
Yasser A. Ahmed
Department of Histology, Faculty of Veterinary Medicine, South Valley University, Qena, Egypt
Mahmoud M. Abd-Elhamied
Department of Histology, Faculty of Veterinary Medicine, South Valley University, Qena, Egypt
Gamal K.M. Ali
Department of Anatomy and Histology, Assiut University, Assiut, Egypt
Research Journal of Veterinary Sciences
Year: 2012 | Volume: 5 | Issue: 2 | Page No.: 42-50







ABSTRACT
The aim of the current study was to follow up the histological and histomorphometrical changes of descending rabbit testis during different postnatal-developmental stages (1-day and 1-4-, 8-, 12-, 16-, 20- and 24-weeks). The testes were found at the sublumbar region just after birth, moved into the inguinal canal after 4 weeks, located near the scrotum after 8 weeks and were completely within the scrotum after 12-16 weeks. Paraffin and semi-thin sections were taken from the testis and examined with light microscopy. The testis was surrounded by a fibrous capsule with no smooth muscle fibers until 12 weeks, but smooth muscle fibers were seen after 16 weeks. Testicular parenchyma was made up of solid seminiferous cords with no lumina and the cords had only 2 types of cells; gonocytes and Sertoli cells until 8 weeks. Lumination of the testicular cords started after 12 weeks forming seminiferous tubules. Seminiferous tubules were lined by spermatogonia, spermatocytes and round spermatids, in addition to Sertoli cells. After 16-24 weeks, spermatogenesis was very active and sperms were seen. Leydig cells were small and less active at 1-day postnatal but became large and more active after 16-24 weeks. Many growth parameters of seminiferous cords or tubules including their diameter and the numbers of germ cells and Sertoli cells were recorded. The current study widely covered different points of rabbit’s testicular maturity including the anatomical location and the histological and histomorphometrical changes of descending rabbit testis.
PDF Abstract XML References Citation
Received: March 31, 2012;
Accepted: April 28, 2012;
Published: June 21, 2012
How to cite this article
Yasser A. Ahmed, Mahmoud M. Abd-Elhamied and Gamal K.M. Ali, 2012. Histological and Histomorphometric Changes of the Rabbit Testis During Postnatal Development. Research Journal of Veterinary Sciences, 5: 42-50.
URL: https://scialert.net/abstract/?doi=rjvs.2012.42.50
URL: https://scialert.net/abstract/?doi=rjvs.2012.42.50
INTRODUCTION
The testis, the male gonad, consists mainly of seminiferous tubules and interstitial tissue containing Leydig cells, demarcated from outside by a thick vascular connective tissue tunica albuginea, sending septa that subdivide the cavity of the testis into smaller incomplete compartments. Seminiferous tubules are the site of spermatogenesis, while Leydig cells are responsible for secreting male sex hormones, testosterone to maintain spermatogenesis (Eurell and Frappier, 2006).
Mammalian spermatogenesis is a developmental complex process that occurs in the seminiferous tubules producing spermatozoa or sperms able to penetrate and fertilize a healthy oocyte produced by the ovary of corresponding female species (Miles and Western, 2012). The seminiferous epithelium contains numerous stages in the developmental germ cells; spermatogonia, spermatocytes and spermatids that lead to mature spermatozoa (Curtis and Amann, 1981). Spermatogonia, the diploid stem cell from which the rest of the cell types are derived, are found peripherally and can be identified by their densely stained round centrally located nuclei. Some spermatogonia remain as undifferentiated stem cells and continually supply new spermatogonia, while the others divide by meiosis to give the haploid primary spermatocytes. Primary spermatocytes, the initial product of spermatogonial maturation, are large cells with a round nucleus filled with clumped chromatin material and usually seen in various stages of the meiotic division. Secondary spermatocytes are haploid and very short-lived rarely seen cells that quickly undergo meiosis to give spermatids. Spermatids are haploid cells observed in different stages of maturation. Some are square cells with nuclei beginning to get quite dense. Others more advanced have tails beginning to develop. In some stages, the excess cytoplasm is being cast off. The spermatids complete differentiation process, spermiogenesis, to become mature sperms with its characteristic structure; head and tail releasing their tails into the lumen (Amann, 2008).
