
Hemangi S. Bhonsle
Mass Spectrometry and Proteomics Group, Division of Organic Chemistry, National Chemical Laboratory, Pune, India
Mahesh J. Kulkarni
Mass Spectrometry and Proteomics Group, Division of Organic Chemistry, National Chemical Laboratory, Pune, India
Trends in Molecular Sciences
Year: 2009 | Volume: 1 | Issue: 1 | Page No.: 11-17







ABSTRACT
The role of molecular mass of protein in determining glycation was addressed in this study. Large molecular mass proteins such as IgG, HSA and BSA, upon glycation showed higher increase in mass compared to small molecular mass proteins such as papain, apomyoglobin and insulin. Also, the extent of glycation was found to be more in HSA, BSA and IgG compared to papain, apomyoglobin and insulin. This study combined with earlier study on albumin glycation suggests that, in addition to the protein structure and turnover, the molecular mass of protein as well as protein abundance determines the glycation.
PDF Abstract XML References Citation
How to cite this article
Hemangi S. Bhonsle and Mahesh J. Kulkarni, 2009. Influence of Protein Molecular Mass on the Glycation. Trends in Molecular Sciences, 1: 11-17.
DOI: 10.3923/tms.2009.11.17
URL: https://scialert.net/abstract/?doi=tms.2009.11.17
DOI: 10.3923/tms.2009.11.17
URL: https://scialert.net/abstract/?doi=tms.2009.11.17
INTRODUCTION
Microvascular complications such as nephropathy, retinopathy, neuropathy etc., are mainly developed due to a non-enzymatic reaction between glucose and protein termed as glycation (Brownlee, 2001). Glycation triggers a cascade of events leading to formation of Advanced Glycation End (AGEs) products. Many studies have suggested that formation of AGEs may be important factor involved in the etiology of diabetic complications (Thornalley, 2003). Biological activity of many proteins like insulin, crystallin, glucose-6-phosphate dehydrogenase, aldehyde reductase, glutathione reductase, Cu-Zn superoxide dismutase, etc., was affected because of glycation process (Hunter et al., 2003). This process has been well demonstrated both in vitro and in vivo in several model proteins like hemoglobin, BSA, HSA, IgG, insulin etc. and these proteins differ in their extent of glycation (Lapolla et al., 2006). As glycation is a non-enzymatic process, theoretically all proteins should get glycated; however, several studies suggest that certain proteins are more prone to undergo glycation than others. For example, glycoxidative modifications have been shown to be targeted only to restricted set of proteins using two-dimensional gel electrophoresis followed by western blotting with anti-AGE antibody (Poggioli et al., 2002). In another study, when mouse serum incubated with 3-deoxyglucosone results in the loss of the activity of the specific enzymes responsible for glucose metabolism. The activity of hexokinase and glucose-6-phosphate dehydrogenase activities were decreased strongly, while glucokinase, glucose-6-phosphatase and phosphofructokinase activities were scarcely affected (Kiho et al., 1996). A recent study demonstrates the glycation of vimentin is based on the structural properties of the protein but not on turnover or high intracellular gene expression level (Kueper et al., 2007). Recent study shows that abundant proteins determine glycation of less abundant proteins (Bhonsle et al., 2008). In this study, albumin the most abundant plasma protein competitively inhibits the glycation of less abundant proteins. Also, it was observed that the extent of glycation was more for a high molecular mass protein (albumin) compared to a low molecular mass protein (insulin) indicates that molecular mass of the protein is one of the determinant factor for glycation. This study aims to prove the protein molecular mass also one of the factors responsible for glycation. In this study, in vitro glycation of several model proteins like IgG, HSA, BSA, apomyoglobin, papain and insulin, were performed by MALDI-TOF-MS. The results support stated hypothesis that the molecular mass of protein, is also responsible in determining the glycation reaction.
MATERIALS AND METHODS
This study was conducted at Division of Organic Chemistry and Center for Material Characterization, National Chemical Laboratory, Pune, India.
