
Maha Z. Rizk
Department of Medicinal Chemistry, National Research Centre, Dokki, Cairo, Egypt
Thanaa A. Fayed
Faculty of Science, Mansoura University, Mansoura, Egypt
Hohey D. Badawy
Faculty of Science, Mansoura University, Mansoura, Egypt
Nagy S. El-Rigal
Department of Medicina1 Chemistry, National Research Centre, Dokki, Cairo, Egypt
Trends in Medical Research
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 66-74







ABSTRACT
The levels of lipid peroxides, glutathione, vitamin C, vitamin E catalase and superoxide dismutase enzymes were measured in the livers of Schistosoma mansoni infected mice (CD strain weighing 20-25 g) at different durations post infection (1, 2, 4, 6 and 8 weeks). Moreover, the liver weight, body weight, liver to body weight and total protein were studied in the same animal groups. Control non-infected groups were run simultaneously with each infected mice group. The data obtained showed that lipid peroxides were elevated throughout the different durations of infection while glutathione decreased with infection. On the other hand, both vitamins C and E showed a reduction in the livers of mice during the different durations of infection. The activity of catalase showed an insignificant change after one and two weeks and a high significant decrease in the livers of four, six and eight weeks S. mansoni infected mice, while, superoxide dismutase significantly decreased one and two weeks post infection with a significant elevation four, six and eight weeks post infection. Moreover, a significant reduction was observed in body weight after four, six and eight weeks of infection accompanied with an elevation in liver weight only after six and eight weeks. Consequently, liver weight/body weight showed an elevation after four, six and eight weeks of infection. Finally, the protein content was significantly lower at one, six and eight weeks post infection with S. mansoni. It could thus be concluded that host-parasite association results in production of free radicals as a result of an oxidative stress where the parasites struggle to overcome the immune response of the host and changes in host liver antioxidants occur as a means to scavenge these radicals.
PDF Abstract XML References
How to cite this article
Maha Z. Rizk, Thanaa A. Fayed, Hohey D. Badawy and Nagy S. El-Rigal, 2006. Effect of Different Durations of Schistosoma mansoni Infection on the Levels of Some Antioxidants in Mice. Trends in Medical Research, 1: 66-74.
URL: https://scialert.net/abstract/?doi=tmr.2006.66.74
URL: https://scialert.net/abstract/?doi=tmr.2006.66.74
INTRODUCTION
Schistosomiasis ranks next to malaria regarding all the endemic parasitic diseases worldwide particularly in Egypt (El-Baz et al., 2003). In parallel with massive research efforts in schistosomiasis over the past 30 years, persistent efforts have been made to understand the basis for infection (Van Nassauw et al., 2001). The tropical parasite Schistosoma mansoni causes granulomatous inflammation after its eggs lodge in the hepatic portal capillaries. Previous studies indicate that the host's response involves the production of reactive oxygen species (Connors et al., 1995; El-Sokkary et al., 2002). Sayed and Williams (2004) reported that schistosomes use alternative electron donors and their variable resistance to overoxidation may reflect their presence in different cellular sites and emphasizes the significant differences in overall redox balance mechanisms between the parasite and the mammalian host.
The consequences of free radical generation in S. mansoni is still unknown (Facundo et al., 2004). In some reports concerning Schistosoma mansoni, Sheweita et al. (2003) mentioned that high levels of free radicals provide new evidence for organ damage since glutathione is decreased and lipid peroxides increased in different organs of hamsters infected with S. hematobium. In this manner. S. mansoni infection alters the hepatic levels of glutathione and activities of glutathione metabolizing enzymes and alterations may affect the capacity of the liver to detoxify or neutralize the toxic effect of endogenous and exogenous compounds.
From the previous information, it was of interest in the present study to investigate the levels of glutathione (GSH) vitamin E (VE), vitamin C (VC), superoxide dismutase (SOD), catalase and lipid peroxides, in infected mice groups at different stages of infection with Schistosoma mansoni in order to trace the changes that occur in these substances throughout the infection period. The objective of the study is to investigate the situation under the disease condition throughout the stages of parasite development, whether shortly after invasion or after egg deposition, since, substances expressed during the life cycle of the parasite such as free radicals may provide useful approaches as stage specific markers for development of drugs that may counteract infection attack. This may be achieved in the future by searching for a nutrient that can protect mitochondrial enzymes and/or stimulate enzymatic activity by elevating levels of substrates and cofactors; that can enhance antioxidant defences; that can scavenge free radicals and prevent oxidant production in mitochondria and finally that can repair mitochondrial membrane.
