
Sukesh
Department of Applied Botany, Mangalore University, Mangalagangotri, Mangalore 574199, India
K.R. Chandrashekar
Department of Applied Botany, Mangalore University, Mangalagangotri, Mangalore 574199, India
Research Journal of Seed Science
Year: 2011 | Volume: 4 | Issue: 2 | Page No.: 106-116







ABSTRACT
Hopea ponga (Dennst.) mabberly seeds were selected to study the germination, viability and the biochemical changes during germination of the seeds at different temperatures. The stored seeds were allowed to germinate using rolled paper towel method, biochemical changes during germination and lipid peroxidation value were determined following standard procedures. The loss in the moisture content during storage is a critical factor influences the viability. The decrease in total carbohydrates during storage with increase in lipid peroxidation associated with increased phenolic contents and free amino acids appear to be contributing factors for the viability of seeds. The synergistic effect of reduction in moisture, increased lipid peroxidation, protein modification may be responsible in reducing in the viability of the seeds of Hopea ponga. The lower temperature (14±2°C) is found to be better suited for the storage of these seeds.
PDF Abstract XML References Citation
Received: November 23, 2010;
Accepted: February 07, 2011;
Published: May 14, 2011
How to cite this article
Sukesh and K.R. Chandrashekar, 2011. Biochemical Changes During the Storage of Seeds of Hopea ponga (Dennst.) Mabberly: An Endemic Species of Western Ghats. Research Journal of Seed Science, 4: 106-116.
URL: https://scialert.net/abstract/?doi=rjss.2011.106.116
URL: https://scialert.net/abstract/?doi=rjss.2011.106.116
INTRODUCTION
The Western Ghats, extending along the West Coast of India, covers an area of 180,000 square kilometers. The Western Ghats comprises the major portion of the Western Ghats of India and Sri Lanka which is one of 34 global biodiversity hotspots for conservation and one of the two on the Indian subcontinent. This contains a lot of endemic and endangered species. Some of the forests of Western Ghats have been declared either as National parks or as Reserve forests. There are about 4500 species of flowering plants of the total estimated 17000 species (Ahmedullah and Nayar, 1987). Nearly one third of identified endemic species of the region are rare and threatened and several believed to be extinct (Shetty et al., 2002).
The family, Dipterocarpaceae may account for 10% of all tree species and 80% of all emergent trees in tropical rain forests (Syamsuwida and Owens, 1997). Dipterocarps occur in both aseasonal lowland rain forests where they are evergreen and in seasonal upland or more xerophytic forests where they are deciduous (Smitinand et al., 1980). The family, Dipterocarpaceae has 680 species belonging to 16 genera, distributed in Malaysia, South East Asia, India, Sri Lanka and Africa (Bawa, 1998; Meher-Homiji, 1979). In India, 31 species of Dipterocarps have been reported, mostly confining to North India. The Western Ghats contains 14 species belonging to 5 genera (RAPA Monograph, 1985). Dipterocarpaceae is one of the main components of the evergreen forests of Western Ghats represented by 6 species in the Western Ghats of Karnataka viz., Hopea cannarensis H. Bedd., Hopea ponga (Dennst.) mabberly, Hopea parviflora Bedd., Vatica chinensis L., Vateria indica L. and Dipterocarpus indicus Bedd. (Bhandary and Chandrashekar, 2003; Shivaprasad et al., 1999, 2002; Vasanthraj et al., 2005; Vasanthraj and Chandrashekar, 2006). Except V. chinensis all are endemic to Western Ghats and D. indicus is endangered one.
A striking feature of many dipterocarp forests is the phenomenon of mass flowering followed by mass fruiting (Ashton et al., 1988; Paquette et al., 2009). Large dipterocarps may produce up to four million flowers and 120,000 fruits (Maycock et al., 2005; Brearley et al., 2007). Although the fruits are poorly protected and are eaten in vast numbers by wild pigs and heavily parasitized by weevils managing dipterocarps for seed production is difficult (Ashton, 1988). Flowering commonly does not occur until trees are 20 to 30 years old (Ng, 1966) by which time most of trees grown to a larger height leading the task of management and seed harvest difficult. Fruit production is episodic and because seeds of most species are recalcitrant and store poorly Syamsuwide and Owens, 1997).