Chemicals
Bovine serum albumin, human serum albumin, apomyoglobin, papain and glucose were procured from Sigma Chemicals Ltd., immunoglobin G (mass standards) from Applied Biosystems, and insulin (Biocon Ltd.) was purchased from local Pharmaceuticals.
In vitro Glycation Experiments
HSA (0.15 mM), BSA (15 mM), papain (42 mM), insulin (34 mM) apomyoglobin (0.05 mM) and IgG (0.006 mM) were used for in vitro glycation assay. Samples were prepared in double distilled water. The in vitro glycation was carried out by incubating equal volume (50 μL) of protein with 0.5 M glucose solution dissolved in 0.2 M phosphate buffer, at 37°C for either 3, 6 or 10 days.
Sample Preparation
Five microliter of the assay mixture was mixed with 45 μL of freshly prepared sinapinic acid (30% acetonitrile, 0.1% trifluoroacetic acid) and loaded onto the stainless steel MALDI plate by dried-droplet method and incubated for 10 min at 37°C.
MALDI Analysis
The mass spectral analysis was done on Voyager-De-STR (Applied Biosystems) MALDI-TOF. A 337 nm pulsed nitrogen laser was used for desorption and ionization. An Aquiris 2 GHz dizitiger board was used for all experiments. MALDI analysis of insulin, apomyoglobin, papain, HSA, BSA and IgG were done using different instrumental settings. For insulin analysis, spectra were acquired in the range of 2500 to 7500 Da, on a positive reflector mode with the following settings: an accelerating voltage 25 kV, grid voltage 68.5% of accelerating voltage, delayed ion extraction time of 350 nsec, low mass ion gate was set to 500 Da. For apomyoglobin analysis, spectra were acquired in the range of 1000 to 25000 Da, on a positive linear mode with the following settings: an accelerating voltage 25 kV, grid voltage 93.2% of accelerating voltage, delayed ion extraction time of 1100 sec, low mass ion gate was set to 1000 Da. For HSA, BSA, papain and IgG analysis, spectra were acquired in the mass range of 10 to 200 kDa, on a positive linear mode with the following settings: an accelerating voltage 25 kV, grid voltage 92% of accelerating voltage, delayed ion extraction time of 1500 nsec, low mass ion gate was set to 4500 Da. For all spectral acquisition, the laser power was set just above the ion generation threshold to obtain peaks with highest possible signal to noise ratio. All spectra were acquired with 50 shots in 3 replications. The spectra were processed for advanced base line correction, noise removal and mass calibration.
RESULTS AND DISCUSSION
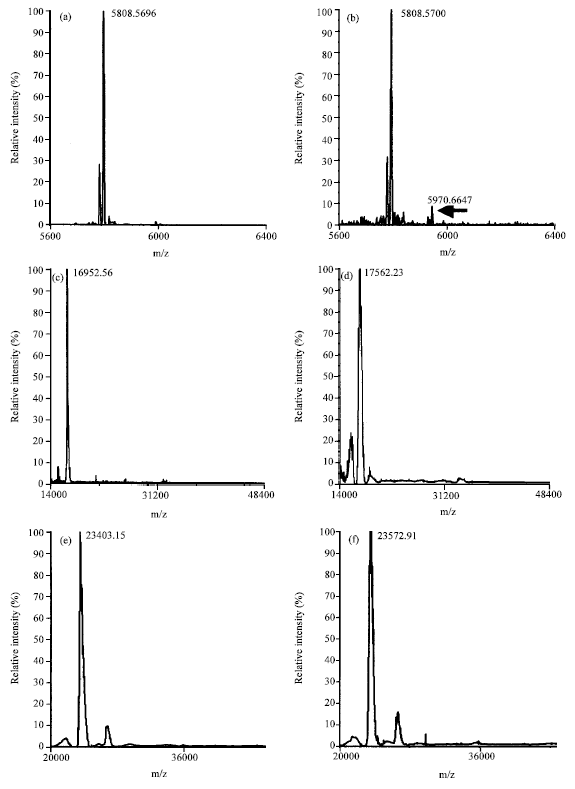
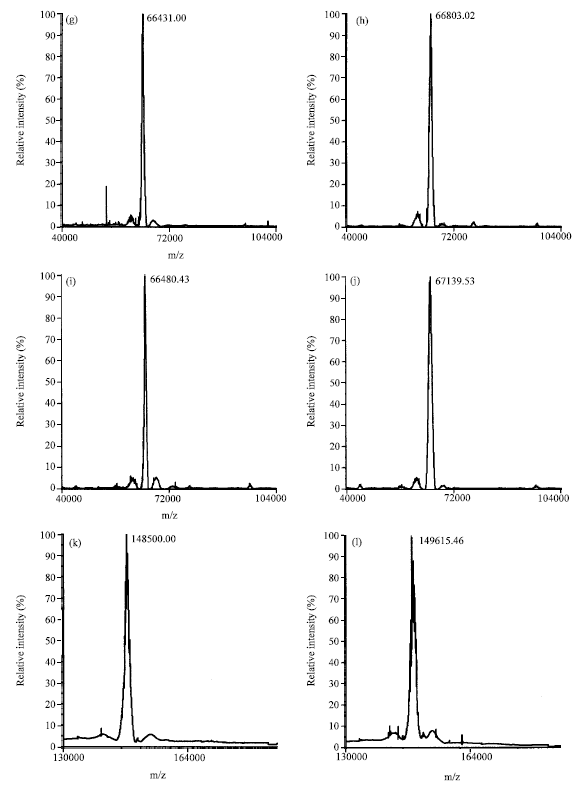
