
Appiah Catherine
Laboratory of Plant Ecology and Ecological Genetics, Graduate School of Environmental Science, Hokkaido University, N 10 W 5 060-0810, Sapporo, Japan
Shiro Tsuyuzaki
Laboratory of Plant Ecology and Ecological Genetics, Graduate School of Environmental Science, Hokkaido University, N 10 W 5 060-0810, Sapporo, Japan
Djietror Jonathan Castel
Laboratory of Plant Ecology and Ecological Genetics, Graduate School of Environmental Science, Hokkaido University, N 10 W 5 060-0810, Sapporo, Japan
Research Journal of Seed Science
Year: 2012 | Volume: 5 | Issue: 4 | Page No.: 126-135







ABSTRACT
Gold mining occurs in four administrative regions in Ghana with tropical forest cover. Establishment of pioneer vegetation immediately after the mining process is one of the most important rehabilitation practices leading to land reclamation and eventual habitat restoration. A major environmental challenge is that post-mining vegetation sites are prone to wild forest fires that dry up the topsoil and produce a layer of ash on the soil surface. We experimentally tested the effect of post-fire ash and soil moisture on seed germination using the Black-eyed pea (Vigna unguiculata L. Walp) and Egusi (Citrullus colocynthis L.). The study focused on the potential for seed germination in ash under high and low moisture conditions within a greenhouse environment. The viable seeds of the two species were planted at depths; 0.0, 0.5 and 1.0 cm. V. unguiculata exhibited the highest germination percentage at 0.0 cm in both moisture treatments, but no germination occurred at 0.5 and 1.0 cm depth, showing that germination was considerably inhibited by the ash depth. C. colocynthis showed a comparatively longer dormancy period but with superior germination percentage at all 3 depths under wet conditions. Due to the lower seed germination rate for V. unguiculata under ash, the species might not be easily adapted to ash-covered soils. However, due to significantly higher germination rates under similar conditions, C. colocynthis could potentially be adapted for post-mine vegetation restoration after the occurrence of wild forest fires.
PDF Abstract XML References Citation
Received: April 25, 2012;
Accepted: July 31, 2012;
Published: January 29, 2013
How to cite this article
Appiah Catherine, Shiro Tsuyuzaki and Djietror Jonathan Castel, 2012. Post Fire Restoration Study of Ash and Moisture Variation Effects on Seed Germination of Citrullus colocynthis and Vigna unguiculata . Research Journal of Seed Science, 5: 126-135.
URL: https://scialert.net/abstract/?doi=rjss.2012.126.135
URL: https://scialert.net/abstract/?doi=rjss.2012.126.135
INTRODUCTION
Post mining land revegetation success in fire-prone ecosystems is influenced by the ecology of the fire-seed-germination interactions during the pioneer vegetation growth. Direct seeding with available native plants is crucial for the enhancement of natural revegetation processes (Tordoff et al., 2000). Two native species are here investigated for their suitability for revegetation of mine lands in Ghana; (a) wild gourd, or bitter apple, egusi, ose-oji, ogiri-isi and avrouda-West Africa, Citrullus colocynthis, (Ogbonna and Obi, 2000), is a species of the gourd family Cucurbitaceae (Schafferman et al., 1998); (b). V. unguiculata a high N-fixing legume (Sangakkara and Bieler, 1988). C. colocynthis is a non-hardy, herbaceous plant that is native to tropical Asia and Africa. The stems are angular and rough; the leaves rough, 5 to 10 cm in length, with 3 to 7 deep lobes and produce solitary pale yellow blooms. Depending on soil and microclimate, each plant produces 20 to 30 fruits, with maximum 10 cm in diameter and green with wavy yellow stripes. The fruits totally transform into all yellow when dry at the dispersal maturity. Seeds of C. colocynthis are small with 2.54 cm or less in length. C. colocynthis can tolerate annual temperature of 14.8 to 27. 