MATERIALS AND METHODS
All chemicals used were of high analytical grade, products of Sigma (USA), Merck and Reidel (Germany), BDH (England) and Fluka (Switzerland) Chemical Co. Praziquantel used is a product of Egyptian International Pharmaceutical Industries Company (E.I.P.I.Co.).
The animals used were healthy male Swiss albino mice of CD strain, obtained from Theodor Bilharz Institute, ranging in weight from 20-25 g. They received fresh, deionized water and standard diet (El-Tkamol-Co.) ad libitum. Mice were divided into two main groups each divided into five subgroups.
The first main infected group was sacrificed one, two, four, six and eight weeks post infection with 100 cercariae of Egyptian Schistosoma mansoni strain.
The second main group served as control and was run simultaneously with each duration of the infected subgroups. The number of animals ranged from 6-10 mouse in each subdivision.
To obtain cercariae, 10-20 shedding Biomphalaria alexandrina snails were used to ensure bisexual infection and placed in a beaker containing 200 mL dechlorinated water. Then, the snails were exposed to sunlight at 8-9 am. Each mouse was infected subcutaneously by 100 cercariae.
Experimental Procedures
The liver was removed from the mouse, plotted with filter paper and weighted. The liver was homogenized in bidistilled water (5, 10 and 20% w/v) according to the parameter measured, using potter-Elvehjem homogenizer with Teflon pestle.
Estimation of Protein
Protein was estimated by the method of Bradford (1976). Bradford solution was added to of 5% homogenate and the developed blue colour was measured after 5 min at 595 nm in spectrometer (Novaspec, LKB, Sweden) against a blank containing water instead of the homogenate. The amount of protein was calculated from a standard curve using serial concentration of bovine serum albumin (1-10 ug).
Estimation of Lipid Peroxides
Lipid peroxides were estimated by thiobarbituric acid reaction as described by Ohkawa et al. (1979) using saturated thiobarbituric acid (A) and trichloroacetic acid (20%) (B) solutions. The working solution was prepared by mixing one volume of solution A with 3 volumes of solution B. 20% tissue homogenate was added to the working solution and mixed. The tubes were boiled, cooled and centrifuged at 3.000 rpm for 10 min. The developed colour was read against the blank at 532 nm in spectrometer and calculated as μmole malonaldehyde per gram tissue.
Estimation of Glutathione
Glutathione was estimated by the method of Moron et al. (1979) using sodium phosphate buffer and dithiobisnitrobenzoic acid (DTNB). The developed colour was read against blank at 412 nm within 5 min in spectrometer. The amount of glutathione was calculated as mg glutathione per gram tissue used from a standard curve plotted for serial concentrations of glutathione (5-100 μg).
Estimation of Vitamin C
Vitamin C was estimated by the method of Jagota and Dani (1982) using Folin-ciocalteu reagent (2.0 M concentration diluted 3-fold with double-distilled water) and TCA. The developed blue colour was read at 760 nm after 10 min in spectrometer against a blank. The amount of ascorbic acid was calculated from a standard curve of vitamin C using 5-70 μg serial concentrations of the vitamin.
Estimation of Vitamin E
Vitamin E was estimated by the method of Baker and Frank (1968) using 7.7 m M α, α-Dipyridyl and 7.4 mM ferric chloride solution. The absorbance of colour was read for the test and standard against the blank at 460 nm.
Estimation of Superoxide Dismutase Activity
The activity of superoxide dismutase was estimated by the method of Nishikimi et al. (1972) using nitroblue tetrazolium, NADH and sodium pyrophosphate buffer. Increase in absorbance with time during the reaction was read in spectrometer at 560 nm.
Estimation of Catalase Enzyme
Catalase activity was assayed according to Lubinsky and Bewley (1979). The reaction for assaying catalase activity was initiated by adding 20 μL 5% liver homogenate of concentration to 2.53 mL of the reaction mixture (sodium potassium phosphate buffer containing H2O2. The disappearance of hydrogen peroxide was monitored by following the decease in absorbance at 230 nm by spectrometer using molar extinction coefficient for hydrogen peroxide of 62.4 (Nelson and Kiesow, 1972).
Statistical Analyses
The analyses included the calculation of the mean value, standard deviation, standard error and t-values at level p≤0.05 according to the method of Ronald et al. (1983). These determinations were calculated for both control and treated animals.