To understand the role of molecular mass in determining of glycation, several model proteins of varying mass like insulin, apomyoglobin, papain, HSA, BSA, IgG were used for in vitro glycation studies. These proteins were incubated with 0.5 M glucose for a period of one week. The glycation reaction was analyzed using MALDI-TOF-MS by monitoring the increase in mass of the proteins. Figure 1a-l show the mass of control and glycated proteins by MALDI-TOF-MS analysis. All the proteins show increase in mass (ΔM) upon glycation. The extent of glycation was lower for low molecular mass proteins such as insulin, apomyoglobin and papain as reflected by lesser increase in mass compared to the high molecular mass proteins such as BSA, HSA and IgG. The increase in mass was analyzed for different proteins during glycation reaction (Fig. 2).
 | |
 | |
| Fig. 1: | MALDI-TOF-MS analysis of control and glycated masses of different proteins. Spectrum labeled as a, c, e, g, I and k show mass of control insulin, apomyoglobin, papain, BSA, HSA, IgG, respectively and spectrum labeled b, d, f, h, j and l show mass of glycated insulin, apomyoglobin, papain, BSA, HSA and IgG, respectively |
 | |
| Fig. 2: | Increase in mass of different proteins on different days of glycation. Protein masses were checked on 3rd, 6th and 10th day of glycation reaction using MALDI-TOF-MS |
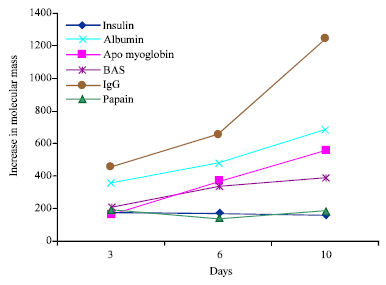
The ΔM did not change for low molecular mass proteins such as insulin and pepsin on different days of glycation. Whereas ΔM increased with increase in duration of glycation reaction for high molecular mass proteins such as BSA, HSA and IgG. The relationship between ΔM and the mass of proteins on different days of glycation were positively correlated with the increase in mass on all different days of glycation 8 (day 3, R2 = 0.846; day 6, R2 = 0.772 and day 10, R2 = 0.799) (Fig. 3). These results show the importance of molecular mass in determining the protein glycation reaction.
Glycation occurs at ε-amino group such as lysine, hydroxylysine or arginine (Zhao et al., 1997). Larger proteins have naturally more number of lysine and arginine residues; therefore, it is obvious to observe increase in the extent of glycation in larger proteins compared to smaller proteins. In a recent study, it was observed that the Human Serum Albumin (HSA), a large molecular mass protein with 59 lysine residues, is heavily glycated in vivo as evidenced by the 31 glycation sites identified from 38 unique glycated peptides (Zhang et al., 2008). One of the major reasons of HSA being highly glycated is simply by virtue of number of lysine residues. Another possible reason could be the nature of amino acid present near the lysine residue. It was demonstrated earlier, that the vicinal amino acids strongly affects lysine reactivity towards the different carbohydrates. The presence of hydrophobic residues such as isoleucine, leucine, phenylalanine strongly increases lysine reactivity (Menella et al., 2006).