8°C (mean of 10 cases = 22.5) and thrives in pH of 5.0 to 7.8 (mean of 8 cases = 6.8) soil acidity. This species was considered for restoration due to several desirable traits; high drought tolerance, ability to thrive in sandy loam, sub-desert soils and along sandy sea coasts where the mean annual temperature ranges from 23-27°C and annual rainfall ranges between 250-370 mm (Ehlers and Hall, 1997). Black-eyed pea, V. unguiculata (L) Walp is a native legume that was chosen on the basis of the stress tolerance, heat tolerance, nitrogen fixing capacity and adaptability to the tropical climate. Sown seeds of V. unguiculata are very susceptible to frost; exhibit optimum germination only in warm seasons with 25-35°C optimum temperature and at specific planting depths per variety (Mahdi et al., 1998; Aikins and Afuakwa, 2006; CRI, 2006). The ecological impact of vegetation removal prior to mining activities in forest regions of Ghana underscores the need to establish pioneer vegetation immediately after the end of mining operations (Torbert and Burger, 2002). Planting of native plant species on mined sites is believed to form an important component of post-mining ecosystem restoration (Bradshaw, 1997; Djietror et al., 2011b) without which the mined surface would be exposed to severe erosion and further degradation. It has therefore, become imperative to establish early fast growing pioneer vegetation that could control soil erosion, maintain the soil fertility and retain the edaphic constitution of the post-mining land sites to near-original levels. Perennial wild fires consume existing vegetation and destroy the pioneer vegetation (Cochrane, 2001; FORIG, 2003) that shelters the topsoil. The conditions of the distinct dry harmattan season occur from late November to early March. High temperature (approximate mean: 40°C) dries out the soil and the pioneer vegetation and this desiccation facilitates bush fires. The post-fire ash accumulation on such disturbed habitats is believed to have a significant impact on initial seed germination (Reyes and Casal, 2004). In Ghana, post-mining land revegetation has largely involved fast growing exotic plant species (Dorgbetor et al., 2012). There is the need for plant species that could produce rapid germination and fast-grown seedlings that will establish quicker revegetation of bare mined sites. Current plant species in use are not well-adapted and are unable to survive and reproduce when planted on the degraded post-mining land sites. Some of the introduced plant species which are adaptable to the degraded post-mining soil conditions, have conversely become persistent, invasive (Djietror et al., 2011a) and quickly replace the indigenous plant communities in the adjacent lands (Djietror et al., 2011b). The impact of accumulated ash from wild fires has been known to have inhibitory or neutral impacts on the germination of many exotic plant seeds (Franzese and Ghermandi, 2011), however little is known about the impact of the post-fire ash load on the seed germination rate of the largely unutilized and ecologically adaptable native African plant species. The aim was therefore, to determine the suitability of native plant species as choice species for post-mining land rehabilitation within fire prone ecosystems. Special attention was given to depth of ash on seed emergence.
MATERIALS AND METHODS
Study site and seed collection: Sourcing of seeds and conducting of greenhouse experiments were done between April 2011 and January, 2012. Seeds of two native species the Black-eyed Pea; V. unguiculata L. Walp and Egusi; C. colocynthis L. were collected from 20 different farm sites around the gold mining areas of AngloGold Ashanti (Obuasi), Newmont Ghana Gold Limited (Kenyasi) and Golden Star Resources (Prestea-Bogoso) in the Ashanti (6°41''15.24''N 1°37'09.194''W), the Brong-Ahafo (7°19'58.45''N, 2°19'57.84''W) and the Western (5°34'35.15''N 2°00'40.54''W) regional areas, respectively (Fig. 1).
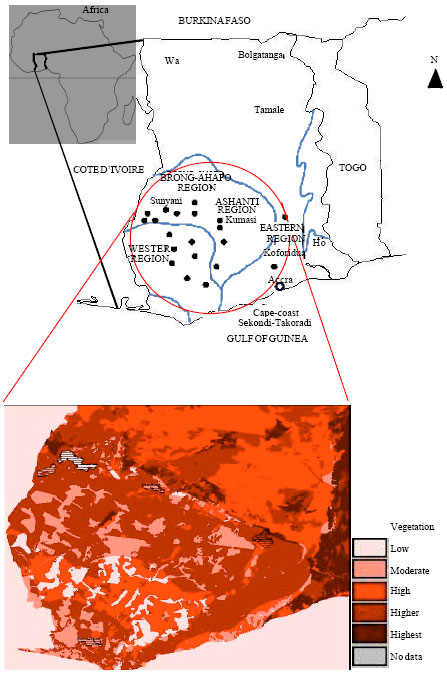 | |
| Fig. 1: | Map of Ghana depicting the tropical forest areas (bound by blue line) and main four administrative regions where gold mining occurred. The seed source sites are marked with black dots. The projected red map shows fire incidence areas (FORIG, 2003) based on vegetation zones and other land cover characteristics |
The main sources of the seeds were certified commercial seed distributors and vegetable crop farmers.
