sodium sulfate and stored in sealed glass vials in a refrigerator at 4°C until analysis. Yield based on dried weight of the sample was calculated (w/w %).
Gas chromatography and mass spectrometry analysis
Gas chromatography analysis with FID detection: The essential oils were analysed using a Hewlett Packard 5890 II GC equipped with Flame Ionization Detector (FID) and HP-5 MS capillary column (5% phenyl/95% dimethyl polysiloxane: 30 mx0.25 mm id, film thickness 0.25 μm). Injector and detector temperature were set at 250 and 280°C, respectively. Oven temperature was kept at 50°C for 1 min then gradually raised to 250°C at 5°C min-1 and subsequently, held isothermal for 4 min. Nitrogen was the carrier gas at a flow rate of 1.2 mL min-1. Diluted samples (1/100 in hexane, v/v) of 1.0 μL were injected manually in the splitless mode. Quantitative data were obtained electronically from FID area percent data without using correction factors.
Gas chromatography analysis with MS detection: Analysis of the oils was performed using a Hewlett Packard 5890 II GC, equipped with a HP 5972 mass selective detector and a HP-5 MS capillary column (30 mx0.25 mm id, film thickness 0.25 μm). For GC/MS detection, an electron ionization system, with ionization energy of 70 eV, a scan time of 1.5 s and mass range 40-300 amu, was used. Helium was the carrier gas at a flow rate of 1.2 mL min-1. Injector and transfer line temperatures were set at 250 and 280°C, respectively. The oven temperature program was the same with GC/FID analysis. Diluted samples (1/10 in hexane, v/v) of 1.0 μL were injected manually in the splitless mode. The identification of the compounds was based on mass spectra (compared with Wiley 275.L, 6th edition mass spectral library) or with authentic compounds and confirmed by comparison of their retention indices either with those of authentic compounds or with data published in the literature as described by Adams (2001). Further confirmation was done from retention index data generated from a series of n-alkanes retention indices (relative to C9-C28 on the HP-5 MS capillary column).
Seed germination and seedling growth experiments: Mature seeds of the annual weeds Phalaris paradoxa, Trifolium campestre and Lolium rigidum were obtained from parent plants collected in fields of Tunisia in July 2009. The plants were dried for 15 days at room temperature before the seeds were extracted. Uniform, healthy seeds were selected and stored at 4°C until used. Prior to use, seeds were sterilized with 15% sodium hypochlorite for 20 min. They were then rinsed with distilled water. Empty and undeveloped seeds were discarded by floating in tap water and the remaining seeds were used. To determine the herbicidal effects of vapors of the essential oil, 20 sterilized seeds were distributed evenly on two layers of filter paper in sterile Petri dishes (9 cm). Subsequently, seeds and filter paper were moistened with 6 mL of distilled water (includes 0.1% Tween-20). The essential oil was dripped on a paper placed on the lid of Petri dish using a micro pipette to obtain final concentrations of 0, 3, 6 and 10 mg oil/Petri dish. Petri dishes were closed with an adhesive tape to prevent escaping of volatile compounds and kept at 25°C in a growth chamber with 12 h of fluorescent light (Dudai et al., 1999; Kordali et al. 2008).
After 10 days the number of germinated seeds and seedling lengths were measured. Germination was measured as the percentage of seeds from which a radical emerges.
To determine the contact herbicidal effect of the essential oil, the oil was dissolved in a tween-water solution (0.1%; v/v). An aliquot of 6 mL of the essential oil solution was transferred to Petri dishes equipped with two layers of filter paper to obtain final concentrations of the treatments 0, 3, 6 and 10 mg/Petri dish. Afterwards, 20 seeds of Phalaris paradoxa, Trifolium campestre and Lolium rigidum were added and distributed evenly on the filter paper. Petri dishes were closed with an adhesive tape to prevent escaping of volatile compounds and were kept at 25°C in a growth chamber with 12 h of fluorescent light (Dudai et al., 1999). The number of germinated seeds and seedling lengths were measured after 10 days. The treatments were arranged in a completely randomized design with three replications including controls.