Trees of Dipterocarps are being continuously exploited for timber, firewood, construction and other purposes. The seeds of many species of Dipterocarpaceae are generally short-lived and incapable of overcoming desiccation (Aguilar et al., 2006; Saha et al., 1992). The effect of desiccation on germination and vigour of the seeds of Hopea parviflora and Hopea ponga was carried out by Dayal and Kaveriappa (2000). The influence of wing loading and viability of seeds of Hopea ponga was investigated by Muralikrishnan and Chandrashekar (1997). Ashton (1990) had carried out the effect of dewinging, seed maturity, light and soil environment on the germination and survival of Shorea trafiliflora. There has been no report on the biochemical changes during the germination of the recalcitrant seeds of endemic Dipterocarps sp. of the Western Ghats. Therefore, the present study was undertaken to investigate the biochemical changes during seed storage and to find out any possible reason for the poor viability of these seeds.
MATERIALS AND METHODS
Hopea ponga seeds were collected from Western Ghats near Andar Village during 2009-2010. The mature seeds were collected on plastic sheets by shaking the branches. The seeds were immediately brought to the laboratory and used for experiment. The wing of the seed was cut by 1cm length. One set of 1500 seeds were incubated in open environment at 28±2°C. Fourteen sealed polythene bags containing 180 seeds each were incubated separately at different temperatures like 14±2, 21±2 and 28±2°C, respectively.
Every alternate day, one bag each from different temperatures was used for the determination of moisture, germination and biochemical parameters. Moisture content was determined at 105±5°C in hot air oven. Germination percentage was determined by using Rolled paper towel method (ISTA, 1985). The germination percentage, root length and shoot length were measured on 15th day after sowing. Total carbohydrates were estimated following Anthrone method (Sadasivam and Manickam, 2009), reducing sugar was determined by following DNAase method (Sadasivam and Manickam, 2009).
Lipid peroxidation value was determined by using TBA-TCA reagent (Stewart and Bewley, 1980). The total phenol estimation was carried out with the Folin-Ciocalteau reagent. The total amino acid was estimated by following the Ninhydrin method (Sadasivam and Manickam, 2009).
The percentage imbibitional leakage was measured using conductivity meter. For this, ten seeds were placed in 5 mL of Double Distilled (DD) water for 1 h with occasional agitation and further 5 mL of DD water was added and the conductivity was measured. The conductivity of the seeds was measured once again after homogenizing the seeds. The percentage leakage was calculated (Stewart and Bewley, 1980) as:
where, C is percentage leakage, Cl is conductivity of leachate and Ch is conductivity of homogenate.
Statistical analysis: The correlation analysis was carried out with SPSS software package version 12.0. The relationship between the parameters observed was determined by Pearson’s correlation analysis (Zar, 1999).
RESULTS
The germination percentage decreased with increasing number of days of storage. The highest germination percentage on the first day was 99.33% (Fig. 1) which reduced to approximately 50% by 13th day in the seeds stored at different temperatures in polythene cover while the seeds kept in open condition lost their viability by 11th day itself. However, seeds stored in all temperatures lost their complete viability by 19th day except seeds stored at 14±2°C which showed complete loss of viability on 21st day.
The initial moisture of seed was 53.49% on the first day which reduced rapidly in the seeds stored under open condition (Fig. 2). The moisture content reduced gradually in seeds stored in all other temperatures. When the moisture reduced beyond 40% the germination was inhibited. The seeds stored at 14±2°C retained the moisture up to 43-45% on 19th day and the germination was only 13.33%.
The Average Root Length (ARL) and Average Shoot Length (ASL) of growing seedlings of Hopea ponga are given in Table 1. The ASL decreased from 6.304 to 3.52 cm and ARL from 7.303 to 5.28 cm on 9th day in seeds kept in open condition.