Seed viability test: To select viable seeds from the sourced seed mass, the seeds were immersed in (NH4)2SO4 solution of specific gravity 1.08. The selected viable seeds were disinfected at 10°C for 24 h in 0.1% Tichrid C solution (0.5 g/500 mL of distilled water) and were rinsed (Van Hezewij and Verkleij, 1996). The seeds of the two species were soaked separately in distilled water for 20 min, sieved and dried at a room temperature for 30 min.
Experimental design and laboratory experiment: The soil samples from the sites in Ghana were collected and analyzed for moisture content and the buffer index using the Spurway extract method. This involved a weak acetic acid extraction of the soil media. Soil samples were obtained from the farm sites where native legumes were traditionally cultivated. Meteorological data photoperiod range (10 to 12 h), precipitation-1100 mm year-1), maximum daily temperature (26.8°C) and minimum daily temperature, (22.2°C) of the seed source sites were obtained from the Ghana Meteorological Service. Both the soil and environmental data served as a basis for controlling the edaphic factors and internal temperature for the greenhouse seed germination experiment at the Field Science Research Laboratory at Hokkaido University, Japan. Seeds quality and purity were evaluated in three pre-experimental trial tests in 0.9x1.5 m glass dishes to determine the viability. The seeds were tested in an incubator above three filter papers (Whatman 181) moistened with 4.5 mL of deionized water, with 4 replicates of 50 seeds per test. The temperature within the incubator was monitored and logged every 30 min using a data logging system (TempScan/1000A, IO Tech, Ohio, USA). The viability rate percentage was 98.6% per 300 seeds.
Experimental design: The two native vegetable seeds were tested in 3 treatments (seeding depths; 0.0, 0.5 and 1.0 cm, Table 1), 2 sub-treatments (seed media; wet and dry) and 4 replications. One hundred seeds of each species were sown randomly in 25x15 cm opaque trays filled with commercial wood ash. The trays were randomly arranged in 4 blocks (Randomized Complete Block Design) that were set on two 3x9 m tables in the greenhouse. The green house was equipped with a sprinkler system for irrigation. To create wet and dry conditions, one table was set in a highly irrigated (wet sub-treatment) of the greenhouse with >50 mm of water per week. The second table was set in the mildly irrigated (dry sub-treatment) with <20 mm of water per week. The mean internal greenhouse air temperature was 26.5°C and mean relative humidity was 50%.
Germination analysis: The number of newly germinated seeds was evaluated at the emergence and initial downward bending of the root system. The parameters for normal seed germination set by the International Seed Testing Association (ISTA, 2005) were used as the germination criteria. The germinated seeds were counted and recorded every 24 h over a five week period. The measurement of the Mean Germination Rate (MGR) was based on the methodology of Silva and Nakagawa (1995):
| Where: | ||
| SGNI | = | Seed number germinated in the interval |
| TI | = | Interval time |
| TGSN | = | Total germinated seed number |
| Table 1: | The different seeding depths, seeding media and wood ash treatment involving vermiculite |
 | |
Statistical analysis: A two-way Analysis of Variance (ANOVA) was done considering the species with depth treatments and moisture treatments as factors. The data obtained from the germination experiments were statistically analyzed for mean separation using IBM SPSS Statistical software (Version 18.0). The species and moisture and depth treatments were set as independent variables. A set of sub-samples from all the treatments were obtained to determine the correlation between the ash moisture content and seeding depth. The experimental period was extended for 5 weeks in order to allow optimum time covering the variable dormancy period of seeds from different sites. At the end of the germination period, each block was sampled for the mean number of germinated seeds in each tray. This was calculated as a percentage of the total seed number (1200 seeds per seed treatment).