Statistical analysis: Data obtained from essential oil analysis, seed germination and seedling growth assays were expressed as mean values and subjected to one-way analysis of variance (ANOVA), using SPSS 13.0 software package. Differences between means were tested through Student-Newman-Keuls test (SNK) and values of p≤0.05 were considered significantly different (Sokal and Rohlf, 1995).
RESULTS AND DISCUSSION
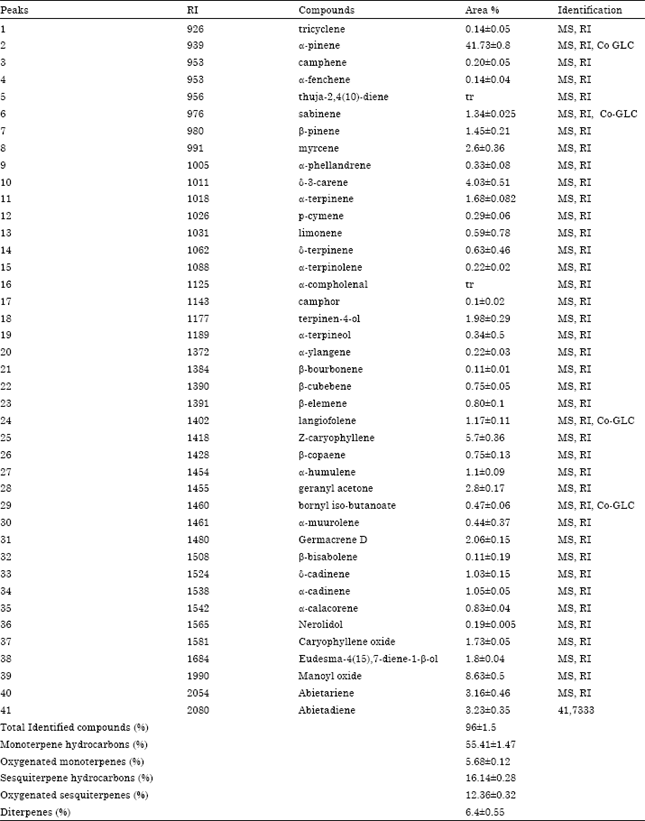
Chemical composition: The volatile components in the essential oils isolated from the leaves of Juniperus oxycedrus and their percentage are shown in Table 1. The hydrodistillation yielded 0.21% (w/w) of yellow oil (on a dry mass basis). Forty one compounds were identified, accounting for 96% of the total oil. J. oxycedrus oil was found to be rich in α-pinene (42%), manoyl oxide (9%) and z-caryophyllene (6%). The monoterpenoid fraction amounted to 61% of the oil and was characterized by a high proportion of monoterpene hydrocarbons (55%), amongst which the most abundant was α-pinene (42%). The sesquiterpenoid fraction (29%) was mainly composed of sesquiterpenoid hydrocarbons (16%), with z-caryophyllene (6%) being the main compound. Manoyl oxide (6%) was the most abundant of the six oxygenated sesquiterpenoids that represented 12% of the total oil. Diterpenes content was 6% with abietariene and abietadiene as the two main compounds. The essential oil of J. oxycedrus was previously investigated. In agreement with our result, most of studies indicated that α-pinene, manoyl oxide and z-caryophyllene were the major components (Adams et al., 1999; Milos and Radonic, 2000; Salido et al., 2002; Loizzo et al., 2007; Medini et al., 2010). Some differences were detected in our oil composition and yield which may be due to several factors such as time of collection, age of plant material collected, genetics, geographic and climatic conditions.
Herbicidal effects of the oil on weed germination and seedling growth: Essential oils are a complex mixture of volatile constituents which are characterized by its volatility. Thus, their phytotoxic effects may be exercised by two modes, a direct contact and the second was the vapors of oil. For these reasons and based on the literature (Kordali et al., 2008). We studied the phytotoxic effect of Juniperus oil via air and in solution on germination and seedling growth of L. rigidium, P. paradoxa and T. campestre.
