
Usman S.F. Tambunan
Department of Chemistry, Faculty of Mathematics and Natural Sciences, University of Indonesia, Depok Campus, Depok 16424, Indonesia
Oksya Hikmawan
Department of Chemistry, Faculty of Mathematics and Natural Sciences, University of Indonesia, Depok Campus, Depok 16424, Indonesia
Theofilus A. Tockary
Department of Chemistry, Faculty of Mathematics and Natural Sciences, University of Indonesia, Depok Campus, Depok 16424, Indonesia
Trends in Bioinformatics
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 18-24







ABSTRACT
The aim of this study was to analyse the mutation possibility of influenza A (H5N1) virus of Banten Province strain. The H5N1 amino acid sequences of haemagglutinin (HA) and neuraminidase (NA) were analysed by multiple alignment method using BLAST tool, ClustalW and Bioedit 7.0.1. programs. The results of analysis were followed up by mutation analysis using Amino Track, secondary structure, post translation modification analysis, phylogenetic tree and homology modelling. From the analysis, five point mutations were revealed, namely change of haemagglutinin at position 185, 272 and 309 and neuraminidase at position 244 and 252. Conserved region prediction were determined to be at position 1-552 of HA and at position 1-39, 41-251 and 253-449 of NA. Secondary structure prediction showed a change in HA structure at position 309, namely coil into helix transformation and in NA structure at position 244, namely coil into extended strand transformation. Change of hydrophobicity happened only to HA at position 185 and 272. Mutation did not have any influence on post translation modification. Phylogenetic tree analysis and homology modelling also did not reveal the creation of a new strain.
PDF Abstract XML References Citation
How to cite this article
Usman S.F. Tambunan, Oksya Hikmawan and Theofilus A. Tockary, 2008. In silico Mutation Study of Haemagglutinin and Neuraminidase on Banten Province Strain Influenza A H5N1 Virus. Trends in Bioinformatics, 1: 18-24.
DOI: 10.3923/tb.2008.18.24
URL: https://scialert.net/abstract/?doi=tb.2008.18.24
DOI: 10.3923/tb.2008.18.24
URL: https://scialert.net/abstract/?doi=tb.2008.18.24
INTRODUCTION
Avian influenza is caused by viral infection of Orthomyxoviridae family in the influenza virus A genus. This virus was originally recognised only as the causative agent of fowl plague in 1955. However, in the last few years, the occurrence of highly pathogenic avian influenza A (H5N1) virus also began to threaten human safety after fowl to human transfection cases (Kawaoka et al., 1989) and was reported to increase (Horimoto and Kawaoka, 2001). This problem arises due to the virus tendency to mutate and recombine with genetical material of other influenza virus (Anwar et al., 2006; Ungchusak et al., 2005). Very limited human-to-human transmission of the H5N1 strain was documented in healthcare workers and family members with contact (Katz Jacqueline et al., 1999; Bridges et al., 2000). For the same reason, some experts are now even fearing the worst possibility of another new strain that might be capable of human to human transfection.
In a recent case of avian influenza outbreak in the Indonesian Province of Banten, several members of a certain family were infected by a presumably new strain of this virus. Some suspected who might actually be the first case of human to human transfection as some of the family members could not recall having direct contact with fowls. This present study is an effort to settle this claim by conducting series of in silico examination on the Banten Province strain`s haemagglutinin and neuraminidase amino acid sequences. It is expected that may reveal the specific mutation sites of the strain and altering of antigenicity, specificity and pathogenicity of the virus (Glaser et al., 2005). As is the case of the most viruses, these three altering parameters are strong indications of a new strain. The reasons for choosing haemagglutinin and neuraminidase for this study are (1) both are well-known as antigenic macromolecules, (2) haemagglutinin`s participation in binding to the host cell is strongly connected to specificity and (3) neuraminidase is a virulence factor.
Haemagglutinin and neuraminidase amino acid sequences of the Banten Province strain (A/Indonesia/CDC1032/2007/(H5N1)) are readily available at the Los Alamos National Laboratory website. Both were examined for conserved region, mutation sites, secondary structural change, hydrophobicity and post-translational modification behaviour. These were later subjected to homology modelling and a phylogenetic tree analysis was conducted to check the strain`s relationship to other strains.
MATERIALS AND METHODS
This in silico study was conducted in 2007 at the Laboratory of Bioinformatics, Department of Chemistry, Faculty of Science, University of Indonesia.
Haemagglutinin and Neuraminidase of Influenza A Virus Sub-Type H5N1 of A/Indonesia/CDC1032/2007 Strain
Haemagglutinin and neuraminidase amino acid sequence of influenza A virus sub-type H5N1 of A/Indonesia/CDC1032/2007 was downloaded in GenBank Flat File (GBFF) format at Los Alamos National Laboratory website (http://www.ncbi.nlm.nih.gov). Other haemagglutinin and neuraminidase amino acid sequences from Indonesian influenza A virus sub-type H5N1 isolates were also collected from this source.
