
M.N. Mohamed Fouzi
Centre for Aquatic Animal Disease Diagnosis and Research, Faculty of Veterinary Medicine and Animal Science, University of Peradeniya, Peradeniya 20400, Sri Lanka
Mohamed Shariff
Aquatic Animal Health Unit, Faculty of Veterinary Medicine, Universiti Putra Malaysia, Malaysia
Fatimah Md Yusoff
Institute of Bioscience, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
Research Journal of Veterinary Sciences
Year: 2012 | Volume: 5 | Issue: 1 | Page No.: 13-24







ABSTRACT
Accumulation of ammonia is toxic and can cause stress in cultured animals leading to immune depression and increase susceptibility to diseases in aquaculture systems. The present study was aimed to quantify stress in shrimp exposed to different levels of total ammonia nitrogen, TAN, (8.1, 3.8 and 1.1 mg L-1) for 10 days and subsequently infect with White Spot Syndrome Virus (WSSV) by feeding positive WSSV shrimp. Haemo-chemicals alanine transaminase (ALT), aspartate transaminase (AST), total protein (TP), glucose and electrolytes (Na+, Cl¯, K+) were measured to quantify the stress caused by ammonia and WSSV infection. Shrimp exposed to 8.1 mg L-1 TAN for 10 days revealed significantly high haemolymph ALT, AST and TP while shrimp exposed to 3.8 mg L-1 TAN showed significantly low TP. These changes in haemolymph parameters showed that exposure to 3.8 mg L-1 TAN for 48 h was sufficient to stress the shrimp. Stress in shrimp exposed to ammonia for 10 days further increased significantly when shrimp were infected with WSSV. During post WSSV infection, haeomolymph TP significantly decreased with increase in TAN 1.1, 3.8 and 8.1 mg L-1 treatments. Therefore, exposure of shrimp to higher ammonia level had stressed the shrimp more and the stress further increased by subsequent infection with WSSV as illustrated by changes of haemo-chemical parameters.
PDF Abstract XML References Citation
Received: November 03, 2011;
Accepted: January 14, 2012;
Published: February 23, 2012
How to cite this article
M.N. Mohamed Fouzi, Mohamed Shariff and Fatimah Md Yusoff, 2012. Stress Quantification in Penaeus monodon Exposed to Different Levels of Ammonia and Subsequent Infection to WSSV. Research Journal of Veterinary Sciences, 5: 13-24.
URL: https://scialert.net/abstract/?doi=rjvs.2012.13.24
URL: https://scialert.net/abstract/?doi=rjvs.2012.13.24
INTRODUCTION
Farming of giant tiger shrimp Penaeus monodon is drastically decreasing in the world due to its susceptibility for many viral diseases. Semi-intensive farming of Penaeus monodon may be subjected to environmental variations such as fluctuation of salinity, temperature, dissolved oxygen, turbidity and pH (Chiba et al., 2004; Joseph and Philip, 2007; Pascual et al., 2003; Rahman et al., 2006; Fotedar and Evans, 2011), perturbation of pond bottom, or accumulation of toxic compounds such as ammonia or nitrite (Mugnier et al., 2008; Chen and Cheng, 1995). Environmental variation such as pH, temperature, sodium chloride, carbon etc., influences even on the enzyme activity in the pond water (Poornima et al., 2008). Many other studies showed effect of industrial waste water on the physical and chemical characteristics of the water (Arimoro et al., 2008). These environmental variations are stressful to the culture organisms as they weaken their defence system or often with lethal effects on shrimp in extreme conditions (Chen and Kou, 1991).
The composition of crustacean haemolymph is a good indicator to stressors such as environmental fluctuation, water borne pollutant and pathological agents. Haemolymph metabolic variables such as protein and glucose are found to vary in response to acute salinity changes (Perazzolo et al., 2002; Joseph and Philip, 2007), hypoxia (Hall and van Ham, 1998; Mugnier et al., 2008), extreme temperatures (Pascual et al., 2003), physiological stress (Mugnier et al., 2006), high ambient ammonia (Chen et al., 1994; Mugnier et al., 2008), repeated handling (Mercier et al., 2006) and infections like WSSV (Yoganandhan et al., 2003). Abbas (2006) reported a study that he examined the toxicity of ammonia on Carp fish at different pH levels. Haemolymph transaminases viz. Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) were found to vary in response to ammonia stress in P. monodon (Pan et al., 2003) and WSSV infection in P. indicus (Mohankumar and Ramasamy, 2006). Haemolymph electrolytes also can be altered by changing osmolality and membrane bound phosphatase of the crustaceans in response to some stresses (Chim et al., 2003). Haemocytes and prophenoloxidase in the haemolymph are also sensitive to many physiological and environmental stress factors such as ammonia, hypoxia and salinity changes (Rao et al., 2008; Rodriguez-Ramos et al., 2008; Mugnier et al., 2008; Chang et al., 2007; Perazzolo et al., 2002). Stress therefore disrupts the immune ability and metabolic performance of shrimp, thereby increasing their susceptibility to microbial infection.
