
Rashad Mukhtar Balal
Department of Horticulture, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Muhammad Mansoor Javaid
Department of Agronomy, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Muhammad Adnan Shahid
Department of Horticulture, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Hira Tariq
Nuclear Institute of Agriculture and Biology, Faisalabad, Pakistan
Muhammad Zubair
Department of Horticulture, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Ahsan Akram
Department of Horticulture, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Gulzar Akhtar
Department of Horticulture, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Muhammad Wajid Khan
Department of Horticulture, University College of Agriculture, University of Sargodha, Sargodha-40100, Pakistan
Science International
Year: 2016 | Volume: 4 | Issue: 2 | Page No.: 30-38







ABSTRACT
In present study, four tomato genotype i.e., Roma, Nagina, Naqeeb and Riogrande were investigated under the saline conditions and the half strength Hoagland’s nutrient solution was provided after a regular interval of time. Results revealed that increased level of salinity significant decreased the number of leaves per plant, root and shoot fresh and dry weight, net photosynthesis rate, chlorophyll content, leaf area and number of stomata, while increase in the concentration of organic osmolytes (total free amino acids and total soluble sugars) was observed in Roma, Nagina, Naqeeb and Riogrande tomato genotypes. The calcium content in root, shoot and leaf markedly reduced in all tested tomato genotypes under saline conditions.
PDF Abstract XML References Citation
How to cite this article
Rashad Mukhtar Balal, Muhammad Mansoor Javaid, Muhammad Adnan Shahid, Hira Tariq, Muhammad Zubair, Ahsan Akram, Gulzar Akhtar and Muhammad Wajid Khan, 2016. Plant Growth is Highly Associated with the Concentration of Organic and Inorganic Osmolytes, Antioxidant Activities in Salt Stress Tomato (Lycopersicon esculentum) Plants. Science International, 4: 30-38.
URL: https://scialert.net/abstract/?doi=sciintl.2016.30.38
URL: https://scialert.net/abstract/?doi=sciintl.2016.30.38
INTRODUCTION
The plants that have to stay on the one place for their whole life so they have to adopt it according to the climatic changes that may be beneficial or harmful for plant growth and development1. There are number of environmental factors which decreased the crop production such as drought, heat, frost, lodging and salinity etc., but in these factors soil salinity dominated on all of the remaining factors worldwide for its prominent marked effects on the crop plants2,3. Among the factors, salt stress is one of the most significant abiotic stress and considers as a sign of disaster for the crop plants2 and is increasing day by day throughout the globe3,4.
In saline soil, when seed is sown very few seedlings emerge5. Salt stress commonly changes the morphological characters of a plant6. Salinity caused a remarkable decrease in the root and stem dry weights and permutation of osmotic and specific ion effects of Cl- and Na+ might possibly caused a decline in the plant biomass7-9. Hajer et al.10 found that shoot and root dry weight of tomato plants significantly decreased with an increase in NaCl concentration, depicting a negative correlation between the same.
Salt stress inhibits the growth and metabolic processes of plants by osmotic stress, ionic stress and oxidative stress11. Furthermore, cell membrane stability, chlorophyll content and photosynthetic activity are significantly hampered under saline conditions12. Salinity affected the net photosynthesis rate by reducing the diffusion of CO2 in respiration process through stomata12. Under salt stress, total quantity of free amino acids increased in tomato fruits13. Ashraf and Harris14 revealed that the different level of NaCl treatments affect the plant dry weight, number of stomata and relative water potential in tomato leaves. Similar results have also been pragmatic by Zushi and Matsuzoe15.
Plant physiology and metabolism influence negatively due the high salt ratios16 and mostly by Na+ salts which not simply decreased Ca2+ availability but also reduced Ca+2 mobility to emergent parts of the plant, that reduce the productive growth of plant parts. In response of high salinity the Na+ and Cl- accumulates above its optimum level causes the deficiency of essential nutrients K+ and Ca+2 which are necessary for plant growth17. Cuin et al.18 observed that for osmotic regulation in tomato different soluble sugars such as fructose, glucose and sucrose increased in response of salt stress. Similarly,19 stated that during salt stress sugars accumulated in different plant tissues and this accumulated sugars play an important role in plant defensive mechanism through osmoregulation process and energy conservation to increase plant tolerance against salinity.
The amino acid behaved as an osmo protectants and enhanced the osmotic efficiency of plants in the situation where the toxic ions of salts create the problems related to water relations20. Morsy et al.21 reported that total free amino acids and soluble protein increased in barley plants as the salt concentration increased in soil. Dantas et al.22 and El-Tayeb23 stated that the numerous enzymes such as S-adenosylmethionine synthetase, glutamine synthetase and isopropylmalate deshydrogenase play an important role in biosynthesis of amino acids these enzymes also help plant in the maintenance of roots osmotic potential and protected the membrane injury during salt stress.
Tomato (Lycopersicon esculentum) is a summer vegetable, consumed worldwide. It is somewhat short duration crop and consider as a cash crop for the small farm holders but unfortunately this cash crop is severely affecting by salinity that reduce its productivity and quality of tomato fruit. However, a significant achievement has been made in improving salinity tolerance in crops by varietal selection12. Many reports on the genotypic variation in commercial cultivars are available regarding osmolytes and antioxidant activity. The objective of this study was to determine the salinity effect on germination, morphological, physiological, biochemical attributes of tomato plant.