Adjacent to the seminiferous epithelium the elongated Sertoli cells located perpendicular to the basal lamina of the seminiferous tubules. Sertoli cells support the germline cells during different stages of spermatogenesis. They are tall columnar cells with large pale oval nucleus containing distinct nucleolus and attached to the basal lamina of the seminiferous tubules (Griswold, 1998). Sertoli cells have many cytoplasmic processes that wrap around the germ cells forming blood-testis barrier through network of junction complex between their cytoplasmic processes and the germ cells; this barrier divide the seminiferous tubules into basal and adluminal compartments to separate the haploid cells from the blood immune system (Merchant-Larios and Taketo, 1991). Furthermore, Sertoli cells phagocytose excess cellular materials and apoptotic bodies (Barone et al., 2004). Unlike the germ cells, Sertoli cells are long lived cells and normally not dividing (Johnson et al., 2008). Cytoplasm of Sertoli cells has numerous mitochondria with tubular cristae, abundant smooth endoplasmic reticulum, free ribosomes, prominent Golgi apparatus and many phagocytosed materials (Lebelo and van der Horst, 2010).
Leydig cells, the androgen producing cells are located in the interstitial vascular connective tissue between the seminiferous tubules (Diagone et al., 2012). They have different shapes; they are cuboidal, polygonal or elongated groups of cells with ovoid nucleus (Gondos et al., 1977). Their cytoplasm is acidophilic and has many lipid droplets (Diagone et al., 2012), abundance of smooth endoplasmic reticulum, a few short cisternae of rough endoplasmic reticulum, rod-like mitochondria with tubular cristae, vacuoles resulting from lipid extraction, filaments, microtubules and glycogen (Kerr, 1991).
The aim of the current study was to investigate the histological and histomorphometrical changes of the descending rabbit testis during the postnatal development.
MATERIALS AND METHODS
Animals: Testicular specimens were obtained from 27 healthy New Zealand white rabbits during different stages of postnatal development; 1-day, 1-, 4-, 8-, 12-, 16-, 20- and 24-weeks. The animals were raised in the animal house of the Faculty of Medicine, Assiut University, Assiut, Egypt. The study conducted at the period between May 2010 to October 2011.
Anatomical localization of the descending rabbit testis: The position of the rabbit testis during different postnatal developmental stages (from 1-day until 24 weeks of age) was noticed and recorded.
Paraffin embedding: Dissected testes were rapidly fixed in either 4% phosphate-buffered paraformaldehyde (pH; 7.4) or Bouin's solution. Testes were sectioned crossly or longitudinally, dehydrated in ascending grades of ethyl alcohol, cleared in methyl benzoate and embedded in paraffin. Sections (5 μm-thicknesses) were cut, stained with hematoxylin and eosin (H and E) as a general stain and Periodic Acid Schiff’s (PAS) for mucopolysaccharides and examined with a light microscope.
Spurr’s resin embedding: Some specimens were fixed in 2.5% gluteraldehyde/4% buffered formaldehyde, post-fixed in 1% osmium tetroxide, dehydrated through ascending grades of acetone and embedded in Spurr’s resin. Semi-thin sections (0.5 μm-thicknesses) were cut using an ultra-microtome (Leica Microsystems, Wetzlar, Germany), stained with toluidine blue and examined with a light microscope.
Histomorphometrical analysis: Histomorphometrical parameters related to testicular growth such as the diameter of seminiferous tubules and numbers of germ cells and Sertoli cells per seminiferous tubule during different developmental stages were evaluated using an image analyzer system.
Statistical analysis: Statistical differences between groups were evaluated using one way ANOVA. Measurements were taken from 3 different animals. For each comparison, a p-value of less than 0.05 was considered to be significant.
RESULTS
Descending of the rabbit testis: The testes of the neonatal rabbit were found at the sublumbar region on both sides of the descending colon. One week postnatal, the testes migrated slightly caudally to lie in both sides of the rectum just caudal to the kidneys. Four weeks postnatal, testes began to descend within the inguinal canal and were mostly extra-abdominal near to the scrotum after 8 weeks, but completely locating within the scrotum after 12-16 weeks (Fig. 1).