Diseases are the major constrain of the aquaculture (Sihag and Sharma, 2012). WSSV is one of the most lethal pathogen of cultured shrimp (Rahman et al., 2006) and their susceptibility could be intensified by acute salinity (Liu et al., 2006), temperature (Rahman et al., 2006), dissolved oxygen and hardness and pH of the water (Kautsky et al., 2000). In spite of many reports on the effect of elevated ammonia which decreases immunocompetence of shrimp (Jiang et al., 2004; Mugnier et al., 2008), the association of shrimp stressed by exposure to ammonia and its subsequent infection by WSSV has been poorly documented except our work on PCR (Fouzi et al., 2010). Hence, present study on P. monodon was therefore aimed (1) to quantify the stress caused by shrimp exposure to different levels of ammonia by determining the haemo-chemical parameters; ALT, AST, total protein, glucose, Na+, K+ and Cl¯ and (2) to determine the association of stress caused by ammonia exposure and subsequent infection of WSSV in shrimp based on the haemo-chemical parameters.
MATERIALS AND METHODS
Shrimp and preparation of ammonia medium: A total of 1000, WSSV-free P. monodon with the body weight of 8.5±1.5 g from a single batch were obtained from a nearby shrimp farm. Twenty-seven shrimp were subjected to Nested Polymerase Chain Reaction (nPCR) assay to screen for WSSV infection. According to veterinary sampling table (Lightner, 1996) the assay would detect infection at 95% probability. The primer and amplification protocol were used according to OIE (2006). Shrimp were first acclimatized for a week in the laboratory tanks at room temperature (28°C) and pH 8.16.
Experimental tanks were prepared and stocked with sterilized sea water with different levels of ammonia by mixing with freshly prepared standard solutions of ammonium sulphate (NH4)2SO4. Standard stock solution of ammonia (1000 mg L-1) was prepared by dissolving 9.434 g of anhydrous grade (NH4)2SO4 in 1000 mL deionized water. Series of standard solutions (100, 10 and 1 mg L-1) were prepared by mixing ammonium ion free seawater. From these standard solutions, tanks were maintained initially at approximately 7, 3 and 1 mg L-1 TAN by controlling the feeding to 0.5% body weight per day divided in two meals. However, there was slight variations in the TAN and reached in average to 8.1, 3.8 and 1.1 mg L-1, respectively. Temperature and pH of the water were constant throughout the experimental period in a closed system.
Experimental protocols: Tanks of 250 L capacity were used for the experiment with density of 40 shrimp per tank. Five groups of tanks each with three replicates (15 tanks) were filled with 180 L sterilized sea water. The treatment tanks were prepared by using the standard stock solution of ammonium sulphate freshly prepared as described above. Each tank was stocked with shrimp and fed with commercial feed twice a day. After stressing the shrimp by exposing to different levels of ammonia for the first 10 days, all groups except for negative control, shrimp were fed approximately 5 g of minced WSSV-infected shrimp. The negative control group was fed with minced healthy shrimp. Shrimp in all treatment tanks except for the control group were starved for 24 h prior to transmission of WSSV to ensure that all shrimp fed on the infected minced shrimp.
Sample collection: During the 10 days ammonia exposure period, haemolymph from shrimp samples (2 shrimp per tank from three replicates of each treatment) were collected from the experimental tanks at 0, 2, 4, 6, 8 and 10 days. The haemolymph was aspirated from the ventral sinus of each shrimp sample into a 1 mL sterile disposable syringe (with 25 gauge needle) containing 0.3 mL of anticoagulant solution (30 mM of tri-sodium citrate, 0.34 mM sodium chloride, 10 mM EDTA, at pH 7.55).
After transmission of WSSV, haemolymph from shrimp samples were (2 shrimp per tank from three replicates of each treatment) collected as described above from experimental tanks at 0, 4, 8, 16, 24, 48, 72, 96 and 120 h post WSSV infection.
In situ measurements for water temperature, pH, dissolved oxygen and salinity in all the tanks was done by multi-parameter probe (YSI-556 MPS, USA) along with the sampling of shrimp. Water samples were also collected to analyze Total Ammonia Nitrogen (TAN) using the method described by Parsons et al. (1984) at every sampling of haemolymph. All water quality parameters were maintained constant except for TAN levels which varied for different treatments. Since ammonia is mainly built up by excess feeding of high protein feed and excreta of shrimp in the culture systems, required ammonia levels were maintained by siphoning the faeces and the feeding rate was minimized to 0.5% of shrimp body weight per day.