MATERIALS AND METHODS
The study was performed and accomplished in 2012 with the collaboration of Nuclear Institute for Agriculture and Biology (NIAB) Faisalabad, Pakistan in Stress Physiology Laboratory, Salinity and Environmental Division.
Plant material and growth conditions: Tomato (Lycopersicon esculentum) genotype i.e., Roma, Nagina, Naqeeb and Riogrande were grown at Institute for Agriculture and Biology (NIAB) Faisalabad in Stress Physiology Laboratory at 25±3°C. Light was provided by fluorescent over-head bulbs set for 8 h light, 16 h dark regime with a photosynthetic photon flux density of 200 μm m–2 sec–1. Plants were grown in 3.5 L pots, filled with sand and manually irrigated with half strength Hoagland nutrient solution. Initially five plants of each variety were sown per pot and were thinned to four plants per pot. When tomato plants have 4-5 true leaf stage, a Hoagland solution with NaCl was added to the Hoagland nutrient solution at 0 and 75 mM. The experiment was laid out in Complete Randomized Design (CRD) with three replications.
Germination of the genotype was recorded after 2 weeks of sowing. Plants were carefully uprooted and washed for the determination of shoot and root fresh weight. Dry weight of plant was measured after drying at 70°C for 72 h. The chlorophyll contents were estimated by using the method of Arnon24.
Leaf Area (LA) was measured using an area meter (LI-COR, model LI-3000). The net photosynthetic rate (Pn), was recorded using an infra-red gas analyze Analytical Development Company, Hoddesdon, England).
For the estimation of total free amino acids, standard method of25 was used while, Total soluble sugars were determined according to the method of26. The calcium contents were determined titrimetrically against EDTA solution (0.01 N) as a standard solution using EBT and calcon indicator as descripbed in US-Staff Hand Book -60 (1962).
RESULTS
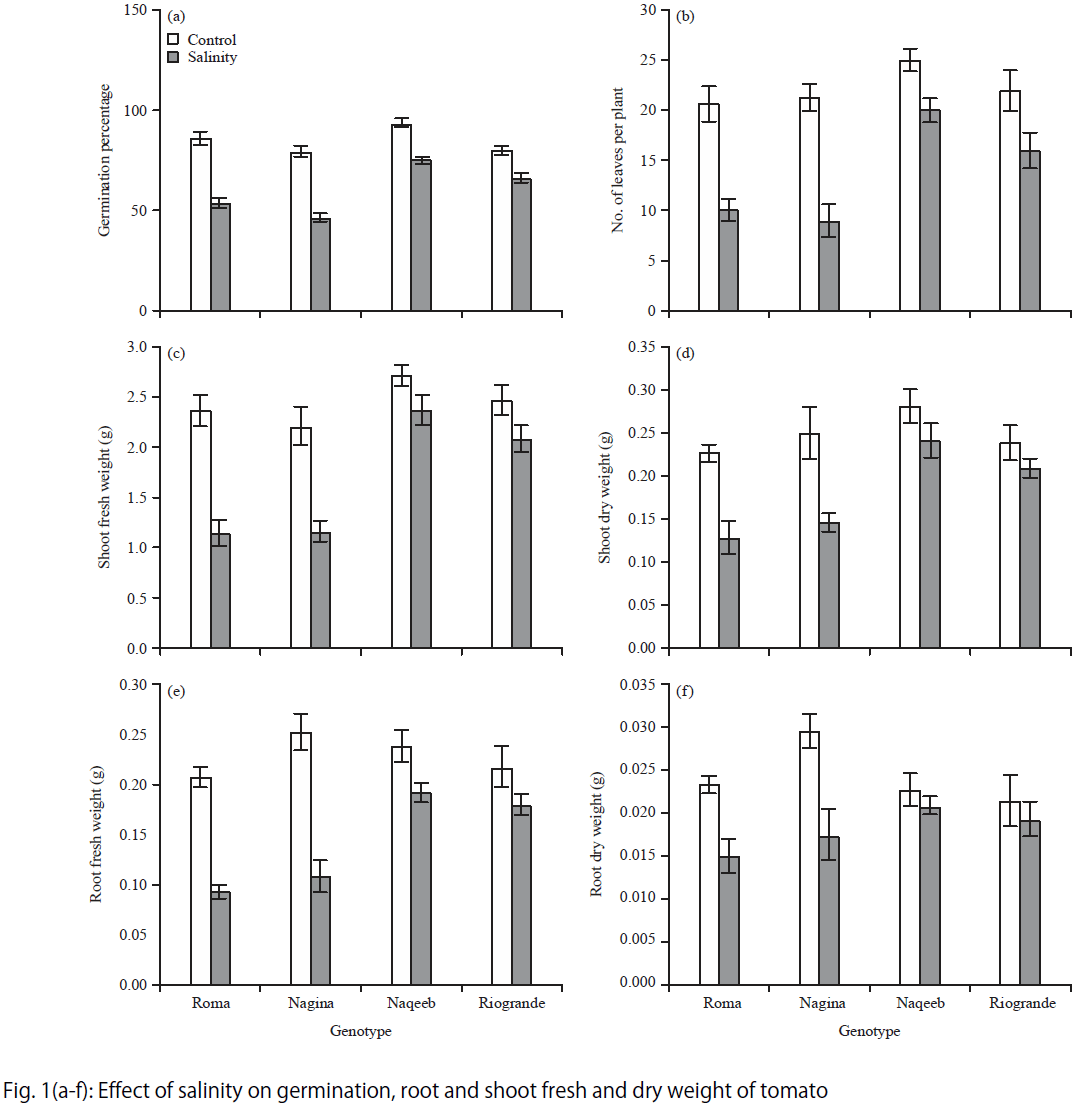
Germination percentage: Data regarding the effect of salt stress on germination percentage showed a significant reduction in the germination percentage of all tested tomato genotypes. The plants growing under non-saline conditions showed the maximum germination percentage while the plants subjected to saline conditions exhibited the minimum germination percentage. The tested genotypes have varied response under saline condition. Naqeeb and Riogrande showed greater salt tolerance and recorded germination more than 65% while, most salt sensitive genotype was Nagina which recorded the germination of 46%.