Theoretically any protein can undergo glycation, as they may have at least one ε-amino acid. Zhang et al. (2008) for the first time identified 76 non-enzymatically glycated proteins from the plasma of diabetic patients. Identification of lesser number of glycated proteins was mainly attributed to the presence of abundant proteins like albumin and IgG. Furthermore, all the identified proteins were present in the moderate-to low-abundance range in the plasma proteome. The discovery of very low abundant glycated proteins was like a needle in a haystack, which suggests that identifying low abundant glycated proteins is a technically challenging task. However, the earlier study suggests that the abundant protein like albumin inhibits/regulates the glycation of less abundant proteins by mere competition (Bhonsle et al., 2008). This study suggests that identifying low abundant glycated proteins is not just a technical challenge but in fact high abundant proteins may inhibit the glycation of less abundant proteins in vivo. Therefore, the conclusion is that the molecular mass of protein as well as protein abundance are also important factors that determine glycation reaction in addition to the protein structure and its turnover.
 | |
| Fig. 3: | Relationship between ΔM and molecular mass of proteins. Where ΔM is the increase in mass of proteins. (a) Correlation between molecular mass of proteins and ΔM on day 3 of glycation, (b) correlation between molecular mass of proteins and ΔM on day 6 of glycation and © correlation between molecular mass of proteins and ΔM on day 10 of glycation |
ACKNOWLEDGMENT
Authors are thankful to Dr. Sivaram, Director NCL for his support and encouragement. This study was carried out as apart of a project entitled Proteomics of diabetic complications sponsored by Department of Biotechnology, India.
REFERENCES
- Bhonsle, H.S., S.K. Singh, G. Srivastava, R. Boppana and M.J. Kulkarni, 2008. Albumin competitively inhibits glycation of less abundant proteins. Protein Peptide Lett., 15: 663-667.
PubMed - Brownlee, M., 2001. Biochemistry and molecular cell biology of diabetic complications. Nature, 414: 813-820.
CrossRefPubMedDirect Link - Hunter, S.J., A.C. Boyd, F.P.M. O` Harte, A.M. McKillop and I.M. Wiggam et al., 2003. Demonstration of glycated insulin in human diabetic plasma and decreased biological activity assessed by euglycemic hyperinsulinemic clamp technique in humans. Diabetes, 52: 492-498.
PubMed - Kueper, T., T. Grune, S. Prahl, H. Lenz and V. Welge et al., 2007. Vimentin is the specific target in skin glycation. J. Biol. Chem., 282: 23427-23436.
CrossRef - Lapolla, A., D. Fedele, R. Seraglia and P. Traldi, 2006. The role of mass spectrometry in the study of non-enzymatic protein glycation in diabetes. Mass Spectrometry Rev., 25: 775-797.
CrossRef - Menella, C., M. Visciano, A. Napolitano, M.D.C. Castillo and V. Fogliano, 2006. Glycation of lysine-containing dipeptides. J. Peptide Sci., 12: 291-296.
CrossRef - Poggioli, S., H. Bakala and B. Friguet, 2002. Age-related increase of protein glycation in peripheral blood lymphocytes is restricted to preferential target proteins. Exp. Gerontol., 37: 1207-1215.
CrossRef - Thornalley, P.J., 2003. Glyoxalase I – structure, function and a critical role in the enzymatic defence against glycation. Biochem. Soc. Trans., 31: 1343-1348.
PubMed - Zhang, Q., N. Tang, A.A. Schepmoes, L.S. Phillips, R.D. Smith and T.O. Metz, 2008. Proteomic profiling of non-enzymatically glycated proteins in human plasma and erythrocyte membranes. J. Proteome Res., 7: 2025-2032.
CrossRef