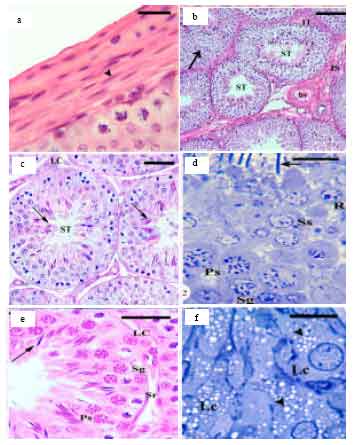
Histological structure of the descending rabbit testis: The testis of the rabbit during early stages of development (1 day to 8 weeks) was surrounded by a thick dense connective tissue tunica albuginea containing a deep vascular layer and covered by flattened mesothelial cells. No smooth muscle cells could be demonstrated during this developmental stage (Fig. 2a).
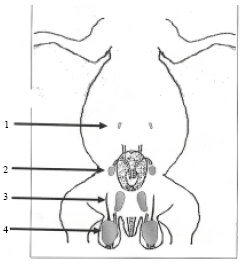 | |
| Fig. 1: | A diagram of the anatomical location of the descending rabbit testes, the testes were located in 1: Sublumbar region one-postnatal-day, 2: The inguinal canal 4-postnatal-weeks, 3: Near to the scrotum 8-postnatal-weeks, 4: Completely within the scrotum 12-16-postnatal weeks |
 | |
| Fig. 2(a-f): | Photographs rabbit testis during early postnatal development Paraffin (a, b, c, e) and semi-thin (d, f) sections stained with H&E (a, b, c, e) and toluidine blue (d, f) were taken from the testis of 1-day-old (a, b, c, d) and 12-week-old (e, f) rabbits, (a) Tunica albuginea (TA) containing wide blood vessels (BV), (b) Seminiferous cords (SC) containing Gonocytes (G) and Sertoli cells (S), separated by interstitial connective tissue (IT) having Leydig cells (LC), (c) Gonocytes (G) in mitotic division (arrowhead) and Sertoli cells (S), (d) Leydig cells (Lc) containing cytoplasmic vacuoles (arrowheads) surrounding blood capillaries (bc), (e) Seminiferous tubules having lumen (ST) surrounded by tunica albuginea (TA) with large blood vessels (bv) sending testicular septae (TS), (f) Spermatogenic cells; spermatogonia (Sg), primary spermatocytes (Ps), rounded spermatids (Rs) surrounded by interstitial Leydig cells (Lc), Bars = 10 μm in a, c, d, f, 50 μm in b and 100 μm in E |
Testicular parenchyma consisted of solid straight testicular cords with no lumina, widely separated from each others by an abundant interstitial tissue and the cords were demarcated by a well developed basal lamina (Fig. 2b). The testicular cords contained two types of cells; pre-spermatogenic cells or gonocytes and Sertoli cells (Fig. 2c). The pre-spermatogenic cells were large rounded cells with rounded nucleus having two distinct nuclei and pale stained cytoplasm and frequently seen in mitotic division (Fig. 2c). These cells were seen in the center of the testicular cords and had no contact to the basal lamina. While, the Sertoli cells were tall columnar with oval or spindle-shaped nucleus and pale cytoplasm and their boundaries were less clear (Fig. 2c). The interstitial tissue contained small singly scattered or grouped spindle, oval, triangular, polyhedral or elongated Leydig cells, which contained small deeply stained nucleus and pale vacuolated cytoplasm due to the presence of lipid droplets (Fig. 2d).
After 12 weeks, seminiferous cord lumination started and the cords termed as seminiferous tubules (Fig. 2e). Seminiferous tubules had no gonocytes, but were lined with stratified germinal epithelium including spermatogonia, spermatocytes and spermatids, in addition to Sertoli cells (Fig. 2f). Spermatogonia were peripherally located small, rounded or oval shaped cells with relatively large, vesicular nucleus containing one or two distinct nucleoli. Primary spermatocytes were the largest cells among the seminiferous epithelium and had rounded nucleus with distinct nucleolus. Secondary spermatocytes were smaller than primary spermatocytes and rarely seen. Spermatids were the smallest germ cells and they are rounded cells with rounded nuclei.