Plant growth attributes: The number of leaves per plant of tested genotypes indicated that salt stress significantly decreased the number of leaves per plant. However, maximum number of leaves was recorded in Naqeeb followed by Riogrande under saline or non-saline conditions when compared to Roma or Nagina (Fig. 1). The Nagina and Roma presented the reduction in number of leaves by 58 and 52% while Naqeeb and Riogrande had decrease in number of leaves by 20 and 27%.
Salinity significantly reduced the shoot fresh weight of plant. The more pronounced maximum reduction in shoot fresh weight of plant was obtained by the Nagina and Roma, while minimum for Riogrande and Naqeeb (Fig. 1). The Roma and Nagina exhibited the reduction in shoot fresh weight by 51 and 47% while, markedly reduction observed by Naqeeb and Riogrande in shoot fresh weight of plant by 13 and 15%. The four tomato genotype under study showed significant dissimilarity both under saline and non-saline conditions. These four genotype these responded differently in saline conditions. The Nagina and Roma exhibited the reduction in shoot dry weight by 42 and 44% while, Naqeeb and Riogrande showed less reduction in shoot dry weight of plant by 14 and 12%.
There was distinct difference detected between the plant submitted to untreated (control) and salinity (75 mM) because salinity induced the progressive reduction in root fresh weights. The Roma and Nagina exhibited the maximum reduction in root fresh weight by 55 and 57% while, minimum reduction was observed in Riogrande and Naqeeb by 17 and 19%, respectively.
The four tomato genotype under study showed significant variations both under saline and non-saline conditions. Under saline conditions tested tomato genotype responded in a different way when they were compared it to genotype grown under non-saline conditions. The Nagina and Roma exhibited the reduction in root dry weight by 41 and 36% while, Riogrande and Naqeeb presented less reduction in root dry weight by 11 and 8%, respectively.
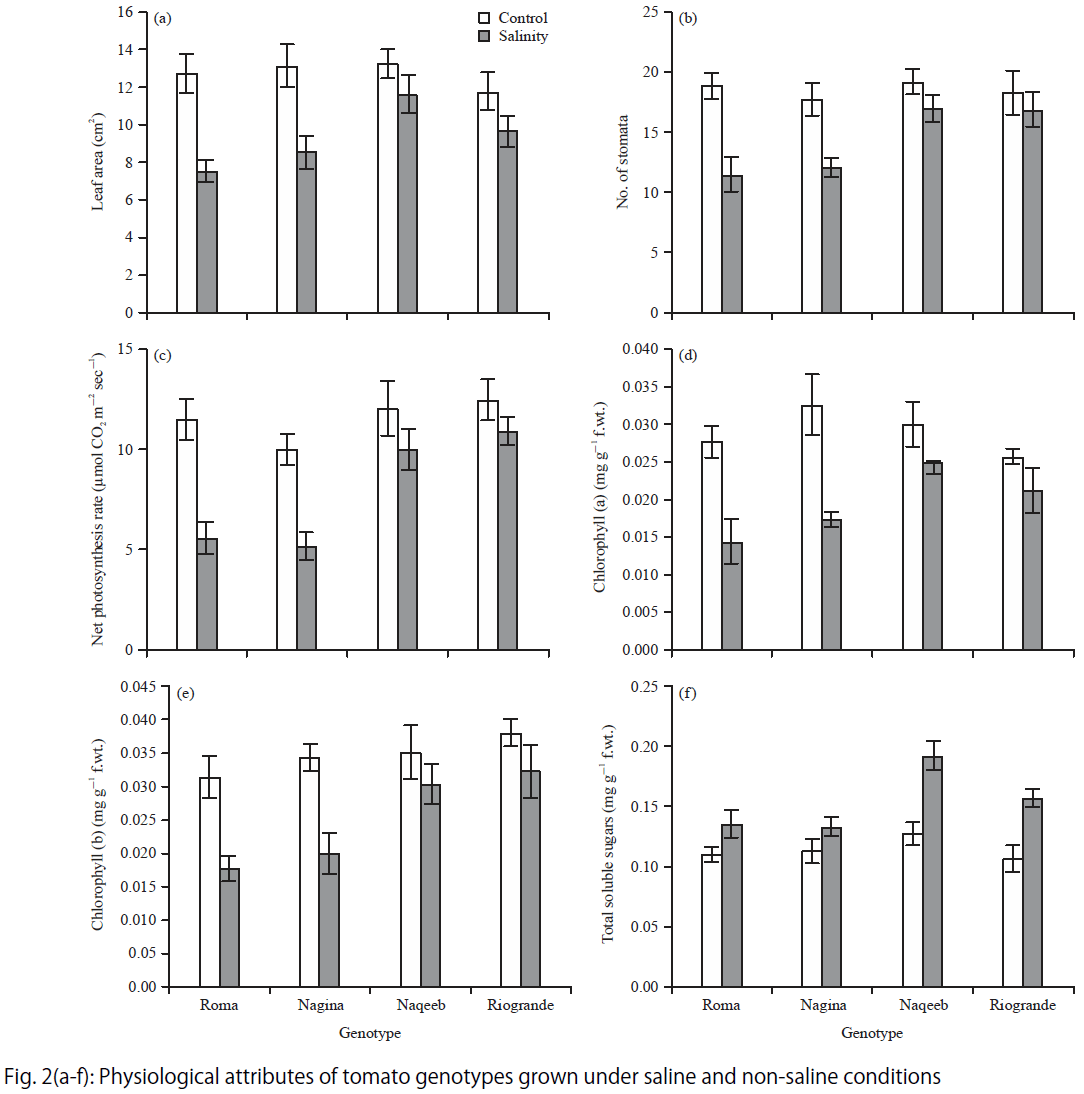