Database Similarity Screening
Amino acid sequences of all Indonesian influenza A (H5N1) virus isolates were screened for 99-100% homology with A/Indonesia/CDC1032/2007 strain using online Basic Local Aligment Search Tool (BLAST) for proteins at National Centre for Biotechnology Information (NCBI) website (http://www.ch.embnet.org/software/BLASTp.html).
Conserved Region Prediction and Mutation Analysis of Influenza A Virus Sub-Type H5N1 of A/Indonesia/CDC1032/2007 Strain
Conserved region is a similar or identical region of sequences. In order to find the conserved region, we used a multiple sequence alignment (MSA) method which aligned three or more biological sequences (protein, DNA, or RNA) using ClustalW and BioEdit 7.0.1 programs. ClustalW program is a multiple sequence alignment program which is created by the European Molecular Biology Laboratory (http://www.ebi.ac.uk/Tools/clustalW/index.html) and BioEdit 7.0.1 program is a biological sequence alignment editor which is created by Tom Hall Ibis Biosciences (http://www.mbio.ncsu.edu/BioEdit/BioEdit.html). ClustalW and BioEdit 7.0.1 programs calculate the best match for the selected sequences and line them up, so that the identities, similarities and differences can be seen. The similar of each amino acid or nucleotide are marked by asteriks (*) and the difference sequences are marked by gab (-). Mutation analysis was conducted using AminoTrackTM toolbox at http://apps.sbri.org/AminoTrack/. AminoTrackTM is a web based tool designed to increase the efficiency with which sequence data is recorded for further analysis. This program is used to identify mutations in viral proteins as these proteins evolve during infection (Mahalanabis et al., 2006).
Secondary Structure, Hydrophobicity and Post-Translational Modification Prediction
Secondary structures were predicted using NNPREDICT at http://www.cmpharm.ucsf.edu/~nomi/nnpredict.html. Hydrophocity prediction was conducted online using ProtScale at http://expasy.org/tools/ProtScale.html. Post-translational modifications were searched using ScanProsite at http://expasy.org/tools/ScanProsite.html.
Homology Modeling
Homology modeling was carried out by comparing influenza A virus sub-type H5N1 of A/Indonesia/CDC1032/2007 strain protein with this available database at Protein Data Bank.
Phylogenetic Analysis
Phylogene of influenza virus type A (H5N1) was presented using TreeView program.
RESULTS AND DISCUSSION
Sequence Analysis
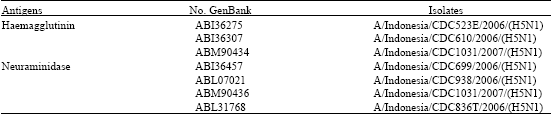
Amino acid sequences of influenza A virus sub-type H5N1 of A/Indonesia/CDC1032/2007 strain was downloaded by the GenBank Accession number of ABM90478 for haemagglutinin sequences and ABM90480 for neuraminidase sequences. The 99-100% homology screening yielded 3 homologs of ABM90478 and 4 homologs for ABM90480 (Table 1).
Conserved Region Prediction and Mutation Analysis of Influenza A Virus Sub-Type H5N1 of A/Indonesia/CDC1032/2007 Strain
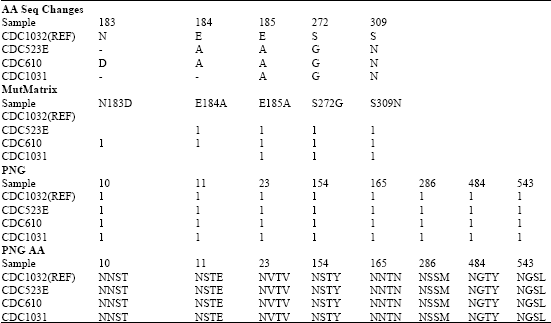
Conserved regions are determined to be at position 1-552 for haemagglutinin and at position 1-39, 41-251 and 253-449 for neuraminidase. Point mutations are observed at position 183, 184, 185, 272 and 309 for haemagglutinin (Table 2a) and at position 40, 63, 239, 244 and 252 for neuraminidase (Table 2b), however specific mutations occurring only to A/Indonesia/CDC1032/2007 strain are at position 185, 272 and 309 for haemagglutinin and 244 and 252 for neuraminidase. The mutation analysis using AminoTrackTM also extracted information on potential n-glycosylation (PNG and PNG AA) sites. However, none of the above mentioned specific mutation occurred at any potential n-glycosylation site. In addition, all mutation, except at position 40 and 252 for neuraminidase, occurred outside the conserved region.