After 16-24 weeks, when the animals reached sexual maturity, the rabbit testis was surrounded by tunica albuginea containing smooth muscle fibers (Fig. 3a). Lumination of the seminiferous tubules increased, the tubules were surrounded by a clear PAS-positive basal lamina and some of them appeared fused together (Fig. 3b).
 | |
| Fig. 3: | Photographs of the rabbit testis during late postnatal development, paraffin (a, b, c, e) and semi-thin (d, f) sections stained with H and E (a, e), PAS and H and E (b, c) and toluidine blue (d, f) were taken from the testis of 16-week-old (c, e, f), 20-week-old (B) and 24-week-old (a, d) rabbits. (a) Smooth muscle fibers (arrowheads) in the tunica albuginea. (B) Seminiferous tubules with PAS-positive basal lamina (ST), some of seminiferous tubules fused together (arrows); note interstitial tissue (IT), blood vessels and intratubular septa (IS). (c) Seminiferous tubules rest on PAS-positive basal lamina and having PAS-positive spermatozoa (arrow). (d) Active spermatogenesis; spermatogonia (Sg), primary spermatocytes (Ps), secondary spermatocytes (Ss), rounded spermatids (R) and spermatozoa (arrow). (e) Seminiferous tubules with wide lumen lined by spermatogonia (Sg), primary spermatocytes (Ps), and spermatozoa (arrow), Sertoli cells (Sr) and Leydig cells (LC). (f) Large active Leydig cells (Lc) containing numerous various sizes and shapes vacuoles (arrowheads). Bars = 10 μm in A, D, E, 100 μm in B, 50 μm in C and 25 μm in E |
 | |
| Fig. 4(a-f): | Histomorphometric changes in the rabbit testis during postnatal development, the diameter of seminiferous tubules (a), the numbers of spermatogenic cells (b, c, d) including gonocytes (a), spermatogonia (b), Primary spermatocytes (c) spermatids (d) Sertoli cells (f) were estimated using image analyzer system. Each column represents mean±SE. Results were considered significant when p-value<0.5 |
Spermatogenesis was very active and all cell types including sperms were seen during this stage (Fig. 3c, d). The elongated Sertoli cells markedly increased in size (Fig. 3f) and Leydig cells were rounded, oval, polyhedral or elongated in shape with large oval nucleus and cytoplasm having a few and numerous small lipid vacuoles (Fig. 3e).
Histomorphometrical studies of the rabbit testis: The histometric data of the present study showed that the diameter of the seminiferous cords of testis of newborn rabbits was 133.3±44.7 μm and nearly remained unchanged until the age of 8 weeks but it increased significantly increased after 24 weeks to reach 450.78±150.53 μm (Fig. 4a). Gonocyte number was 7.7±3.3/seminiferous cords at 1 day-postnatal, gradually decreased to reach 1.7±0.4 after 8 weeks and then completely disappeared from the seminiferous tubules after 12 weeks (Fig. 4b). Spermatogonia had never been seen before the 8 weeks of postnatal development, but 22.7±2.4 spermatogonia were found after 12 weeks and then gradually decreased to 14.2±0.5 after 24 weeks (Fig. 4c). Primary spermatocytes (37.2±4.9) were firstly detected after 12 weeks and increased later on reaching 76.4±3.67 after 24 weeks (Fig. 4d). Similarly, spermatids (7.6±4.3) were firstly observed after 12 weeks and then increased to 111.4±23.6 by 24 weeks of age (Fig. 4e). Total spermatogenic cells (obtained from summation of spermatogonia, primary spermatocytes and spermatids) increased from 67.6±6.9 after 12 weeks of age to reach 202.2±27.2 after 24 weeks of postnatal development indicating increased spermatogenesis during this time (data not shown). Sertoli cell number was 32±1.6 after birth and gradually decreased to reach 5.4±0.7 after 24 weeks of age (Fig. 4f).
DISCUSSION
The current study was conducted to follow up the most dramatic histological and histomorphometrical changes associated with the descending process of the rabbit testis during postnatal development.