Haemo-chemical analyses: Haemolymph parameters were analyzed by the Automatic Analyzer (Hitachi 902, USA) using specific kits for each parameter. Alanine Aminotransferase (ALT) was analysed by using ALT (ALAT/GPT) Roche kit (Cat. No. 10851132) with its measuring range of 4-600 U L-1 (0.07 μkat L-1). Aspartate Aminotransferase (AST) was analysed by using AST (ASAT/GOT) Roche kit (Cat. No. 10851124) with its measuring range of 4-800 U L-1 (0.07 μkat L-1). Haemolymph total protein concentration was analysed by using TP Roche kits (Cat. No. 11553836 316) with its measuring range between 2-150 g L-1. Haemolymph glucose concentration was analysed by using Gluco-quant Glucose/HK kit (Cat. No. 1447513) with its measuring range between 2-750 mg d L-1 (0.11-41.6 mmol L-1). Additional accessory was used to analyse haemolymph electrolytes (Na+, K+ and Cl-1) with the Automatic Analyzer (Model 902). Measuring ranges of Na+, K+ and Cl¯ were 80-450, 1 to 100 and 10-500 mmol L-1, respectively.
Statistical analysis: Comparisons of haemo-chemicals between prior and post infection of WSSV were carried out by student’s t-test using SPSS 15.0. ANOVA was used to compare the means of haemo-chemicals among shrimp exposed to different levels of ammonia both prior and post infection.
RESULTS
Effect of ammonia exposure on haemo-chemicals of P. monodon prior to WSSV infection: Prior to WSSV infection, haemolymph ALT, AST and TP were interestingly different among shrimp exposed to different concentration of TAN, whereas glucose and electrolytes did now any significant differences. Concentration of haemolymph ALT in shrimp exposed to 8.1 mg L-1 TAN was significantly higher (p<0.01) than all other treatments and control shrimp (0 mg L-1) (Fig. 1a,b).
Similarly, ALT of treatment 8.1 mg L-1 TAN was significantly higher (p<0.01) than that of other treatments and control shrimp except at days 6 and 8 where it was not significantly (p>0.01) different with that of treatment 3.8 mg L-1 TAN (Fig. 1). The concentration of both AST and ALT were not significantly different (p>0.01) among shrimp exposed to 3.8, 1.1 and 0 mg L-1 (control) TAN.
Haemolymph TP of treatment 8.1 mg L-1 TAN was significantly lower (p<0.01) than that of other treatments and control shrimp except at days 6 and 8 where TP of 8.1 mg L-1 TAN was not significantly different (p>0.01) with that of treatment 3.8 mg L-1 TAN (Fig. 2). Total protein in shrimp exposed to 1.1 mg L-1 ammonia was also not significantly different (p>0.01) from that of control during the experiment except at day 6. However, shrimp exposed to 3.8 mg L-1 ammonia showed significantly lower protein than that of control except at day 8.
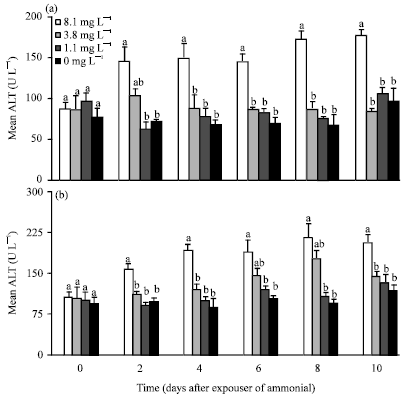 | |
| Fig. 1(a-b): | (a) Haemolymph ALT (U L-1) and (b) AST (U L-1) of P. monodon exposed to 8.1, 3.8, 1.1 and 0 mg L-1 TAN over 10 days. Each bar represents the mean value of 6 samples with standard error. Means with different letters indicate that they were significantly different (p<0.05) |
Effect of ammonia and WSSV infection on the mortality of P. monodon: Although, haemo-chemical parameters showed changes pre WSSV infection, Shrimp started to die only after 48 h post WSSV infection. Shrimp exposed to 8.1 mg L-1 TAN had significantly higher mortality rate than all other shrimp at 72 and 96 h post infection (Fig. 3a,b) and all died at 120 h.