RESULTS
Germination rate in both wet and dry treatment for V. unguiculata and C. colocynthis was significantly (p<0.0001) different between the treated burial depths of the seeds in wood ash. In the control treatment (depth = 0.0 cm), the mean germination percentage (32.2%) for both wet and dry treatments, was significantly higher than the germination rate at seeding depth of 0.5 cm (7.83%) and at 1.0 cm (0.4%). The wet treatment resulted in significantly different (p<0.0001) percentages between the three depth treatments. The highest percentage was seeding treatment without ash and the lowest was in 1.0 cm ash depth (Fig. 2). On average, the germination of the seeds began 3-5 days earlier in the control treatment than the ash treatments. Between the dry and wet ash treatments, the wet treatment (22.9%) was significantly (p<0.0001) higher than the germination percentage within the dry ash treatment (4.04%). The rate of germination was faster among seeds of V. unguiculata than for C. colocynthis. The weekly mean germination percentages presented in Fig. 3 show 100 % germination for V. unguiculata at the end of the second week of the germination period. A variation in germination rates with respect to ash depth treatments occurred among seeds of C. colocynthis.
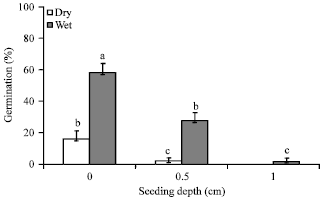 | |
| Fig. 2: | Variation in the mean germination percentages at different depths of wood ash investigated under both dry and wet conditions. Standard error is indicated on the bars, different letters depict the existence of significant difference (p<0.01, Tukey test) between treatments and seedling depth |
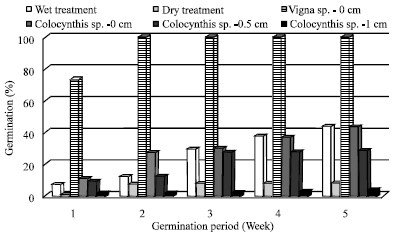 | |
| Fig. 3: | The weekly mean germination percentages of Vigna unguiculata and Citrullus colocynthis at different depths of wood ash experimented under dry and wet ash conditions |
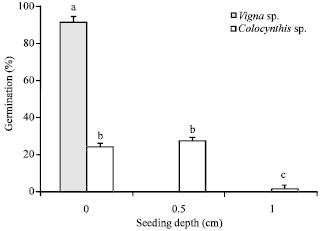 | |
| Fig. 4: | Germination rate of V. unguiculata and C. colocynthis expressed as percentage at the different seed burial depths under wet ash condition, Standard error is indicated on the bars, different letters depict the existence of significant difference (p<0.01, Tukey test, n = 1200) between ash depths and species |
Rate of germination consistently increased between 1st and 5th Week at 0 cm and between 2nd and 5th Week at 0.5 cm. There was no marked variation in the germination rate at 1.0 cm during the germination period. The germination rate under the dry treatment did not show clear increase while the wet treatment resulted in consistent increase between 1st and 5th Week (Fig. 3).
Within the wet treatment (Fig. 4), germination rate of C. colocynthis did not significantly vary between 0.5 cm (27.3%) and the control 0.0 cm (24.3%) though these rates were significantly (p<0.001) higher than the depth of ash at 1.0 cm (1.5%). The results depict a comparatively lower germination rate in the dry treatment for seeds of C. colocynthis (Fig. 5). In the dry treatment, the highest germination rate occurred without ash (4.4%). However, this percentage was not significantly different (p>0.201) from the germination rate when the seeds were sown at the depth of 0.5 cm (1.6%) and 1.0 cm (0.1%), probably due to low seed germination.