RESULTS
From the result of Table 1 it is obvious that infected mice showed a highly significant increase in the levels of lipid peroxides with percentages 31.29, 40.0, 89.0 and 66.78% after one, two, six and eight weeks of infection, respectively, while a significant elevation was detected in the lipid peroxides after four weeks of infection with percentage change 8.69%. On the oher hand Schistosoma mansoni infection showed a highly significant decrease in liver glutathione of mice, with percentages 44.0, 19.88, 24.60, 32.46 and 40.0% after one, two, four, six and eight weeks of infection, respectively.
| Table 1: | Estimation of lipid peroxides and glutathione in the liver of mice during different durations of Schistosoma mansoni infection |
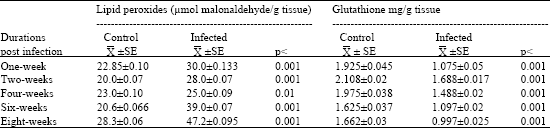 | |
| Data are means (±SE) of lipid peroxides and glutathione determinations. Significant change as compared to control (p<0.001 : highly significant and p<0.01: significant) | |
| Table 2: | Determination of vitamin C and vitamin E in the liver of mice during different durations of Schistosoma mansoni infection |
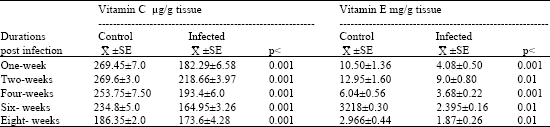 | |
| Data are means (±SE) of vitamin C and vitamin E determinations. Significant change as compared to control (p<0.001: highly significant and p<0.01: significant) | |
It is obvious that liver vitamin C of infected mice showed a highly significant decrease after one, two, four, six and eight weeks post infection, with percentages 32.34, 18.90, 23.78, 29.70 and 6.80%, respectively Table 2. Also, vitamin E showed a highly significant decrease after one and four weeks of infection with percentages 61.0 and 39.0%, respectively and a significant reduction after two, six and eight weeks of infection with percentages 30.50, 25.57 and 36.95%, respectively as compared with clean control mice.
Table 3 shows that, an insignificant decrease of the catalase activity was detected after one and two weeks of infection, while a highly significant decrease of the liver enzyme activity was detected after four, six and eight weeks of the infection with percentages 18.42, 19.93 and 25.0%, respectively as compared with normal control mice. On the other hand superoxide dismutase enzyme showed a significant decrease in the liver of infected mice with percentages 10.67 and 5.57%, respectively after one and two weeks of the infection with S. mansoni. Also, the enzyme activity showed a highly significant increase after four, six and eight weeks of the infection giving percentages 11.85, 13.70 and 17.13%, respectively as compared with normal control mice.
It is obvious (Table 4) that an insignificant change is observed in the body weight after one and two weeks of infection, while a significant decrease was recorded with percentage 7.67, 4.66 and 15.80% after four six and eight weeks of infection with S. mansoni, respectively. Liver weight showed an insignificant change after one and four weeks of infection and a significant increase after two weeks of infection with percentage 4.0% while, a highly significant increase is recorded in liver weight after six and eight weeks of infection with percentages 25.60 and 22.0%, respectively. Liver weight/ body weight ratio showed an insignificant change after one and two weeks, while a slightly significant increase with percentage 4.90% was shown after four weeks of infection and a highly significant increase after six and eight weeks of infection with percentage 32.0 and 38.39% as compared with normal control .
Table 5 shows the level of total proteins in control and infected mice during different periods of infection with S. mansoni. It is observed that the level of total protein showed a significant decrease after one week of infection with percentage 4.72% and an insignificant change after two weeks of infection.