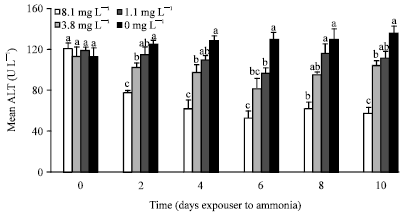 | |
| Fig. 2: | Total protein (g L-1) in haemolymph of P. monodon exposed to 8.1, 3.8, 1.1 and 0 mg L-1 TAN over 10 days. Each bar represents the mean value of 6 samples with standard error. Means with different letters are significantly different (p<0.05) |
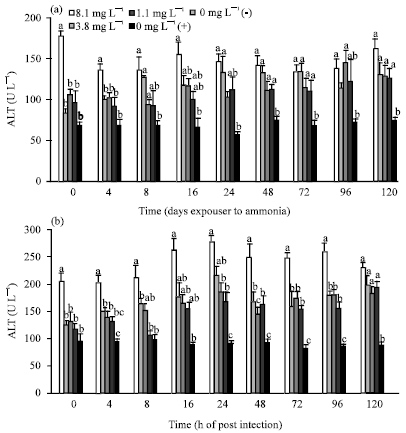 | |
| Fig. 3(a-b): | Haemolymph (a) ALT (U L-1) and (b) AST (U L-1) of Penaeus monodon post WSSV infection after 10 days exposure to 8.1, 3.8, 1.1 and 0 mg L-1 TAN. Each bar represents the mean value of 6 samples with standard error. Means with different letters indicate that they were significantly different (p<0.05) |
Shrimp exposed to 1.1 mg L-1 ammonia level and positive control shrimp (0 mg L-1) had significantly higher mortality rate than other treatments at 120 h (5th day) post infection. No shrimp mortality was recorded in the negative control (0 mg L-1 (-)) during the experiment.
Changes of haemo-chemical parameters during post WSSV infection: All seven haemo-chemical parameters examined were significantly different between shrimp sampled at pre and post WSSV infection (Table 1). Significant higher (p<0.01) concentration of ALT, AST, glucose and K+ and significant lower concentrations of total protein, Na+ and Cl¯ were observed in shrimp sampled after WSSV infection compared to shrimp sampled before infection (Table 1).
Shrimp exposed to 8.1 mg L-1 TAN had significantly higher ALT and AST concentrations than the negative control shrimp (0 mg L-1 (-)) at all sampling times (Fig. 3). Levels of ALT and AST were not significantly different among treatments 3.8, 1.1 and 0 (positive control) mg L-1 TAN at any sampling time. Level of ALT of treatment 8.1 mg L-1 TAN was not significantly different (p>0.01) with treatments 3.8, 1.1 and 0 mg L-1 TAN except at 4 h post infection. ALT in positive control shrimp was significantly higher (p<0.01) than that of negative control at 48, 72 and 120 h post infection. AST in positive control shrimp was also significantly higher (p<0.01) than negative control at 24, 72, 96 and 120 h post infection. At 4 and 120 h post infection shrimp in all treatments and positive control were not significantly different (p>0.01).
Haemolymph glucose level in shrimp exposed to 8.1 mg L-1 was significantly higher (p<0.01) than other treatments and controls except at 24 h post infection (Fig. 4a,b).
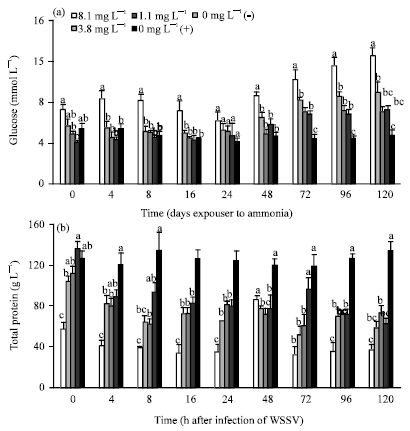 | |
| Fig. 4(a-b): | Haemolymph (a) glucose (mmol L-1) and (b) total protein (g L-1) of WSSV infected P. monodon after 10 days exposure to 8.1, 3.8, 1.1 and 0 mg L-1 TAN. Each bar represents the mean value of 6 samples with standard error. Means with different letters indicate that they were significantly different (p<0.05) |
| Table 1: | Biochemical parameters in shrimp pre WSSV and post WSSV infections (Means±Standard Error of Mean) |
 | |
| Values with different letters shows significant difference values at p = 0.01, student’s t-test | |
Glucose in shrimp exposed to 3.8 mg L-1 TAN was significantly higher (p<0.01) than negative control shrimp from 72 h post infection. Shrimp exposed to 1.1 and 0 mg L-1 TAN showed significantly higher glucose level than negative control at 72 and 96 h post infection.
Total protein in shrimp exposed to 8.1, 3.8 and 1.1 mg L-1 TAN was significantly lower (p<0.01) than the negative control (0 mg L-1 (-)) from 4 h post infection onwards (Fig. 4). Total protein in positive control shrimp was significantly lower (p<0.01) than that of negative control from 16 h post infection onwards except at 72 h. Total protein of treatments 3.8, 1.1 and 0 mg L-1 TAN were not significantly different (p>0.01) at all sampling times.