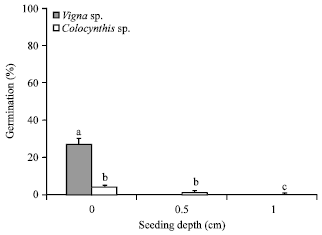 | |
| Fig. 5: | Germination rate of V. unguiculata and C. colocynthis at the different seed burial depths under dry ash condition, Standard error is indicated on the bars, different letters depict the existence of significant difference (p<0.01, Tukey test, n = 1200) between depth and species |
V. unguiculata exhibited the highest germination rate at 0.0 cm depth (without ash) in the wet treatment (Fig. 4). Germination rate was 91.2% at 0 cm depth, while no germination occurred at 0.5 and 1.0 cm depth in the wet ash treatment. Germination rate of V. unguiculata (27.2%) in the dry treatment was significantly (p<0.0001) higher in the control (Fig. 5). Similar to the germination trend recorded in the wet ash treatment (Fig. 4), no germination of V. unguiculata occurred at the seeding depth of 0.5 and 1.0 cm in the dry ash treatment (Fig. 5). When seeded at 0 cm depth in both dry and wet treatment conditions, V. unguiculata showed a significantly (p<0.0001) higher germination rate (20.1%) than did C. colocynthis without ash treatment. Conversely, C. colocynthis germinated in ash treatment, though at a reduced rate (8.5%) and the percentage germination was higher at shallow depth (<0.5 cm).
DISCUSSION
The germination rate of V. unguiculata was generally higher than C. colocynthis. Without ash treatment, V. unguiculata showed comparatively higher seed germination rate, but depicted inferior germination rate at the two seeding depths under wet ash conditions. Seeds of both species could not germinate at higher seeding depth (>1.0 cm) in both wet and dry ash treatments.
Increased seed germination rates were reported in fire-prone habitats (Reyes and Casal, 2004). Increase in the post-fire seed germination, dependent on the specific species involved, could range from the impact of smoke (Reyes and Casal, 2004; Bankole et al., 2005; Susana et al., 2011), heat and or temperature changes (Covell et al., 1986; Craufurd et al., 1996) that affect the seed epicarp and pericarp layers due to the chemicals released during a fire and in the resulting ash that gets accumulated (Bankole and Joda, 2004; Lobato et al., 2009). However, the quantity of ash that persists on a site after a wild fire can considerably alter the rate of seed germination and the dynamics of the pioneer vegetation that emerges during the first stage of succession (Rivard and Woodard, 1989). In this particular study on the germination of African species, the germination rate of V. unguiculata was higher in the control treatments (without ash) than in the treatments with either wet or dry ash. However, ash could induce a stagnating effect on seed germination on soil within fire-prone habitats (Gonzalez-Rabanal and Casal, 1995; Baxter et al., 1994) immediately after the occurrence of forest fire. This trend suggests V. unguiculata might not easily germinate on revegetation sites that are prone to fire. On the positive aspects of this species, seed germination was very high without ash, especially under wet treatment conditions at very shallow seeding depth. With a very high N-fixing capacity (Kendle and Bradshaw, 1992), this native legume could be adapted to post-mining land sites that have not been exposed to wild fires. The potential for seed germination may actually be optimum during the major rainy season (May-June). V. unguiculata may be the best considered for rapid pioneer vegetation under planned irrigation conditions (Sangakkara, 1998; CRI, 2006) in controlled revegetation units (Bradshaw, 2000; Djietror et al., 2011b).
C. colocynthis showed the highest seed germination under the depth of 0.5 cm wet ash treatment. There is evidence that ash could facilitate the seed germination rate in some related legume species (Wolfson et al., 2005). In related studies it has been established that most species in fire-prone primary vegetation in tropical and subtropical areas exhibit increased seed germination rate. Thus, the effects of fire products on seed germination can be complementary when the ash thickness is thin (Wolfson et al., 2005; Susana et al., 2011).
Potentially, depending on the variations in topsoil moisture content during dry and wet periods, there could be variable survival rates among the native species within varying quantities of post-fire ash. Owing to the ability to germinate in fire run habitats, the native African legume species could be easily adapted for use as pioneer species in post-mining land rehabilitation that might culminate in the eventual full ecosystem restoration. Germination is higher at shallower seeding depth (Kirby, 1993; Karayel and Ozmerzi, 2008) and the germination rate tends to decrease as the seeding depth of the ash is increased (Campbell et al., 1991). In particular, C. colocynthis seeded in 0.5 cm deep showed a higher germination rate. This implies that while the ash inhibited seedling emergence at higher depths (>1.0 cm), it also had a positive effect on this species given that the species is characterized by long dormancy period. At seeding depth of 0 cm, the germination rate of C. colocynthis in wet ash was as high as it occurred at 0.5 cm. The seeding depth of 0.5 cm might be the optimum seeding depth for C. colocynthis in conditions of ash accumulation under rain-fed conditions. The optimum germination depth of this native species should be an important ecological parameter for revegetation in the fire-prone ecosystem that spans the gold-mining region of Ghana with a consistently widening degraded mine land area.