Physiological attributes: The plants growing under non-saline conditions showed the highest net photosynthesis rate while the plants submitted to saline conditions exhibited the minimum net photosynthesis rate (Fig. 2). All the tested genotypes showed the reduction in net photosynthesis rate but maximum reduction was noted for Nagina and Roma while, minimum for Naqeeb and Riogrande. The Nagina and Roma presented reduction in net photosynthesis rate by 49 and 52% while, Riogrande and Naqeeb had decrease in by 12 and 17%.
Salt stress significantly reduced the number of stomata in all tested tomato genotype. The maximum number of stomata of plant observed under non-salinized condition while the minimum number of stomata under saline conditions (Fig. 2). The four tomato genotype under study showed significant dissimilarity both under saline and non-saline conditions. Under salinity the Naqeeb and Riogrande exhibited the minimum decrease in number of stomata by 11 and 8% while maximum decrease showed by Nagina and Roma by 32 and 39%.
Salinity has decreasing effect on the leaf area of all tomato genotype. The tomato genotype performed differently under control and saline conditions (Fig. 2). The genotype displayed the reduction in leaf area but minimum reduction was noted for Riogrande and Naqeeb while, maximum for Roma and Nagina. The Roma and Nagina exhibited the reduction in leaf area by 41 and 35% while, low reduction observed by Naqeeb and Riogrande in leaf water potential of plant by 13 and 18%.
 |
Organic osmolytes
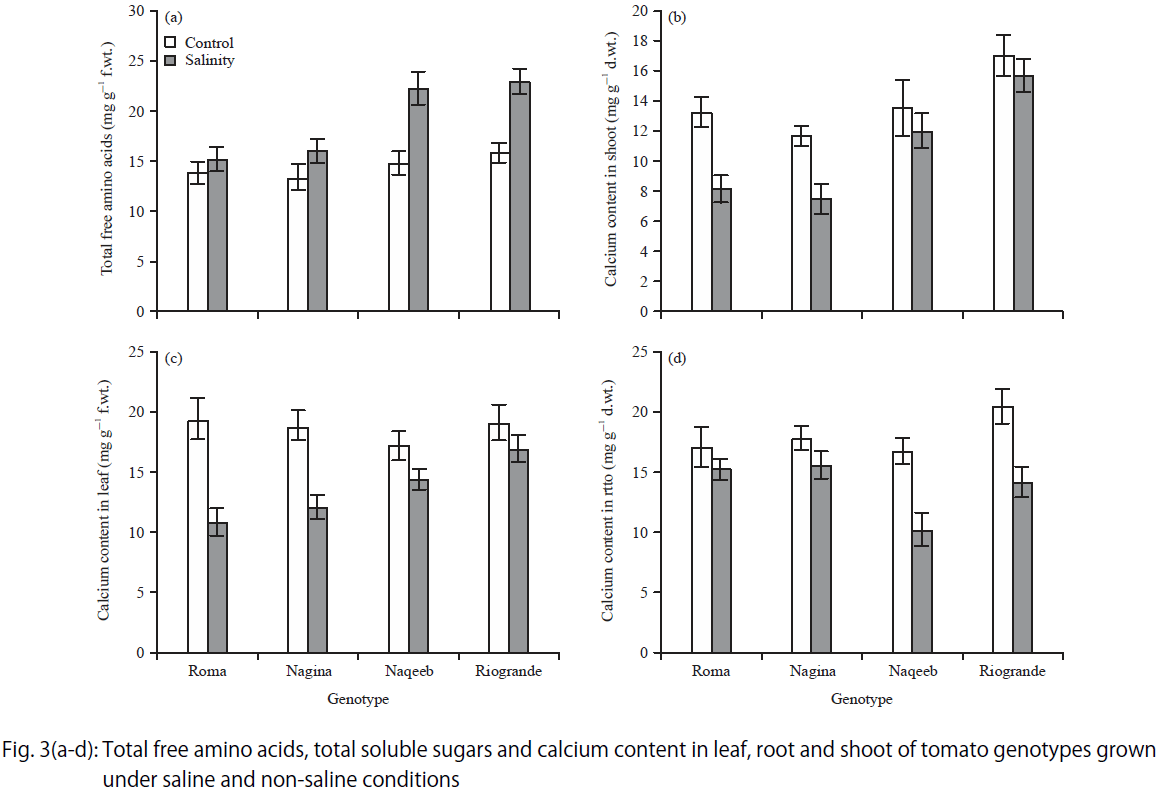
Total free amino acids and total soluble sugars: Under saline conditions a varied pattern in total free amino acids were observed in comparison to non-saline conditions (Fig. 3). All the tested genotype showed the increase in total free amino acids but maximum increase was noted for Riogrande and Naqeeb and minimum increase was observed in Nagina and Roma. The Roma and Nagina presented the minimum increase of total free amino acids by 10 and 20% while, Naqeeb and Riogrande had maximum increase in total free amino acids by 50 and 45%.