Secondary Structure, Hydrophobicity and Post-Translational Modification Prediction
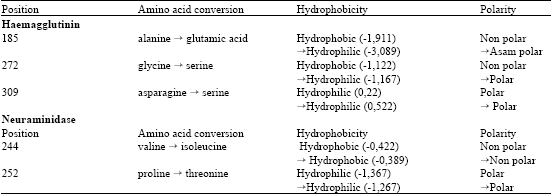
In order to better understand what sort of changes this mutations possibly might have on the A/Indonesia/CDC1032/2007 strain, secondary structure, hydrophobicity and post-translational modification in this mutation site are examined. Secondary structures of haemagglutinin and neuraminidase amino acids are presented at Table 3. For haemagglutinin, helix changes into coil structure occur at position 309, while for position 185 and 272, no secondary structure change is detected. For neuraminidase, coil changes into extended strand structure at position 244, while for 252, no secondary structure change is detected. Hydrophobicity is evaluated using ProtScale software. This software displays the polarity of the initial amino acids and amino acids after the mutation (Table 4).
| Table 1: | A/Indonesia/CDC1032/2007 (H5N1) strain |
 | |
| Table 2a: | Mutation analysis of A/Indonesia/CDC1032/2007(H5N1) strain haemagglutinin using AminoTrack |
 | |
| Table 2b: | Mutation analysis of A/Indonesia/CDC1032/2007 (H5N1) strain neuraminidase using AminoTrack |
 | |
Haemagglutinin sequence at position 185, hydrophobic alanine is replaced by hydrophilic glutamic acid, resulting in a non-polar into polar transition, at position 272, glysine is replaced by serine, resulting in non-polar to polar transition and at position of 309, asparagine is replaced by serine, resulting the insignificant polar into very slightly more polar transition. For neuraminidase, both mutations (244 and 252) cause insignificant change of hydrophobicity. Post-tanslation modification analysis is shown in Table 5, where no modification is observed to occur at any of the mutation sites (Table 5).
| Table 3: | Secondary structure prediction of A/Indonesia/CDC1032/2007 (H5N1) strain haemagglutinin and neuraminidase |
 | |
| Table 4: | Hydrophobicity prediction of A/Indonesia/CDC1032/2007 (H5N1) strain |
 | |
| Table 5: | Post-translational modification prediction of A/Indonesia/CDC1032/2007 (H5N1) strain |
 | |
 | |
Homology Modeling
Homology modelling at Swiss-model server yielded PDB 2fk0 for haemagglutinin and PDB 2hty for neuraminidase. Protein Data Bank database searching revealed that this virus isolate is still similar in structure to Vietnam isolate (A/Vietnam/1203/2004 (H5N1)).
Phylogenetic Analysis
Phylogenetic tree analysis places avian influenza A virus sub-type H5N1 of A/Indonesia/CDC1032/2007 strain in separate cluster to human influenza.
CONCLUSION
Present study revealed that all mutation occur outside the conserved region, except at position 40 and 252 for neuraminidase. In addition, there is no post-translational modification occur at any of the mutation sites. The in silico study cannot prove that A/Indonesia/CDC1032/2007 (H5N1) strain is not a totally new strain. We conclude that the existing mutations in haemagglutinin and neuraminidase might only be a case of antigenic drift. Based on the phylogenetic tree analysis and 3-dimensional homology modelling, the mutation is not significant and an influenza A (H5N1) virus of Banten Province strain can not spread from human to human.
ACKNOWLEDGMENTS
We would like to thank Dr. Ridla Bakri for his support and comments on the manuscript. This research was supported by grants from Ministry of Education (Grant Hibah A3 No. 60/DK-A3/06/2007).
REFERENCES
- Anwar, T., K. Lal Sunil and U. Khan, 2006. In silico analysis of genes Nucleoprotein, Neuraminidase and Hemagglutinin: A comparative study on different strains of influenza A (Bird Flu) virus Sub-type H5N1. J. Silico Biol., 6: 161-168.
PubMedDirect Link - Bridges, C.B., J.M. Katz, W.H. Seto, P.K.S. Chan, D. Tsang and W. Ho et al., 2000. Risk of influenza A (H5N1) infection among health care workers exposed to patients with influenza A (H5N1), Hong Kong. J. Infect. Dis., 181: 344-348.
Direct Link - Glaser, L., J. Stevens, D. Zamarin, A.I. Wilson and G.A. Sastre et al., 2005. A single amino acid substitution in 1918 influenza virus haemagglutinin changes receptor binding specifity. J. Virol., 79: 11533-11536.
Direct Link - Katz, J.M., W. Lim, C.B. Bridges, T. Rowe and J.H. Primmer et al., 1999. Antibody response in individuals Infected with avian influenza A (H5N1) viruses and detection of anti-H5 antibody among household and social contacts. J. Infect. Dis., 180: 1763-1770.
PubMedDirect Link - Kawaoka, Y., S. Krauss and R.G. Webster, 1989. Avian-to-human transmission of the PB1 gene of influenza A viruses in the 1957 and 1968 pandemics. Virology, 63: 4603-4608.
PubMedDirect Link - Mahalanabis, M., J. Blue and N.L. Haigwood, 2006. AminoTrack™: Automating the entry and analysis of mutations in multiple protein sequences using a spreadsheet format. Proceedings of the International Conference on Bioinformatics and Computational Biology (BIOCOMP'06), November 15, 2006, CSREA Press, Las Vegas, pp: 549-555.
- Ungchusak, K., P. Auewarakul, S.F. Dowell, R. Kitphati, W. Auwanit and P. Puthavathana, 2005. Probable person to person transmission of avian influenza (H5N1). N. Engl. J. Med., 352: 333-340.
Direct Link