Although, the electrolytes (Na+, K+, Cl¯) were significantly different between at pre infection and post infection, they were not significantly different (p>0.01) among different ammonia treatments either at pre infection or post infection.
DISCUSSION
The results of the current study demonstrates that exposure of shrimp to different levels of total ammonia (at 8.1, 3.8, 1.1 mg L-1 TAN) significantly changed the haemo-chemical parameters without causing any shrimp mortality before WSSV infection and interestingly after WSSV infection there were more haemo-chemical changes with shrimp mortalities.
Although, the shrimp were exposed for 10 days to TAN (pre WSSV infection) shrimp mortality was observed only at 48 h post WSSV infection. Previous reports had indicated that shrimp were stressed without dying when they were exposed to maximum levels of TAN ranging from 54.6 for 24 h to 20 mg L-1 for 10 days (Jiang et al., 2004; Mugnier and Justou, 2004; Chen and Lin, 1992; Chen and Kou, 1991). However, shrimp mortality was reported when they were exposed to TAN ranging from 100 mg L-1 for 14 h to 20 mg L-1 for 20 days (Chen and Lin, 1992; Chen and Kou, 1991). Hence, for the present study shrimp were exposed to a maximum of 8.1 mg L-1 TAN for 15 days to induce stress without causing mortality. In the present study, it was interesting to note that shrimp that had undergone 10 days exposure to TAN, showed earlier mortality at 48 h post WSSV infection as compared to the non stressed shrimp which died after 3 days (Jiang et al., 2004), has also reported that shrimp generally die at 4 days after WSSV infection. The early mortality in the present study was most likely due to combined stress of ammonia and WSSV infection.
The present study showed that there was significant elevations in ALT and AST activities in haemolymph of shrimp subjected to 8.1 mg L-1 TAN. Pan et al. (2003) also observed similar results associated with the increased ammonia stress up to 20 mg L-1 TAN for 72 h. The increase concentration of ALT and AST can be found when body is subjected to any stress (Tunez et al., 2005). Since, ALT and AST did not show significant differences in the shrimp exposed to 3.8 and 1.1 mg L-1 TAN, it is believed that low level of TAN does not damage the tissues. Increased activities of ALT and AST in shrimp exposed to a high concentration of TAN (8.1 mg L-1) might have caused tissue damage. Tissue damages such as disruption and a subsequent collapse of pilaster cells, epithelial necrosis and hyperplasia of gill lamellae have been reported in estuarine crabs exposed to acute ammonia (De Freitas Rebelo et al., 2000).
In addition to elevation of haemolymph ALT and AST after exposed for 10 days to 8.1 mg L-1 TAN, the present study also showed that WSSV infection further increased ALT and AST which were significantly different to prior infection levels. Despite the absence of of a significantly increase levels of ALT and AST in shrimp exposed to lower TAN (3.8, 1.1 mg L-1) during the pre WSSV infection, in the post infection levels of ALT and AST were triggered to significantly higher levels. The increased ALT and AST levels by WSSV infection in the present study are similar to the findings of Mohankumar and Ramasamy (2006). They suggested that elevated levels of AST and ALT might have originated from extensive tissue damage and the amount of AST and ALT released was directly correlated with the number of cells affected by WSSV infection.
Pan et al. (2003) indicated that AST and ALT are enzymes involved in the transfer of amino groups and are mainly used as general indicators of vertebrate liver function. The crustacean hepatopancreas is assumed to be homologous to the mammalian liver and pancreas and is responsible for major metabolic events. Any stress or toxicity which causes damage to the hepatopancreas may result in high ALT and AST concentrations in crustaceans. Wang et al. (1997) showed degenerated cells with characteristics hypertrophied nuclei in WSSV-infected mesodermal and ectodermal tissues of moribund shrimp. From our study, we may suggest that the tissue damage caused by ammonia stress might have been further increased by an additional stress of WSSV infection; thereby causing a significant increase of AST and ALT levels resulting in early shrimp mortality.
The results on total protein analyses showed that the higher ammonia stress reduced protein levels in the haemolymph. Shrimp exposed to 8.1 mg L-1 TAN had significantly lower total protein in haemolymph than the control (0 mg L-1) and shrimp exposed to lower concentrations of ammonia (3.8, 1.1 mg L-1 TAN). Mugnier et al. (2008) described similar results on reduced total protein concentration observed in ammonia treated L. stylirostris. Chen et al. (1993) also reported the decrease of total protein when P. chinensis was exposed to as high as 10 mg L-1 of ammonia for 4 h. The reduced total protein concentration in the haemolymph in response to high ammonia may be due to catabolism of haemocyanin and total protein to free amino acids (Chen et al., 1994) to adjust the osmoregulation (Chen et al., 1993). It may be otherwise due to the proteolysis as shown by an increase in free amino acids or as a result of increased permeability to water (Mugnier and Justou, 2004).