Salt stress significantly increased the total soluble sugars in all tested tomato genotype. The maximum total soluble sugars was observed under salinized condition while the minimum total soluble sugar was obtained under control conditions (Fig. 3). The four tomato genotype under study showed significant variation both under saline and non-saline conditions. Observed changes indicated that Naqeeb and Riogrande exhibited the maximum increase of total soluble sugars by 51 and 48% while minimum increase of total soluble sugars showed by Nagina and Roma of 18 and 23%.
 |
Calcium contents: The shoot calcium content indicated that salt stress significantly decreased the calcium content in shoot (Fig. 3). All the tested genotype showed the reduction in calcium content Roma and Nagina presented the reduction in calcium content by 38 and 36% while, Riogrande and Naqeeb had decrease in by 8 and 11%.
It was clearly indicated from the data that under salt stress root calcium content significantly decreased. The plants growing under non-saline conditions showed the high root calcium content while the plants submitted to saline conditions exhibited the low root calcium content (Fig. 3). The Roma and Nagina presented the minimum decrease in calcium content of roots by 11 and 13% while Naqeeb and Riogrande had maximum decrease in calcium content of roots by 39 and 31%. In present study, the plant growing under control and salt stress showed a remarkable difference regarding leaf calcium content. Under saline conditions these genotype responded in a different way when compared it to genotype grown under non-saline conditions. All genotype showed the decrease in leaf calcium content but minimum decrease was observed for Naqeeb and Riogrande while maximum decrease was seen in Roma and Nagina. The Nagina and Roma exhibited the decrease in leaf calcium content by 36 and 44% while, Riogrande and Naqeeb presented minimum decrease in leaf calcium content by 11 and 16%.
 |
DISCUSSION
Salt stress reduced the morphological, physiological and plant growth attributes of tested tomato genotypes. The main effects of salinity was observed as low germination percentage of seeds and reduction in shoot and root lengths27,28. The findings of present study revealed that the germination percentage was significantly decreased under salinity. The salt or excessive ions concentration may be toxic to the seed embryo and there may be excessive salts in the soil to reduce the osmotic potential to such a point which hinder or stop the uptake of water needed for mobilization of nutrient essential for seed germination5. The germination reduced under salt stress, because the salt stress creates the osmotic effect in soil which adversely affects the plant physiology and its optimum nutritional requirements, which is essential for their healthy and successful growth28,29. There are various reports which revealed that salinity has decreasing effect on germination percentage30,31.
From the results it is clear that shoot fresh weight, shoot dry weight, root fresh weight, root dry weight and number of leaves per plant showed a negative correlation with salt stress32 as given in Fig. 1. Plant growth suppressed by the low production and restricted supply of photosynthetic assimilates to leaves that eventually reduced plant growth. The minimum production of fresh and dry shoot and root weight may be due to the disturbance in plant metabolic mechanism that lead a failure in turgor maintenance, nutritional imbalance and ionic stress. Present results are according to the earlier reports shoot dry weight decreased under salt stress reported by Kausar et al.33 and Zehra et al.34.
Salt stress reduced the net photosynthesis rate which adversely influenced the plant growth35. In present study salt stress significantly reduced the net photosynthesis rate of salt tolerant and sensitive tomato genotype but the adverse effects of salinity were more prominent in salt sensitive Roma and Nagina genotype. The toxic ions (Na+ and Cl¯) destroyed the cell organelles involve in the process of photosynthesis, by damaging their membranes of chloroplasts and mesophyll which ultimately reduced the activity of photosynthetic apparatus. Under salt stress these toxic ions adversely affected the net photosynthesis rate and may be reducing the availability of CO236 and irregularities working of the photochemical system in salinized plants37-39.