CONCLUSION
The utilization of plant materials for the amendment of bare post-mining landscapes and the revegetation of fire-prone mine land sites in Ghana, currently incorporate a limited use of native grasses and legumes. C. colocynthis showed a potential for germination in >0.5 cm depth of wood ash, even under conditions of reduced moisture content of the germination medium. The species could therefore, be adaptable for initial revegetation of ash covered post-fire land habitats including severely disturbed mined land sites. V. unguiculata is considered a highly suitable legume with a high capacity for seed germination under moisture conditions. The species however, has a weaker potential for germination in ash and therefore, may not be a suitable plant species for initial site revegetation (pioneer vegetation establishment) on fire prone sites with large quantity of ash deposit. V. unguiculata could be incorporated into existing vegetation to enhance species richness during the latter stages of mine land rehabilitation. It is important to advance this study by investigating the ecology, growth and survival dynamics of the native species within the pioneer vegetation and a climax community of species. Furthermore, future research should investigate other native species that might be adaptable to regenerate following fire disturbance on post mining sites. Additional research is needed to experimentally test the use of native plant materials and inorganic amendments to improve soil fertility for supporting the pioneer vegetation.
REFERENCES
- Aikins, S.H.M. and J.J. Afuakwa, 2006. Growth and dry matter yield responses of cowpea to different sowing depths. ARPN J. Agric. Biol. Sci., 3: 50-54.
Direct Link - Bankole, S.A., A. Osho, A.O. Joda and O.A. Enikuomehin, 2005. Effect of drying method on the quality and storability of 'Egusi' melon seeds (Colocynthis citrullus L.). Afr. J. Biotechnol., 4: 799-803.
CrossRefDirect Link - Baxter, B.J.M., J. Van Staden, J.E. Granger and N.A.C. Brown, 1994. Plant-derived smoke and smoke extracts stimulate seed germination of the fire-climax grass Themeda triandra. Environ. Exp. Bot., 34: 217-223.
CrossRef - Bradshaw, A., 1997. Restoration of mined lands-using natural processes. Ecol. Eng., 8: 255-269.
CrossRefDirect Link - Bradshaw, A., 2000. The use of natural processes in reclamation: Advantages and difficulties. Landscape Urban Plann., 51: 89-100.
CrossRef - Campbell, C.A., F. Selles, R.P. Zentner, J.G. McLeod and F.B. Dyck, 1991. Effect of seeding date, rate and depth on winter wheat grown on conventional fallow in S.W. Saskatchewan. Can. J. Plant Sci., 71: 51-61.
Direct Link - Cochrane, M.A., 2001. Synergistic interactions between habitat fragmentation and fire in evergreen tropical forests. Conserv. Biol., 15: 1515-1521.
CrossRef - Covell, S., R.H. Ellis, E.H. Roberts and R.J. Summerfield, 1986. The influence of temperature on seed germination rate in grain legumes. I. A comparison of chickpea, lentil, soyabean and cowpea at constant temperatures. J. Exp. Bot., 37: 705-715.
CrossRefDirect Link - Craufurd, Q., H. Ellis, J. Summerfield and L. Menin, 1996. Development in cowpea (Vigna unguiculata). I. The influence of temperature on seed germination and seedling emergence. Exp. Agric., 32: 1-12.
CrossRefDirect Link - Djietror, J.C., M. Ohara, S. Tsuyuzaki and C. Appiah, 2011. On the path of invasion: Disturbance promotes the growth vigor among siam weeds in a mine land ecosystem. Res. J. For., 5: 144-153.
CrossRefDirect Link - Djietror, J.C., M. Ohara and C. Appiah, 2011. Predicting the establishment and spread of siam weed in Australia: A test of abiotic cues on seed dormancy and germination. Res. J. For., 5: 115-127.
CrossRefDirect Link - Dorgbetor, W.H.K., G.N.N. Dowuona, S.K.A. Danso, J.K. Amatekpor, A.O. Ogunkunle and E. Boateng, 2012. Evaluation of quality of some rehabilitated mined soils within the AngloGold-Ashanti concession in Ghana. Int. J. Geosci., 3: 50-61.