It was noticed that the descending of the rabbit testis into the scrotum showed 4 sequential phases; in neonatal rabbit, the testis were located at the sublumbar region. Testes were deviated caudally into the inguinal canal after 4 weeks, near to the scrotum after 8 weeks and completely located within the scrotum after 12-24 weeks. This normal way of the testicular descending explained in the other studies and any abnormality in this descending pathway is clinically evident as cryptorchidism (Kaleva and Toppari, 2003; Amann and Veeramachaneni, 2007). Movement of two ligaments appears to play an important role in determining the position of the gonads; the gubernaculum, which develops below the gonad in the inguinal area of the abdominal cavity and the cranial suspensory ligament which develops between the gonads and the dorsal abdominal wall near to the last rib (Hutson et al., 1997).
Histological examination of the rabbit testis during postnatal development showed that the testis of the was surrounded by a thick dense connective tissue capsule, tunica albuginea containing large blood vessels and covered with flattened mesothelial cells. No smooth muscle cells could be demonstrated until 12 weeks of age. However, capsule showed differentiated smooth muscle fibers after 16 weeks, when the testes were completely descended in the scrotum.
Seminiferous tubules were observed as solid cords without a central lumen until the 8th week of postnatal development. Pre-spermatogenic or gonocytes, the only germ cells seen, were located in the centre of the seminiferous cords and surrounded by the Sertoli cells. Similar observation was seen in the sheep testis (Herrera-Alarcon et al., 2007). It is known that the pre-spermatogonia enter waves of mitotic division during the early testicular development and simultaneously they undergo morphological differentiation to form other spermatogenic cells (Curtis and Amann, 1981; De Rooij and Russell, 2000). Lumination of the rabbit testicular cords was demonstrated after 12 postnatal-weeks when the diameter of the seminiferous tubules significantly increased. Appearance of the seminiferous tubule lumen is likely due to accumulation of tubular fluid secreted by the Sertoli cells as an earlier event of spermatogenesis (Russell et al., 1989). The seminiferous tubules of the rabbit testis at 12 weeks of age were lined by a stratified germinal epithelium characterized by the first appearance of the round spermatids in addition to spermatogonia and spermatocytes. During the period 16-24 weeks of postnatal development, the rabbit became sexually mature (Hafez and Kodituwakku, 1970), spermatogenesis was very active as indicated by the increased total number of the spermatogenic cells and sperms were seen. In addition, the presence of contractile smooth muscle fibers within the capsule during this stage of development is important for squeezing the testis allowing moving of the sperms into the clearly formed lumen.
Histomorphometry revealed that the number of the Sertoli cells was higher during the first 8 weeks, decreased at 12 weeks but remained unchanged until 24 weeks postnatal. Similar observation was recorded in previous study (Herrera-Alarcon et al., 2007). It is widely accepted that Sertoli cells are essential in the development and differentiation of germ cells (Shinohara et al., 2003). Sertoli cell proliferation occurs during a limited period comprised between sexual differentiation and puberty as Sertoli cells differentiate to functional adult cells, they loss the capacity of proliferation (Sharpe et al., 2003). Sertoli cells have nutritive, protective and supportive functions for spermatogenic cells phagocytose regressive sperms and produce an androgen-binding protein and secret constituents of intratubular fluid (Cigankova, 1983), which facilitates the lumination of the seminiferous cords. Furthermore, they form the blood-testis barrier (Tindall et al., 1975) to keep the developing haploid spermatogenic cells protected from the action of immune cells of the blood.
Leydig cells were found just after birth and were numerous small scattered or grouped of cells that became larger, more active and had many lipid droplets in the testis of the adult rabbit. These cells are known to secret testosterone to activate the hypothalamo-hypophyseal testicular axis for the continued development of the male reproductive system (Ariyaratne and Mendis-Handagama, 2000; Habert et al., 2001).
CONCLUSION
In conclusion, the current study localized the anatomical position of the rabbit testis during postnatal development. This study has described the histological and histomorphometrical changes of the rabbit testis from 1 day to 24 weeks of postnatal development.
REFERENCES
- Amann, R.P., 2008. The cycle of the seminiferous epithelium in humans: A need to revisit? J. Androl., 28: 469-487.
PubMed - Amann, R.P. and D.N.R. Veeramachaneni, 2007. Cryptorchidism in common eutherian mammals. Reproduction, 133: 541-561.
CrossRefDirect Link - Ariyaratne, H.B. and S.C. Mendis-Handagama, 2000. Changes in the testis interstitium of Sprague Dawley rats from birth to sexual maturity. Biol. Reprod., 62: 680-690.