Our results showed that the oil significantly inhibited the germination and seedling growth of all tested weeds. J. oxycedrus oil strongly inhibited the germination in a dose dependent manner with the effect being significantly more effective with direct contact than via air (Table 2). In fact, germination of all tested weeds was significantly reduced at lower doses (3 and 6 mg/Petri dish), at the dose of 6 mg/Petri dish, the germination of L. rigidium, P. paradoxa and T. campestre were reduced, respectively by 80, 90 and 76% (direct contact) and by 62, 48 and 31% (vapors of oil). At the dose of 10 mg/Petri dish, germination of all weeds was totally inhibited only with direct contact at the highest dose while at the same dose, when weeds were exposed to the vapors of oils; their germination was reduced to 8.33, 35 and 36.66%, respectively for (L. rigidium, P. paradoxa and T. campestre).
Results presented in Table 3 and 4 show that seedling growth of all weeds was affected by the application of the oil, in fact, seedling growth of all weeds were reduced compared to control, on the other hand, the inhibitory effects of the oils were dependent on the dose used, tested weeds and method of application (direct contact or via air). At the dose of 6 mg/Petri dish, roots growth of L. rigidium, P. paradoxa and T. campestre were reduced, respectively by 80, 71 and 85% (direct contact) and by 47, 18 and 44% (vapors of oil). While, at the dose of 10 mg/Petri dish, their growth was totally inhibited only with direct contact but, at the same dose and with vapors of oil, roots growth was only reduced by 58, 33 and 70%, respectively for (L. rigidium, P. paradoxa and T. campestre). Similarly, shoots growth of all weeds was significantly reduced at all doses and totally inhibited at the dose of 10 mg/Petri dish on the direct contact application.
| Table 1: | Chemical composition of Juniperus oxycedrus L. essential oil |
 | |
| RI: Retention index; MS: Mass spectrometry; Co-GLC: Co-injection; tr: Trace (< 0.1%). %: Mean percentage of three independent replicates calculated by GC-FID on non-polar HP-5 capillary column | |
| Table 2: | Inhibitory effects of essential oils of J. oxycedrus on germination of weeds |
 | |
| Means in the same column with the same letter are not significantly different at p≤0.05 | |
| Table 3: | Inhibitory effects of essential oils of J. oxycedrus on aerial parts growth of weeds (mm) |
 | |
| Means in the same column with the same letter are not significantly different at p≤0.05 | |
| Table 4: | Inhibitory effects of essential oils of J. oxycedrus on roots growth of weeds (mm) |
 | |
| Means in the same column with the same letter are not significantly different at p≤0.05 | |
Our data agree with literature reports on the inhibitory activity exerted by essential oils on weeds germination and seedling growth (Kordali and Atabeyoglu, 2006; Singh et al., 2006). It has been shown that the herbicidal effects of essential oils resulted from the combined effect of several constituents exerting additive, synergetic or antagonistic effects (Kordali and Atabeyoglu, 2006; Kordali et al., 2009). Thus, putting in evidence the compound(s) responsible for the activity of the oil, it is necessary to test pure compounds and the various possible combinations of them. Several studies have tested the activity of pure and combined compounds such as limonene, α-pinene, decanone, z-caryophyllene, nonanol, nonanone, terpinen-4-ol, bornyl acetate, citronellal, camphor, carvacrol, thymol, geraniol, menthol and borneol (Kil et al., 2000; De Feo et al., 2002; Kordali and Atabeyoglu, 2006; Singh et al., 2006; Kordali et al., 2009; Wang et al., 2009). In addition, in our study, the oil was rich in monoterpenes, especially α-pinene and the sesquiterpenes z-caryophyllene and manoyl oxide which are known for their phytotoxic effects (Kil et al., 2000; De Feo et al., 2002; Singhet al., 2006; Wang et al., 2009). Numerous studies showed the phytotoxic potential of essential oil isolated from different plants. Scrivanti et al. (2003) demonstrated that the oil isolated from Schinus areira and its major compound α-pinene (85 %) inhibited Zea mays root growth.