Reduced total shrimp haemolymph protein can occur as a result of salinity and repeated handling stress (Mercier et al., 2006) and in destalked females extirpated males (Perazzolo et al., 2002). The above causes of decreased protein levels can be ruled out in the present study as the shrimp were maintained under standard conditions in all the experimental tanks. Thus, the decreased level of total protein in haemolymph during the pre infection period was due to the stress caused by ammonia. WSSV infection further reduced the total protein as it was significantly lower in positive control than in negative control. Jiang et al. (2004) also reported similar result that plasma protein was reduced in shrimp infected with WSSV after being stressed by exposing to 5 mg L-1 unionized ammonia. However, their findings on shrimp challenged with WSSV without undergoing exposure to ammonia showed an increased total protein for first 6 h of post infection followed by a reduction. In the present study, further reduction of total protein in the post WSSV infection may be due to catabolism of plasma protein by proteases reported to be encoded by WSSV (Yoganandhan et al., 2003).
During the exposure to different levels of ammonia for 10 days, there was no significant difference in haemolymph glucose in P. monodon. Mugnier et al. (2008) also reported that haemolymph glucose concentration in L. stylirostris exposed to 44-71.5 mg L-1 for 24 h was not significantly different with that of control shrimp. However, previous studies (Mugnier and Justou, 2004) showed that the hyperglycemia was observed in the shrimp treated with ammonia. Hyperglycemia is a consequence of stress and an indicator of a wide range of perturbations (Mugnier et al., 2008). The stress caused by ammonia in the present study might not be enough to induce hyperglycemia. However, glucose concentration significantly increased after the shrimp exposed to ammonia were infected with WSSV. Yoganandhan et al. (2003) reported increased level of haemolymph glucose in WSSV-infected P. indicus and indicated that it may be due to the transport of glucose and carbohydrate from hepatopancreas and muscle to haemolymph. Increase of blood glucose level in P. monodon was also observed to be caused by other stressors such as depletion of dissolved oxygen, increased dissolved carbon dioxide levels, or perturbation which may be encountered in a pond (Hall and van Ham, 1998). In addition, Mohankumar and Ramasamy (2006) indicated that reduced feeding in shrimp infected with WSSV is due to decreased oxidation of glucose. Hence, the hyperglycemia and reduced oxidation of glucose may result in high haemolymph glucose in WSSV-infected shrimp.
There was significant difference in electrolyte levels between pre WSSV and post WSSV infection shrimp, although there was no significant difference of electrolytes among the treatments either at pre infection and post infection periods. When an animal is exposed to stressful condition, the changes in electrolytes level were observed in the body fluid by many studies (Uboh et al., 2008; Maryam et al., 2007; Anwar and Zia-ur-Rahman, 2002). However, Chen and Chen (1996) reported that there was a significant decrease of haemolymph electrolytes under ammonia stress at concentration of 1.47 mmol L-1. The stress exerted by the ammonia concentration (0.82 mg L-1 un-ionized ammonia) in the present study was not sufficient to change the haemolymph electrolytes. However, the WSSV infection caused significantly lower Na+ and Cl¯ and higher K+ concentrations. A recent study by Chang et al. (2007) showed that ammonia stress can be reduced by dopamine injection which result in elevations of osmolality, Na+ and Cl¯ and reduction of K+ level in the haemolymph. This finding may be similar to our results. The elevation of electrolytes may be due to the change of permeability and osmolality of membranes of gills and other surface. Change of permeability and osmolality may be due to impaired Na+, K+-ATPase activity in haemolymph, hepatopancrease, gills and muscles of WSSV-infected shrimp (Mohankumar and Ramasamy, 2006).
CONCLUSION
Changes of haemo-chemical parameters during the first 10 days to ammonia exposure showed that as low as 3.8 mg L-1 TAN exposure for 2 days was sufficient to stress the shrimp as indicated by the significant increase of ALT and AST and the low total protein concentrations. Shrimp mortality was observed as early as 48 h post WSSV infection, after exposure to ammonia. Along with the increase levels of exposure to ammonia there was also a trend of increase stress on shrimp. Shrimp exposed to ammonia and subsequently infected with WSSV showed significantly greater stress compared to shrimp prior to the infection. And also it is crucial take control measures to reduce the nutrient level in the shrimp pond water as suggested by Venkatesan et al. (2006).
REFERENCES
- Chang, C.C., Z.R. Wu, C.S. Chen, C.M. Kuo and W. Cheng, 2007. Dopamine modulates the physiological response of the tiger shrimp Penaeus monodon. Aquaculture, 270: 333-342.