CrossRefDirect Link - Ehlers, J.D. and A.E. Hall, 1997. Cowpea (Vigna unguiculata L. Walp.). Field Crops Res., 53: 187-204.
CrossRefDirect Link - Gonzalez-Rabanal, F. and M. Casal, 1995. Effect of high temperatures and ash on germination of ten species from gorse shrubland. Plant Ecol., 116: 123-131.
CrossRefDirect Link - Karayel, D. and A. Ozmerzi, 2008. Evaluation of three depth-control components on seed placement accuracy and emergence for a precision planter. Applied Eng. Agric., 24: 271-276.
Direct Link - Kendle, A.D. and A.D. Bradshaw, 1992. The role of soil nitrogen in the growth oftrees on derelict land. Arboricult. J.: Int. J. Urban For., 16: 103-122.
CrossRef - Kirby, E.J.M., 1993. Effect of sowing depth on seedling emergence, growth and development in barley and wheat. Field Crop Res., 35: 101-111.
CrossRef - Lobato, A.K.S, B.G.S. Filho, R.C.L. Costa, M.C. Goncalves-Vidigal and E.C. Moraes et al., 2009. Morphological, physiological and biochemical responses during germination of the cowpea (Vigna unguiculata Cv. Pitiuba) seeds under salt stress. World J. Agric. Sci., 5: 590-596.
Direct Link - Mahdi, L., C.J. Bell and J. Ryan, 1998. Establishment and yield of wheat (Triticum turgidum L.) after early sowing at various depths in a semi-arid Mediterranean environment. Field Crops Res., 58: 187-196.
CrossRef - Ogbonna, P.E. and I.U. Obi, 2000. Effect of poultry manure and planting date on the growth and yield of Egusi melon (Colocynthis citrullus, L.) in the Nsukka plains of southeastern Nigeria. Samaru J. Agric. Res., 16: 63-74.
Direct Link - Reyes, O. and M. Casal, 2004. Effects of forest fire ash on germination and early growth of four Pinus Species. Plant Ecol., 175: 81-89.
Direct Link - Sangakkara, U.R. and P. Bieler, 1988. Seedling growth of cowpea and mungbean as affected by seed size and seeding depth. J. Agron. Crop Sci., 160: 102-105.
CrossRef - Sangakkara, U.R., 1998. Growth and yields of cowpea (Vigna unguiculata L. Walp.) as influenced by seed characters, soil moisture and season of planting. J. Agron. Crop Sci., 180: 137-142.
Direct Link - Schafferman, D., A. Beharav, E. Shabelsky and Z. Yaniv, 1998. Evaluation of Citrullus colocynthis, a desert plant native in Israel, as a potential source of edible oil. J. Arid Environ., 40: 431-439.
CrossRef - Susana, Z., B. Oscar and A. Orozco-Segovia, 2011. Seed germination of montane forest species in response to ash, smoke and heat shock in Mexico. Acta Oecol., 37: 256-262.
CrossRef - Tordoff, G.M., A.J.M. Baker and A.J. Willis, 2000. Current approaches to the revegetation and reclamation of metalliferous mine wastes. Chemosphere, 41: 219-228.
CrossRef - Franzese, J. and L. Ghermandi, 2011. Seed longevity and fire: Germination responses of an exotic perennial herb in NW Patagonian grasslands (Argentina). Plant Biol., 13: 865-871.
CrossRefDirect Link - Van Hezewij, M.J. and J.A.C. Verkleij, 1996. The effect of nitrogenous compounds on in vitro germination of Orobanche crenata Forsk. Weed Res., 36: 395-404.
CrossRef - Wolfson, B.A.S., T.E. Kolb, C.H. Sieg and K.M. Clancy, 2005. Effects of post-fire conditions on germination and seedling success of diffuse knapweed in Northern Arizona. For. Ecol. Manage., 216: 342-358.
CrossRef - Rivard, P.G. and P. M. Woodard, 1989. Light, ash and pH effects on the germination and seedling growth of Typha latifolia (cattail). Canadian J. Bot., 67: 2783-2787.
CrossRefDirect Link