Singh et al. (2006) has demonstrated that exposure of seedling to α-pinene inhibited seedling growth by causing oxidative damage in root tissue. Kil et al. (2000) reported that z-caryophyllene was an important sesquiterpene of essential oil of Artemisia lavandulaefolia which suppressed the seedling growth of Achyranthes japonica. Wang et al. (2009) showed that z-caryophyllene at a dose of 3 mg L-1 significantly inhibited the germination and seedling growth of Brassica campestris and Raphanus sativus. Generally, both monoterpenoids and sesquiterpenoids appear to be involved in these phytotoxic effects of the oil, for this reason the herbicidal activity of our oil was attributed to the presence of both sesquiterpenes and monoterpenes and the synergism between components does play an important role.
Although the exact mechanisms of essential oil effects on germination and seedling growth inhibition remain unclear, such inhibitory effects could be caused by allelochemicals interfering with physiological and biochemical processes in target species (Weir et al., 2004). Indeed, it has been reported that the inhibition of germination may be the consequence of the inhibition of water uptake (Gniazdowska and Bogatek, 2005).
Weir et al. (2004) have reported that allelochemicals caused an increasing of abscisic Acid (ABA) content, decreased indole-3-acetic acid (IAA) and Zeatin Riboside (ZR) contents. On the other hand, previous studies showed that essential oils cause many changes in plant seedlings leading to reduction in some organelles such as mitochondria and inhibition of DNA synthesis (Koitabashi et al., 1997; Zunino and Zygadlo, 2004; Nishida et al., 2005). One or more of these factors could explain the blockage of the seed germination and the death of embryon.
Like other abiotic environmental stresses such as salt and drought, it has been documented that essential oils and their individual components, enhanced loss of membrane integrity by increasing the generation of reactive oxygen species.
In addition, oxidative stress induced by essential oils was evidenced by increased activity of antioxidant enzymes activities, like superoxide dismutase, ascorbate peroxidase, guaiacol peroxidase, catalase and glutathione reductase which are potent scavengers of reactive oxygen species (Maness et al., 1999; Weir et al., 2004; Singh et al., 2006). Abrahim et al. (2003) have demonstrated that α-pinene acts on the seedling growth of Zea mays by two mechanisms: uncoupling of oxidative phosphorylation and inhibition of electron transfer which result the uncoupling of mitochondrial energy metabolism and inhibition of the mitochondrial ATP production. In the same report, it demonstrates that the actions of α-pinene on isolated mitochondria are consequences of unspecific disturbances in the inner mitochondrial membrane.
CONCLUSIONS
The essential oil of Juniperus oxycedrus obtained by hydrodistillation was mainly composed of α- and β-pinene, z-caryophyllene and manoyl oxide. Our results showed that J. oxycedrus essential oil displayed a phytotoxic effect against dicot and monocot weeds. According to our knowledge, this is the first report regarding J. oxycedrus essential oil herbicidal activity. The development of natural pesticides would help to decrease the negative impact of synthetic agents, such as residues, resistance and environmental pollution. In this respect, essential oils, as natural herbicides, present two mains characters: the first is their natural origin which means more safety to the people and the environment and the second is that they have be considered at low risk for resistance development by weeds. It is believed that it is difficult to develop resistance to such a mixture of oil components with apparently different mechanisms of action. Based on our preliminary results, the essential oils of J. oxycedrus could be suggested as alternative herbicides. However, further studies are required to determine the cost, applicability, safety and phytotoxicity against the cultured plants of these agents as potential herbicide.
REFERENCES
- Abrahim, D., A.C. Francischini, E.M. Pergo, A.M. Kelmer-Bracht and E.L. Ishii-Iwamoto, 2003. Effects of α-pinene on the mitochondrial respiration of maize seedlings. Plant Physiol. Biochem., 41: 985-991.