CrossRef - Chen, J.C. and C.T. Chen, 1996. Changes of osmotic and electrolyte concentrations in the haemolymph of Penaeus japonicas exposed to ambient ammonia. Comp. Biochem. Physiol. C, 114: 35-38.
CrossRef - Chen, J.C. and S.Y. Cheng, 1995. Hemolymph oxygen content, oxyhemocyanin, protein levels and ammonia excretion in the shrimp Penaeus monodon exposed to ambient nitrite. J. Comp. Physiol. B, 164: 530-535.
CrossRef - Chen, J.C. and Y.Z. Kou, 1991. Accumulation of ammonia in the hemolymph of Penaeus japonicas exposed to ambient ammonia. Dis. Aquat. Org., 11: 187-191.
Direct Link - Chen, J.C., S.Y. Cheng and C.T. Chen, 1994. Changes of haemocyanin, protein and free amino acid levels in the haemolymph of Penaeus japonicas exposed to ambient ammonia. Comp. Biochem. Physiol. A, 109: 339-347.
CrossRef - Chen, J.C., F.H. Nan, S.Y. Cheng and S.S. Sheen, 1993. Effects of ambient ammonia on ammonia-N and protein concentrations in hemolymph and ammonia-N excretion of Penaeus chinensis. Mar. Ecol. Prog. Ser., 98: 203-208.
Direct Link - Chiba, S., L. Aoki and T. Ogata, 2004. Response of the pandalid shrimp Pandalus latirostris to dissolved oxygen, salinity and turbidity. Fish. Sci., 70: 1174-1176.
CrossRef - Chim, L., R. Bouveret, P. Lemaire and J.L.M. Martin, 2003. Tolerance of the shrimp Litopenaeus stylirostris, Stimpson 1894, to environmental stress: Interindividual variability and selection potential for stress-resistant individuals. Aquacult. Res., 34: 629-632.
CrossRef - Fouzi, M., M. Shariff, A.R. Omar, F.M. Yusoff and S.W. Tan, 2010. TaqMan real-time PCR assay for relative quantification of white spot syndrome virus infection in Penaeus monodon Fabricius exposed to ammonia. J. Fish Dis., 33: 931-938.
CrossRef - Hall, M.R. and E.H. van Ham, 1998. The effects of different types of stress on blood glucose in the giant tiger prawn Penaeus monodon. J. World Aquacult. Soc., 29: 290-299.
CrossRefDirect Link - Jiang, G., R. Yu and M. Zhou, 2004. Modulatory effects of ammonia-N on the immune system of Penaeus japonicas to virulence of white spot syndrome virus. Aquaculture, 241: 61-75.
CrossRef - Joseph, A. and R. Philip, 2007. Acute salinity stress alters the haemolymph metabolic profile of Penaeus monodon and reduces immunocompetence to white spot syndrome virus infection. Aquaculture, 272: 87-97.
CrossRef - Kautsky, N., P. Ronnback, M. Tedengren and M. Troell, 2000. Ecosystem perspectives on management of disease in shrimp pond farming. Aquaculture, 191: 145-161.
CrossRef - Liu, B., Z. Yu, X. Song, Y. Guan, X. Jiang and J. He, 2006. The effect of acute salinity change on White Spot Syndrome (WSS) outbreaks in Fenneropenaeus chinensis. Aquaculture, 253: 163-170.
CrossRefDirect Link - Mohankumar, K. and P. Ramasamy, 2006. Activities of membrane bound phosphatases, transaminases and mitochondrial enzymes in white spot syndrome virus infected tissues of Fenneropenaeus indicus. Viral Res., 118: 130-135.
CrossRefDirect Link - Mugnier, C., E. Zipper, C. Goarant and H. Lemonnier, 2008. Combined effect of exposure to ammonia and hypoxia on the blue shrimp Litopenaeus stylirostris survival and physiological response in relation to molt stage. Aquaculture, 274: 398-407.
CrossRefDirect Link - Mugnier, C. and C. Justou, 2004. Combined effect of external ammonia and molt stage on the blue shrimp Litopenaeus stylirostris physiological response. J. Exp. Mar. Biol. Ecol., 309: 35-46.
CrossRef - Mugnier, C., H. Lemonnier and A. Legrand, 2006. Physiological response of the blue shrimp Litopenaeus stylirostris to short term confinement on a pond bottom. Aquaculture, 253: 703-711.
CrossRefDirect Link - Pan, C.H., Y.H. Chien and B. Hunter, 2003. The resistance to ammonia stress of Penaeus monodon Fabricius juvenile fed diets supplemented with astaxanthin. J. Exp. Mar. Biol. Ecol., 297: 107-118.
CrossRefDirect Link - Parsons, R.T., M. Yoshiaki and G.M. Lalli, 1984. A Manual of Chemical and Biological Methods for Seawater Analysis. 1st Edn., Pergamon Press, Oxford, UK., ISBN: 9780080302874, Pages: 173.