The salt stress has marked effects on the leaf area and these ions change the metabolic processes of cell wall and these toxic ions accumulated in cell wall that reduced the cell wall flexibility40. Hura et al.41 further explain that due to cell wall rigidity and decreased turgor potential influenced the cell enlargement and ultimately reduced leaf area reduced. These results are in accordance to the findings of Shahid et al.42.
The chlorophyll contents that are present in the plant structure chloroplast and this structure is responsible for the synthesis of chlorophyll contents i.e., chlorophyll (a), chlorophyll (b) and total chlorophyll. Under salt stress the chlorophyll started its degradation because the enzyme cholorophyllase activated under saline conditions43 and in the result chloroplast thylakoid membrane damaged therefore the contents of chloroplast started to come out, the low amount of chlorophyll contents remain in chloroplast that synthesized the very low amount of chlorophyll content. The results are in lined with the findings of Akram et al.44, El-Khallal et al.45 and Boughalleb et al.46.
Salt stress affected the various ionic attributes of tested tomato genotype. The present study investigated the Na+, Cl¯, Ca2+ and K+ in roots, shoot and leaves. It was noted that salinity enhanced the Na+ and Cl¯ accumulation in plant root, stem and leaves. But in case of Ca2+, a decreasing pattern was recorded in tested genotype. Hence, both salt sensitive and salt tolerant genotype exhibited the significant reduction in Ca2+ ions but tolerant genotype maintained the maximum amount of these ions in their leaves as compared to roots while the opposite trend was seen in sensitive genotype. Musyimi47 reported that an antagonistic effect were seen in toxic and beneficial ions. Results of the present finding are according to the reports of Zahoor et al.48 and Nivas et al.49.
Under salt stress tomato genotype affected by the osmotic stress so the plants undergo the process of osmotic adjustment by accumulating different organic osmolytes such as total free amino acids, total soluble sugars, proline and glycinebetaine etc. in various plant parts. Higher the accumulation of organic osmolytes the maximum osmotic adjustment potential within the plants15. Under salt stress the osmoprotectant, total free amino acids naturally produced within the plant parts for maintaining of osmotic adjustment. The positive correlation was observed between total free amino acids and salt tolerance. Similar findings had been reported by El-Khallal et al.45, De Azevedo Neto and Tabosa50 and Bai et al.51.
Total soluble sugars in plants, total soluble sugar production through photosynthesis is a vital process and sugar status modulates the internal regulators and environmental signals to govern growth and development. Thus many plants use sucrose as the major form of transported carbon and it has an important storage role. Sugars also play an important role in regulating the cell metabolism52. These are various reports that showed the accumulation of sugars in different plant parts that increased in response to salt stress53,54. Present study results are compatible with the results of Williams55 who observed that increase in soluble carbohydrate under saline conditions.
CONCLUSION
The results of study revealed that increased level of salinity significant decreased the morphological, physiological attributes of tomato plant. Similarly the calcium content in root, shoot and leaf were markedly reduced in all tested tomato genotypes under saline conditions. While as salinity defense mechanism an increase in the concentration of organic osmolytes (total free amino acids and total soluble sugars) was observed in salt tolerant tomato genotypes i.e., Naqeeb and Riogrande.
REFERENCES
- Gill, P.K., A.D. Sharma, P. Singh and S.S. Bhullar, 2002. Osmotic stress-induced changes in germination, growth and soluble sugar content of Sorghum bicolor (L.) moench seeds. Bulg. J. Plant Physiol., 28: 12-25.
Direct Link - Reynolds, M. and R. Tuberosa, 2008. Translational research impacting on crop productivity in drought-prone environments. Curr. Opin. Plant Biol., 11: 171-179.
CrossRefDirect Link - Eker, S., G. Comertpay, O. Konuskan, A.C. Ulger, L. Ozturk and I. Cakmak, 2006. Effect of salinity stress on dry matter production and ion accumulation in hybrid maize varieties. Turk. J. Agric. For., 30: 365-373.
Direct Link - Foolad, M.R., 2004. Recent advances in genetics of salt tolerance in tomato. Plant Cell Tissue Org. Cult., 76: 101-119.
CrossRefDirect Link - Jamil, M., L. Deog Bae, J. Kwang Yong, M. Ashraf, L. Sheong Chun and R. Eui Shik, 2006. Effect of salt (NaCl) stress on germination and early seedling growth of four vegetables species. J. Cent. Eur. Agric., 7: 273-282.
Direct Link - Mahmoodzadeh, H. and N.M. Bemani, 2007. Effects of salinity on the morphology and yield of two cultivars of canola (Brassica napus L.). J. Agron., 6: 409-414.
Direct Link - Maggio, A., G. Raimondi, A. Martino and S. de Pascale, 2007. Salt stress response in tomato beyond the salinity tolerance threshold. Environ. Exp. Bot., 59: 276-282.
CrossRefDirect Link - Hajer, A.S., A.A. Malibari, H.S. Al-Zahrani and O.A. Almaghrabi, 2006. Responses of three tomato cultivars to sea water salinity 1. Effect of salinity on the seedling growth. Afr. J. Biotechnol., 5: 855-861.