| Table 3: | Estimation of catalase and superoxide dismutase enzyme activities in the liver of mice during different durations of Schistosoma mansoni infection |
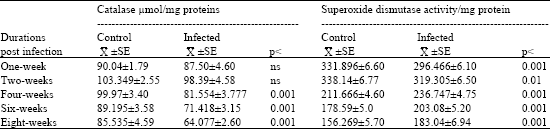 | |
| Data are means (±SE) of catalase and superoxide dismutase determinations. Significant change as compared to control (p<0.001: highly significant, p<0.01: significant and ns: insignificant). Data are means (±SE) of catalase and superoxide dismutase determinations | |
| Table 4: | Body weight, liver weight and liver weight/body weight ratio of control and infected mice during different durations post infection with S. mansoni |
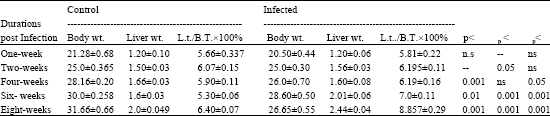 | |
| Data are means (±SE) of body wt., liver wt. and liver wt./body wt. ratio. Significant change as compared to control (p<0.001: highly significant, p<0.01: significant, 0.05: slightly significant and ns: insignificant) | |
| Table 5: | Determination of total protein in the liver of control and infected mice during different durations post infection with Schistosoma mansoni |
 | |
| Data are means (±SE) of total protein level. Significant change as compared to control (p<0.001: highly significant, p<0.01: significant and ns : insignificant) Total protein is expressed in mg protein/g tissue used | |
After four weeks of infection the total protein showed a highly significant increase with percentage 25.50%, while after six and eight weeks of infection the total protein showed a highly significant decrease with percentage 11.90 and 16.0%, respectively.
DISCUSSION
It was previously reported that the understanding of how schistosomes interact with their hosts, which is a highly complex strategy, may be one mechanism by which a suitable vaccine or chemotherapeutic drug could be developed (El-Ansary and Daihan, 2005). Previous authors reported that granuloma macrophages isolated from hepatic, intestinal and pulmonary lesions were found to release significant amounts of O2 and H2O2 radicals (Shaheen et al.,1994). The oxidative processes which occur upon contact with S. mansoni eggs trapped in the liver seem to proceed uncontrolled, since the enzymatic activities involved in O2 and H2O2 detoxification decrease drastically. Such events may be, at least in part, responsible for the pathology associated with schistosomiasis (Gharib et al., 1999). The infection with S.mansoni not only triggers the production of reactive oxygen species, but it also leads to the alteration of the antioxidant defence mechanism (Pascal et al., 2000).
The data obtained in the present study show that lipid peroxides were elevated by S. mansoni throughout the different durations of infection. It should be pointed out that fibrosis associated with the disease is stimulated by reactive end products of lipid peroxidation (Bedossa et al., 1994; Soliman et al., 2001). In addition, there is a positive correlation between collagen deposition during schistosomal infections and production of malonaldehyde by hepatic cells (Parola et al., 1996).
Results on glutathione content in infected mice livers revealed a highly significant reduction resulting from an oxidative stress due to schistosomiasis. Such depletion may be caused by increased cytotoxicity with H2O2 which leads to inhibition in glutathione reductase, the latter responsible for keeping glutathione in its reduced state (Harlan et al., 1984). An interesting finding which coincides with the present data was shown by Yegen et al. (1990) that reduction of cellular GSH is accompanied by increased lipid peroxidation.
The present work was also extended to investigate the level of vitamin C in the different Schistosoma infected mice groups. It was found that this vitamin was significantly reduced to scavenge the free radicals produced by the parasite since peroxyl radicals are effectively trapped by ascorbate (Frei et al., 1989).
Data obtained for vitamin E in infected mice livers revealed a significant reduction in the vitamin since the latter is required to protect biological membranes against oxidative degeneration. The administration of vitamin E in the diet shows a strong antioxidant activity against lipid peroxidation and also it protects hepatocytes against toxic injury (Nardini et al., 1993; Sokol et al., 1998). Reduction in the vitamin content due to oxidative stress was previously reported by Warren and Reed (1991).
In the present research, the activity of superoxide dismutase greatly declined one week post infection while an increase in the enzyme activity started from the 4th week till the end of the infection period. The decrease in SOD may result from production of H2O2 during oxidative metabolism as indicated by Pinteaux et al. (1996). Moreover the increase in enzyme activity may be an adaptive response to conditions of increased peroxidative stress in the liver. Previously, Shaheen et al. (1994) reported an increase in hepatic SOD in mice infected with S. mansoni. An interesting finding was reported by Sanz et al. (1996) who showed that cell defence mechanisms against oxidative toxicity increase in liver to suppress oxidative imbalance, thus SOD increases and GSH decreases.
Furthermore, the present data reveals a highly significant and progressive reduction in catalase activity which started four weeks post S. mansoni infection. In agreement with this, Gharib et al. (1999) showed that peroxide dismutation yields H2O2 which is detoxified by catalase resulting in decrease in its activity. In a recent study, Hanna et al. (2005) added that eosinophil peroxidase and its substrate H2O2 are released by inflammatory cells in the immediate vicinity of parasite eggs. Also in a more recent study El-Rigal et al. (2006) reported the inhibitory effect of S.mansoni on glutathione, vitamin C, vitamin E and catalase in infected mice while lipid peroxides increased.