Direct Link - Pascual, C., A. Sanchez, A. Sanchez, F. Vargas-Albores, G. LeMoullac and C. Rosas, 2003. Haemolymph metabolic variables and immune response in Litopenaeus setiferus adult males: The effect of an extreme temperature. Aquaculture, 218: 637-650.
CrossRefDirect Link - Perazzolo, L.M., R. Gargioni, P. Ogliari and M.A.A. Barracco, 2002. Evaluation of some hemato-immunological parameters in the shrimp Farfantepenaeus paulensis submitted to environmental and physiological stress. Aquaculture, 214: 19-33.
CrossRef - Rahman, M.M., C.M.E. Bonilla, M. Corteel, J.J. Dantas-Lima and M. Wille et al., 2006. Effect of high water temperature (33 °C) on the clinical and virological outcome of experimental infections with White Spot Syndrome Virus (WSSV) in Specific Pathogen-Free (SPF) Litopenaeus vannamei. Aquaculture, 261: 842-849.
CrossRefDirect Link - Rodriguez-Ramos, T., G. Espinosa, J. Hernandez-Lopez, T. Gollas-Galvan and J. Marrero et al., 2008. Effects of Echerichia coli lipopolysaccharides and dissolved ammonia on immune response in southern white shrimp Litopenaeus schmitti. Aquaculture, 274: 118-125.
CrossRefDirect Link - De Freitas Rebelo, M., E.M. Rodriguez, E.A. Santos and M. Ansaldo, 2000. Histopathological changes in gills of the estuarine crab Chasmagnathus granulata (Crustacea-Decapoda) following acute exposure to ammonia. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 125: 157-164.
PubMed - Tunez, I., M.C. Munoz, M.A. Villavivicencio, F.J. Medina and E.P.de Prado et al., 2005. Hepato- and neurotoxicity induced by thioacetamide: Protective effects of melatonin and dimethylsulfoxide. Pharmacol. Res., 52: 223-228.
PubMed - Wang, C.S., K.F.J. Tang, G.H. Kou and S.N. Chen, 1997. Light and electron microscopic evidence of white spot disease in the giant tiger shrimp, Penaeus monodon (Fabricius) and the kuruma shrimp, Penaeus japonicas (Bate), cultured in Taiwan. J. Fish Dis., 20: 323-331.
CrossRef - Sihag, R.C. and P. Sharma, 2012. Probiotics: The new ecofriendly alternative measures of disease control for sustainable aquaculture. J. Fish. Aquat. Sci., 7: 72-103.
CrossRefDirect Link - Poornima, R., M.K. Sahu, K. Sivakumar and V. Pushpavalli, 2008. Optimization of α-amylase production by actinomycete strain AE-19 isolated from shrimp pond. Trends Applied Sci. Res., 3: 45-52.
CrossRefDirect Link - Rao, M.S., B. Rajitha, E. Pavitra and N. Anjaneyulu, 2008. Changes of copper and protein profiles in hepatopancreas and hemolymph tissues during different molt stages of white shrimp, Litopenaeus vannamei (Boone, 1931). Biotechnology, 7: 153-156.
CrossRefDirect Link - Venkatesan, R., K.P.K. Vasagam and T. Balasubramanian, 2006. Culture of marine microalgae in shrimp farm discharge water: A sustainable approach to reduce the cost production and recovery of nutrients. J. Fish. Aquatic Sci., 1: 262-269.
CrossRefDirect Link - Arimoro, F.O., C.M.A. Iwegbue and O. Osiobe, 2008. Effects of industrial waste water on the physical and chemical characteristics of a tropical coastal river. Res. J. Environ. Sci., 2: 209-220.
CrossRefDirect Link - Abbas, H.H., 2006. Acute toxicity of ammonia to common carp fingerlings (Cyprinus carpio) at different pH levels. Pak. J. Biol. Sci., 9: 2215-2221.
CrossRefDirect Link - Uboh, F.E., M.I. Akpanabiatu, I.J. Atangwho and Y. Alozie, 2008. Exposure to gasoline vapours: A potential risk factor for atherosclerosis in male and female rats. J. Pharmacol. Toxicol., 3: 600-609.
CrossRefDirect Link - Anwar, H. and Zia-ur-Rahman, 2002. Effect of Ascaridia galli infestation on electrolytes and vitamins in chickens. J. Biol. Sci., 2: 650-651.
CrossRefDirect Link - Tehranipour, M., H.R. Ali, B.R. Morteza, P. Kazem and R. Amin, 2007. Determination of the cerebrospinal fluid electrolytes alteration in the developing rats born from diabetic mothers. J. Biol. Sci., 7: 969-972.
CrossRefDirect Link