Direct Link - Zrig, A., H.B. Mohamed, T. Tounekti, M. Ennajeh, D. Valero and H. Khemira, 2015. A comparative study of salt tolerance of three almond rootstocks: Contribution of organic and inorganic solutes to osmotic adjustment. J. Agric. Sci. Technol., 17: 675-689.
Direct Link - Montesano, F. and M.W. van Iersel, 2007. Calcium can prevent toxic effects of Na+ on tomato leaf photosynthesis but does not restore growth. J. Am. Soc. Hortic. Sci., 132: 310-318.
Direct Link - Okhovatian-Ardakani, A.R., N. Mehrabanian, F. Dehghani and A. Akbarzadeh, 2010. Salt tolerance evaluation and relative comparison in cuttings of different pomegranate cultivars. Plant. Soil Environ, 56: 176-185.
Direct Link - Ashraf, M. and P.J.C. Harris, 2004. Potential biochemical indicators of salinity tolerance in plants. Plant Sci., 166: 3-16.
CrossRefDirect Link - Zushi, K. and N. Matsuzoe, 2006. Free amino acid contents of tomato fruit grown under water and salinity stresses. Acta Hortic., 724: 91-96.
CrossRefDirect Link - Kaymakanova, M. and N. Stoeva, 2008. Physiological reaction of bean plants (Phaseolus vulg. L.) to salt stress. Gen. Applied Plant Physiol., 34: 177-188.
Direct Link - Abbas, T., R.M. Balal, M.A. Shahid, M.A. Pervez, C.M. Ayyub, M.A. Aqueel and M.M. Javaid, 2015. Silicon-induced alleviation of NaCl toxicity in okra (Abelmoschus esculentus) is associated with enhanced photosynthesis, osmoprotectants and antioxidant metabolism. Acta Physiologiae Plantarum, Vol. 37.
CrossRefDirect Link - Cuin, T.A., A.J. Miller, S.A. Laurie and R.A. Leigh, 2003. Potassium activities in cell compartments of salt‐grown barley leaves. J. Exp. Bot., 54: 657-661.
CrossRefDirect Link - Tejera, N.A., M. Soussi and C. Lluch, 2006. Physiological and nutritional indicators of tolerance to salinity in chickpea plants growing under symbiotic conditions. Environ. Exp. Bot., 58: 17-24.
CrossRefDirect Link - Flores, P., M. Carvajal, A. Cerda and V. Martinez. 2001. Salinity and ammonium/nitrate interactions on tomato plant development, nutrition and metabolites. J. Plant Nutr., 24: 1561-1573.
CrossRefDirect Link - Morsy, M.R., L. Jouve, J.F. Hausman, L. Hoffmann and J.M. Stewart, 2007. Alteration of oxidative and carbohydrate metabolism under abiotic stress in two rice (Oryza sativa L.) genotypes contrasting in chilling tolerance. J. Plant Physiol., 164: 157-167.
CrossRefDirect Link - Dantas, B.F., L. de Sa Ribeiro and C.A. Aragao, 2005. Physiological response of cowpea seeds to salinity stress. Revista Brasileira Sementes, 27: 144-148.
CrossRefDirect Link - El-Tayeb, M.A., 2005. Response of barley grains to the interactive e.ect of salinity and salicylic acid. Plant Growth Regul., 45: 215-224.
CrossRefDirect Link - Yancey, P.H., M.E. Clark, S.C. Hand, R.D. Bowlus and G.N. Somero, 1982. Living with water stress: Evolution of osmolyte systems. Science, 217: 1214-1222.
CrossRefPubMedDirect Link - Ouyang, B., T. Yang, H. Li, L. Zhang and Y. Zhang et al., 2007. Identification of early salt stress response genes in tomato root by suppression subtractive hybridization and microarray analysis. J. Exp. Bot., 58: 507-520.
CrossRefDirect Link - Yemm, E.W., E.C. Cocking and R.E. Ricketts, 1955. The determination of amino-acids with ninhydrin. Analyst, 80: 209-214.
CrossRefDirect Link - Riazi, A., K. Matsuda and A. Arslan, 1985. Water-stress induced changes in concentrations of proline and other solutes in growing regions of young barley leaves. J. Exp. Bot., 36: 1716-1725.
CrossRefDirect Link - Wolf, B., 1982. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal., 13: 1035-1059.
CrossRefDirect Link - Mane, A.V., B.A. Karadge and J.S. Samant, 2010. Salinity induced changes in photosynthetic pigments and polyphenols of Cymbopogon nardus (L.) Rendle. J. Chem. Pharmaceut. Res., 2: 338-347.
Direct Link - Krishnamurthy, L., R. Serraj, C.T. Hash, A.J. Dakheel and B.V.S. Reddy, 2007. Screening sorghum genotypes for salinity tolerant biomass production. Euphytica, 156: 15-24.
CrossRefDirect Link - Kausar, A., M.Y. Ashraf, I. Ali, M. Niaz and Q. Abbass, 2012. Evaluation of sorghum varieties/lines for salt tolerance using physiological indices as screening tool. Pak. J. Bot., 44: 47-52.
Direct Link - Zehra, A., B. Gul, R. Ansari and M.A. Khan, 2012. Role of calcium in alleviating effect of salinity on germination of Phragmites karka seeds. South Afr. J. Bot., 78: 122-128.