With respect to body and liver weights, it was found that a significant reduction in body weight was recorded after four weeks of infection. These changes may occur due to the presence of the developing parasite worms and the initiation of egg deposition and also due to several metabolites released by the parasite which affect the host hepatic tissue (El-Marzouki and Amin, 1997). In a good connection to the present results, Ahmed and Gad (1995); Magalhaes et al. (1995); Fiore et al. (1996) and Soliman et al. (2001) found a reduction in body weight , an increase in liver weight and an increase in liver weight/body weight from the sixth week post-infection with S. mansoni, with initiation of schistosomal egg deposition.
On the other hand, data on total proteins recorded an increase after four weeks of infection then a significant decline after six and eight weeks. In hepatic fibrosis as a result of bilharzial infection, protein anabolism decreases while protein catabolism increases. Impairment in protein synthesis was previously supported by Coutinho et al. (2002) that malabsorption may be a contributing factor in decrease of protein synthesis through a defect in absorption of amino acids.
In conclusion, the objective of the present research was to study some aspects concerning host-parasite interaction. Previous studies have dealt with the production of free radicals in schistosomiasis, but only after egg deposition in liver and granuloma formation. No reports were cited concerning the trace of these elaborated substances throughout the developmental stages of the parasite in order to provide knowledge on the host-parasite complex interaction, to elucidate stage-specific markers and to develop a suitable target drug that can counteract the parasite at a certain stage of infection. The free radicals are mostly released by inflammatory cells in the vicinity of parasite eggs and the collagen fibers built around eggs create a barrier preventing released compounds from diffusing freely into surrounding areas. This allows the parasitized liver to cope with the threat posed by parasite eggs. This study may help to elucidate stage-specific bioactive markers which may help to develop novel intervention strategies that can modulate the host immune system.
REFERENCES
- Ahmed, S.M. and M.Z. Gad, 1995. Effect of Schistosoma infection and its treatment on some key enzymes of glucose metabolism in mice livers. Arzneimittelforschung, 45: 1324-1328.
Direct Link - Bedossa, P., K. Houglum, C. Trautwein, A. Holstege and M. Chojkier, 1994. Stimulation of collagen α-1(1) gene expression is associated with lipid peroxidation in hepatocellular injury: A link to tissue fibrosis. Hepatology, 19: 1262-1271.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Connors, V.A., I. Deburon and W.O. Granath, 1995. Schistosoma mansoni.: Interleukin-1 increases phagocytosis and superoxide dismutase by hemocytes and decreases output of cercariae in schistosome-susceptible Biomphalaria glabrata. Exp. Parasitol., 80: 139-148.
Direct Link - El-Ansary, A. and S.A. Daihan, 2005. Stage-specifically expressed schistosome proteins as potential chemotherapeutic targets. Med. Sci. Monit., 11: 94-103.
Direct Link - El-Baz, M.A., T.A. Morsy, M.M. E. Bandary and S.M. Motaweae, 2003. Clinical and parasitological studies on the efficacy of mirazid in treatment of Schistosomiasis haematobium in Tatoon, EISA center, El-Fayoum Governorate. J. Egypt. Soc. Parasitol., 33: 761-776.
Direct Link - El-Rigal, N.S., S.A. Aly, M. Rizk and A. Said, 2006. Effect of Ailanthus altissima and Ziziphus spina christi extracts on some hepatic marker enzymes and antioxidants in Schistosoma mansoni infected mice. Pol. J. Food Nutr. Sci., 15: 199-206.
Direct Link - El-Sokkary, G.H., H.M. Omar, A.T. Hassanein, S. Cuzzocrea and R.J. Reiter, 2002. Melatonin reduces oxidative damage and increases survival of mice infected with S. mansoni. Free Rad. Biol. Med., 32: 319-332.
PubMedDirect Link - Facundo, H.T.F., C.T. Brandt, J.S. Owen and V.L.M. Lima, 2004. Elevated levels of erythrocyte-conjugated dienes indicate increased lipid peroxidation in S. mansoni patients. Braz. J. Med. Biol. Res., 37: 957-962.
CrossRefDirect Link - Fiore, M., R. Moroni, E. Alleva and L. Alae, 1996. Schistosoma mansoni: Influence of infection on mouse behaviour. Exp. Parasitol., 83: 46-54.
CrossRefPubMedDirect Link - Frei, B., L. England and B.N. Ames, 1989. Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Natl. Acad. Sci. USA., 86: 6377-6381.