CrossRef - Adams, R.P., J. Altarejos, C. Fernandez and A. Camacho, 1999. The leaf essential oils and taxonomy of Juniperus oxycedrus L. subsp. Oxycedrus, subsp. badia (H. Gay) debeaux and subsp. Macrocarpa (Sibth. & Sm.) Bali. J. Essent. Oil Res., 11: 167-172.
Direct Link - Angelini, L.G., G. Carpanese, P.L. Cioni, I. Morelli, M. Macchia and G. Flamini, 2003. Essential oils from Mediterranean Lamiaceae as weed germination inhibitors. J. Agric. Food Chem., 51: 6158-6164.
PubMed - Kordali, Y.B. and O. Atabeyoglu, 2006. The allelopathic effect of pictacia leaf extracts and pure essential oil components on pelargonium ringo deep scarlet F1 hybrid seed germination. J. Applied Sci., 6: 2040-2042.
CrossRefDirect Link - Buriro, U.A., F.C. Oad, S.K. Agha and G.S. Solangi, 2003. Post emergence weed control in wheat. J. Appl. Sci., 3: 424-427.
CrossRefDirect Link - De Feo, V., F. de Simone and F. Senatore, 2002. Potential allelochemicals from the essential oil of Ruta graveolens. Phytochemistry, 61: 573-578.
PubMed - De Medina, F.S., M.J. Gamez, I. Jiminez, J. Jimenez, J.I. Osuna and A. Zarzuelo, 1994. Hypoglycemic activity of Juniper Berries. Planta Med., 60: 197-200.
PubMedDirect Link - Dudai, N., A. Poljakoff-Mayber, A.M. Mayer, E. Putievsky and H.R. Lerner, 1999. Essential oils as allelochemicals and their potential use as bioherbicides. J. Chem. Ecol., 25: 1079-1089.
CrossRefDirect Link - Ghasemi, Y., A. Khalaj, A. Mohagheghzadeh, A.R. Khosravi and M.H. Morowvat, 2007. Composition and antimicrobial activity of the essential oil and extract of Hypericum elongatum. J. Appl. Sci., 7: 2671-2675.
CrossRefDirect Link - Kil, B.S., D.M. Han, C.H. Lee, Y.S. Kim, K.Y. Yun and H.G. Yoo, 2000. Allelopathic effects of Artemisia lavandulaefolia. Korean J. Ecol., 23: 149-155.
Direct Link - Kordali, S., A. Cakir, H. Ozer, R. Cakmakci, M. Kesdek and E. Mete, 2008. Antifungal, phytotoxic and insecticidal properties of essential oil isolated from Turkish Origanum acutidens and its three components, carvacrol, thymol and p-cymene. Bioresour. Technol., 99: 8788-8795.
CrossRefDirect Link - Kordali, S., A. Cakir, T.A. Akcin, E. Mete, A. Akcin, T. Aydin and H. Kilic, 2009. Antifungal and herbicidal properties of essential oils and n-hexane extracts of Achillea gypsicola Hub-Mor. and Achillea biebersteinii Afan. (Asteraceae). Ind. Crops Prod., 29: 562-570.
CrossRefDirect Link - Koitabashi, R., T. Suzuki, T. Kawazu, A. Sakai, H. Kuroiwa and T. Kuroiwa, 1997. 1,8-cineole inhibits root growth and DNA synthesis in the root apical meristem of Brassica campestris L. J. Plant Res., 110: 1-6.
CrossRef - Kolahi, M., B. Peivastegan, I. Hadizadeh and S.M. Seyyednejad, 2009. Inhibition of germination and seedling growth of wild oat by rice hull extracts. J. Applied Sci., 9: 2857-2860.
CrossRefDirect Link - Koudou, J., P. Edou, L.C. Obame, I.H. Bassole and G. Figueredo et al., 2008. Volatile components, antioxidant and antimicrobial properties of the essential oil of Dacryodes edulis G. don from gabon. J. Applied Sci., 8: 3532-3535.