PubMed - Barone, F., S. Aguanno, A. D'Alessio and A. D'Agostino, 2004. Sertoli cell modulates MAA-induced apoptosis of germ cells throughout voltage-operated calcium channels. Faseb. J., 18: 353-354.
PubMed - Cigankova, V., 1983. Prenatal and postnatal development of Sertoli cells in rams. Vet. Med. (Praha), 28: 613-620.
PubMedDirect Link - Curtis, S.K. and R.P. Amann, 1981. Testicular development and establishment of spermatogenesis in Holstein bulls. J. Anim. Sci., 53: 1645-1657.
PubMedDirect Link - De Rooij, D.G. and L.D. Russell, 2000. All you wanted to know about spermatogonia but were afraid to ask. J. Androl., 21: 776-798.
CrossRefPubMedDirect Link - Diagone, K.V., M.A. Feliciano, M.R. Pacheco and W.R.R. Vicente, 2012. Histology and morphometry of the testes of adult domestic cats (Felis catus). J. Feline. Med. Surg., 14: 124-130.
CrossRef - Gondos, B., K.P. Morrison and R.H. Renston, 1977. Leydig cell differentiation in the prepubertal rabbit testis. Biol. Reprod., 17: 745-748.
Direct Link - Griswold, M.D., 1998. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol., 9: 411-416.
CrossRefPubMedDirect Link - Hafez, E.S.E. and G.E. Kodituwakku, 1970. Postnatal development of the oviduct of intact and ovariectomized rabbits. J. Morphol., 130: 353-365.
CrossRefDirect Link - Herrera-Alarcón, J., E. Villagómez-Amezcua, E. González-Padilla and H. Jiménez-Severiano, 2007. Stereological study of postnatal testicular development in Blackbelly sheep. Theriogenology, 68: 582-591.
CrossRefPubMedDirect Link - Hutson, J.M., S. Hasthorpe and C.F. Heyns, 1997. Anatomical and functional aspects of testicular descent and cryptorchidism. Endocr. Rev., 18: 259-280.
PubMed - Johnson, L., D.L. Thompson Jr. and D.D. Varner, 2008. Role of Sertoli cell number and function on regulation of spermatogenesis. Anim. Reprod. Sci., 105: 23-51.
PubMed - Kaleva, M. and J. Toppari, 2003. Genetics and hormones in testicular descent. Hormones (Athens), 2: 211-216.
PubMed - Kerr, J.B., 1991. Ultrastructure of the seminiferous epithelium and intertubular tissue of the human testis. J. Electron. Microsc. Tech., 19: 215-240.
PubMed - Lebelo, S.L. and G. van der Horst, 2010. The ultrastructure of the Sertoli cell of the vervet monkey, Chlorocebus aethiops. Tissue Cell, 42: 348-354.
PubMed - Merchant-Larios, H. and T. Taketo, 1991. Testicular differentiation in mammals under normal and experimental conditions. J. Elect. Microsc. Tech., 19: 158-171.
PubMed - Miles, D.C. and P.S. Western, 2012. Germ cell sex and cell cycle. Histol. Histopathol., 27: 445-457.
PubMed - Russell, L.D., A. Bartke and J.C. Goh, 1989. Postnatal development of the Sertoli cell barrier, tubular lumen, and cytoskeleton of Sertoli and myoid cells in the rat, and their relationship to tubular fluid secretion and flow. Am. J. Anat., 184: 179-189.
PubMed - Sharpe, R.M., C. McKinnell, C. Kivlin and J.S. Fisher, 2003. Proliferation and functional maturation of Sertoli cells and their relevance to disorders of testis function in adulthood. Reproduction, 125: 769-784.
PubMedDirect Link - Shinohara, T., K.E. Orwig, M.R. Avarbock and R.L. Brinster, 2003. Restoration of spermatogenesis in infertile mice by Sertoli cell transplantation. Biol. Reprod., 68: 1064-1071.
Direct Link - Tindall, D.J., R. Vitale and A.R. Means, 1975. Androgen binding protein as a biochemical marker of formation of the blood-testis barrier. Endocrinology, 97: 636-648.
PubMed