CrossRefPubMedDirect Link - Hanna, S., B. Gharib, H. Lepidi, J.C. Montet, H. Dumon and M.D. Reggi, 2005. Experimental schistosomiasis, protective aspects of granulomatous reaction in the mouse liver. Parasitol. Res., 96: 6-11.
CrossRefDirect Link - Harlan, J.M., J.D. Levine, K.S. Callahan, B.R. Schwartz and L.A. Harker, 1984. Glutathione redox cycle protects cultured endothelial cells against lysis by extracellularly generated hydrogen peroxide. J. Clin. Invest., 73: 706-713.
CrossRefDirect Link - Moron, M.S., J.W. Depierre and B. Mannervik, 1979. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Gen. Subj., 582: 67-78.
CrossRefPubMedDirect Link - Nardini, M., C. Scaccini, M.D. Aquino, P.C. Benedetti, G. Tomassi and M.D. Felice, 1993. Lipid peroxidation in liver microsomes of rats fed soybean, olive and coconut oil. J. Nutr. Biochem., 4: 39-44.
Direct Link - Nelson, D.P. and L.A. Kiesow, 1972. Enthalpy of decomposition of hydrogen peroxide by catalase at 25 C (with molar extinction coefficientof H2O2solutions in the UV). Anal. Biochem., 49: 474-478.
CrossRef - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Parola, M., G. Leonarduzzi, G. Robino, E. Albano, G. Poli and M.U. Dianzani, 1996. On the role of lipid peroxidation in the pathogenesis of liver damage induced by long-standing cholestasis. Free Rad. Biol. Med., 20: 351-359.
PubMedDirect Link - Pascal, M., O.M.A. Allah, N.E. Elwali, A. Mergani and M.A. Quarshi et al., 2000. Hyaluronate levels and markers of oxidative stress in the serum of Sudanese subjects at risk of infection with Schistosoma mansoni. Trans. R. Soc. Trop. Med. Hyg., 94: 66-70.
Direct Link - Sanz, N., F.C. Diez and M. Cascales, 1996. Variations of hepatic antioxidant systems and DNA ploidy in rats aged 2 to 8 months. Biochem. Biophys. Acta, 1315: 123-130.
PubMedDirect Link - Sayed, A.A. and D.L. Williams, 2004. Biochemical characterization of 2-Cys peroredoxins from S. mansoni. J. Biol. Chem., 279: 26159-26166.
Direct Link - Shaheen, A.A., A.A. Abd-El-Fattah and F.A. Ebeid, 1994. Effects of praziquantel treatment on lipid peroxide levels and superoxide dismutase activity in tissues of healthy and Schistosoma mansoni infected mice. Arzneimittelforschung, 44: 94-96.
PubMedDirect Link - Sokol, R.J., J.M. Mckim. M.C. Goff, S.Z. Ruyle and M.W. Devereux et al., 1998. Vitamin E reduces oxidant injury to mitochondria and hepatotoxicity of taurochenodeoxycholic acid in rat. Gastroenterol., 114: 164-174.
Direct Link - Soliman, K.M., A.K. El-Ansary and A.M. Mohamed, 2000. Effect of carnosine administration on certain metabolic parameters in bilharzial infected hamsters. J. Egypt. Soc. Parasitol., 30: 455-468.
PubMed - Van Nassauw, L., J. Bogers, E.V. Marck and J.P. Timmermans, 2001. Role of reactive nitrogen species in neuronalcell damage during intestinal schistosomiasis. Cell Tissue Res., 303: 329-336.
CrossRefDirect Link - Warren, D.L. and D.J. Reed, 1991. Modificationof hepatic vitaminE stores in vivo.I.Alterations in plasma and liver vitaminE content by methyl ethyl ketone peroxide. Arch. Biochem. Biophys., 285: 45-52.
Direct Link - Yegen, B., A. Dedeoglu, I. Aykac, S. Oktay and A.S. Yalcin, 1990. Effect of cold-restraint stress on glutathione and lipid peroxide levels in the liver and glandular stomach of rats. Pharm. Res., 221: 45-48.
PubMedDirect Link - Jagota, S.K. and H.M. Dani, 1982. A new colorimetric technique for the estimation of vitamin C using folin phenol reagent. Anal. Biochem., 127: 178-182.
CrossRefDirect Link - Nishikimi, M., N.A. Rao and K. Yagi, 1972. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun., 46: 849-854.
CrossRefPubMedDirect Link