CrossRefDirect Link - Loizzo, M.R., R. Tundis, F. Conforti, A. Saab, G. Statti and F. Menichini, 2007. Comparative chemical composition, antioxidant and hypoglycaemic activities of Juniperus oxycedrus ssp. Oxycedrus L. berry and wood oils from Lebanon. Food Chem., 105: 572-578.
CrossRef - Maness, P.C., S. Smolinski, D.M. Blake, Z. Huang, E.J. Wolfrum and W.A. Jacoby, 1999. Bactericidal activity of photocatalytic TiO2 reaction: Toward an understanding of its killing mechanism. Applied Environ. Microbiol., 65: 4094-4098.
PubMed - Milos, M. and A. Radonic, 2000. Gas chromatography mass spectral analysis of free and glycosidically bound volatile compounds from Juniperus oxycedrus L. growing wild in Croatia. Food Chem., 68: 333-338.
CrossRef - Shonouda, M.L., S. Osman, O. Salama and A. Ayoub, 2008. Insecticidal Effect of Chrysanthemum coronarium L. flowers on the pest Spodoptera littoralis boisd and its parasitoid Microplitis rufiventris Kok. with identifying the chemical composition. J. Applied Sci., 8: 1859-1866.
CrossRefDirect Link - Nishida, N., S. Tamotsu, N. Nagata, C. Saito and A. Sakai, 2005. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. J. Chem. Ecol., 31: 1187-1203.
CrossRefDirect Link - Nour, A.H., S.A. Elhussein, N.A. Osman, N.E. Ahmed, A.A. Abduelrahman, M.M. Yusoff and A.H. Nour, 2009. Antibacterial activity of the essential oils of sudanese accessions of Basil (Ocimum basilicum L.). J. Applied Sci., 9: 4161-4167.
CrossRefDirect Link - Razavi, S.M., 2011. Plant coumarins as allelopathic agents. Int. J. Biol. Chem., 51: 86-90.
CrossRefDirect Link - Salam, M.A. and H. Kato-Noguchi, 2010. Allelopathic potential of methanol extract of bangladesh rice seedlings. Asian J. Crop Sci., 2: 70-77.
CrossRefDirect Link - Salido, S., J. Altarejos, M. Nogueras, A. Sanchez, C. Pannecouque, M. Witvrouw and E. De Clercq, 2002. Chemical studies of essential oils of Juniperus oxycedrus ssp. Badia. J. Ethnopharmacol., 81: 129-134.
CrossRef - Scrivanti, L.R., M.P. Zunino and J.A. Zygadlo, 2003. Tagetes minuta and Schinus areira essential oils as allelopathic agents. Biochem. Syst. Ecol., 31: 563-572.
CrossRefDirect Link - Singh, H.P., D.R. Batish, S. Kaur, K. Arora and R.K. Kohli, 2006. α-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot., 98: 1261-1269.
CrossRefDirect Link - Singh, H.P., D.R. Batish and R.K. Kohli, 2003. Allelopathic interactions and allelochemicals: New possibilities for sustainable weed management. Critic. Rev. Plant Sci., 22: 239-311.
CrossRefDirect Link - Verdeguer, M., M.A. Blazquez and H. Boira, 2009. Phytotoxic effects of Lantana camara, Eucalyptus camaldulensis and Eriocephalus africanus essential oils in weeds of mediterranean summer crops. Biochem. Syst. Ecol., 37: 362-369.
CrossRef - Wang, R., S. Peng, R. Zeng, L.W. Ding and Z. Xu, 2009. Cloning, expression and wounding induction of β-caryophyllene synthase gene from Mikania micrantha H.B.K. and allelopathic potential of β-caryophyllene. Allelopathy J., 24: 35-44.
Direct Link - Weir, T.L., S.W. Park and J.M. Vivanco, 2004. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant Biol., 7: 472-479.
CrossRefDirect Link - Zunino, M.P. and J.A. Zygadlo, 2004. Effect of monoterpenes on lipid oxidation in maize. Planta, 219: 303-309.
PubMed
pawan Reply
what chemicals can i use to make curry more sour