CrossRefDirect Link - Nasri, N., I. Saidi, R. Kaddour and M. Lachaal, 2015. Effect of salinity on germination, seedling growth and acid phosphatase activity in lettuce. Am. J. Plant Sci., 6: 57-63.
CrossRefDirect Link - Abdeshahian, M., M. Nabipour and M. Meskarbashee, 2010. Chlorophyll fluorescence as criterion for the diagnosis salt stress in wheat (Triticum aestivum) plants. Int. J. Chem. Biol. Eng., 3: 184-186.
Direct Link - Nedjimi, B., 2011. Is salinity tolerance related to osmolytes accumulation in Lygeum spartum L. seedlings? J. Saudi Soc. Agric. Sci., 10: 81-87.
CrossRefDirect Link - Munns, R., R.A. James and A. Lauchli, 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot., 57: 1025-1043.
CrossRefPubMedDirect Link - Flexas, J., A. Diaz-Espejo, J. Galmes, R. Kaldenhoff, H. Medrano and M. Ribas-Carbo, 2007. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell Environ., 30: 1284-1298.
CrossRefDirect Link - Souza, R.P., E.C. Machado, J.A.B. Silva, A.M.M.A. Lagoa and J.A.G. Silveira, 2004. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot., 51: 45-56.
CrossRefDirect Link - Hura, T., K. Hura, M. Grzesiak and A. Rzepka, 2007. Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiologiae Plantarum, 29: 103-113.
CrossRefDirect Link - Shahid, M.A., R.M. Balal, M.A. Pervez, T. Abbas, M.A. Aqeel, A. Riaz and N.S. Mattson, 2015. Exogenous 24-epibrassinolide elevates the salt tolerance potential of pea (Pisum sativum L.) by improving osmotic adjustment capacity and leaf water relations. J. Plant Nutr., 38: 1050-1072.
CrossRefDirect Link - Yasar, F., S. Kusvuran and S. Ellialtioglu, 2006. Determination of anti-oxidant activities in some melon (Cucumis melo L.) varieties and cultivars under salt stress. J. Hortic. Sci. Biotechnol., 81: 627-630.
Direct Link - Akram, M., M.A. Malik, M.Y. Ashraf, M.F. Saleem and M. Hussain, 2007. Competitive seedling growth and K+/Na+ ratio in different maize (Zea mays L.) hybrids under salinity stress. Pak. J. Bot., 39: 2553-2563.
Direct Link - El-Khallal, S.M., T.A. Hathout, A.E.A. Ashour and A.A. Kerrit, 2009. Brassinolide and salicylic acid induced growth, biochemical activities and productivity of maize plants grown under salt stress. Res. J. Agric. Biol. Sci., 5: 380-390.
Direct Link - Boughalleb, F.,M. Denden and B.B. Tiba, 2009. Photosystem II photochemistry and physiological parameters of three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea under salt stress. Acta Physiologiae Plantarum, 31: 463-476.
CrossRefDirect Link - Musyimi, D.M., 2007. Photosynthetically active radiation influence on gas exchange parameters of avocado seedlings growing under saline conditions. J. Boil. Sci., 7: 1400-1405.
CrossRefDirect Link - Zahoor, M., R. Khaliq, Z.U. Zafar and H.U.R. Athar, 2011. Degree of salt tolerance in some newly developed maize (Zea mays L.) varieties. Iran. J. Plant Physiol., 1: 223-232.
Direct Link - Nivas, D., D.K. Gaikwad and P.D. Chavan, 2011. Physiological responses of two Morinda species under saline conditions. Am. J. Plant Physiol., 6: 157-166.
CrossRefDirect Link - De Azevedo Neto, A.D. and J.N. Tabosa, 2000. Salt stress in maize seedlings: Part I growth analysis. Revista Brasileira Engenharia Agricola Ambiental, 4: 159-164.
Direct Link - Bai, W.B., P.F. Li, B.G. Li, H. Fujiyama and F.C. Fan, 2008. Some physiological responses of Chinese Iris to salt stress. Pedosphere, 18: 454-463.
CrossRefDirect Link - Long, X.H., Z.R. Huang, Y.L. Huang, J. Kang, Z.H. Zhang and Z.P. Liu, 2010. Response of two Jerusalem artichoke (Helianthus tuberosus) cultivars differing in tolerance to salt treatment. Pedosphere, 20: 515-524.
CrossRefDirect Link - Alia, E.A. and I.A. Gahiza, 2007. Accumulation of amino acids in Anabaena oryzae in response to sodium chloride salinity. J. Applied Sci. Res., 3: 263-266.
Direct Link - Shahid, M.A., R.M. Balal, M.A. Pervez, T. Abbas and M. Ashfaq et al., 2012. Differential response of pea (Pisum sativum L.) Genotypes to salt stress in relation to the growth, physiological attributes antioxidant activity and organic solutes. Aust. J. Crop Sci., 6: 828-838.
Direct Link - Williams, L.E., R. Lemoine and N. Sauer, 2000. Sugar transporters in higher plants: A diversity of roles and complex regulation. Trends Plant Sci., 5: 283-290.
PubMedDirect Link