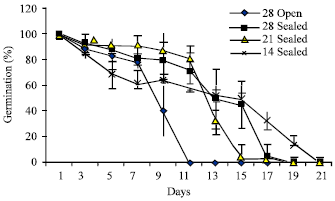 | |
| Fig. 1: | Germination percentage of seeds of Hopea ponga |
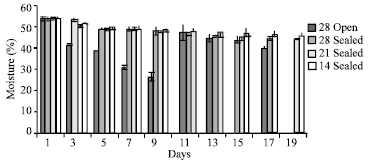 | |
| Fig. 2: | Moisture content of the seeds of Hopea ponga |
| Table 1: | Average Shoot Length (ASL) and average Root Length (ARL) of Hopea pnga seedlings (cm) |
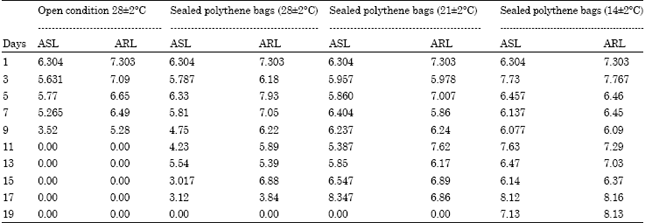 | |
| ASL: Average shoot length, ARL: Average root length | |
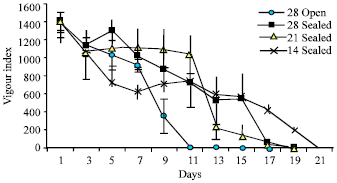 | |
| Fig. 3: | Vigour index of seeds of Hopea ponga |
However, the decrease in the ARL and ASL was not pronounced in the case of seeds stored in different temperatures.
The Vigour index was highest in the seeds kept for germination on the first day (1400.71) and decreased thereafter (Fig. 3). The vigour index decreased from 1400.71 to zero at the end of the 11th day in seeds kept in open condition. There was a slight increment in the vigour index in the seeds kept for germination under sealed condition up to 5th day followed by a decrease and became zero on the 17th day. There was not much variation in the vigour index of the seeds kept under 21±2°C up to 11th day but thereafter vigour index reduced drastically till 13th day and reduced zero on 19th day. The vigour index decreased rapidly up to 7th day in the seeds kept under 14±2°C and later a gradual decrease was observed.
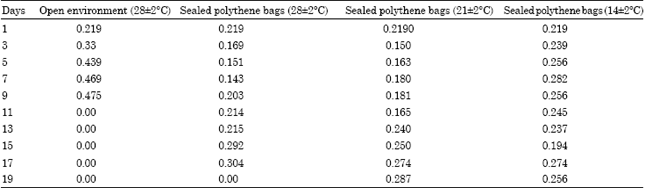
There was a decrease in total carbohydrates during storage in all the treatments (Fig. 4), however, there was a slight increment in total carbohydrates on 5th day and 7th day in seeds kept under 14±2°C. There was an increase in reducing sugar contents and phenolics contents from first day onwards (Fig. 5). The maximum reducing sugar observed on 7th day in the case of seeds stored at 28±2°C and later on it declined. The highest reducing sugar was observed on 15th day in the case of seeds stored at 28±2°C in sealed polythene bags. The reducing sugar was maximum on 17th day in the seeds stored at lower temperature.
Lipid peroxidation activity increased from first day onwards in all seeds (Table 2). However, there was a rapid increment in the lipid peroxidation activity in the seeds stored in open condition.
| Table 2: | Lipid peroxidation in the seed axis of Hopea ponga |
 | |
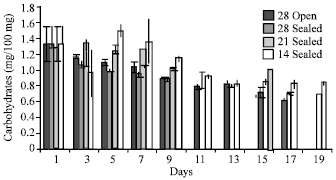 | |
| Fig. 4: | Total carbohydrates in the seeds of Hopea ponga |
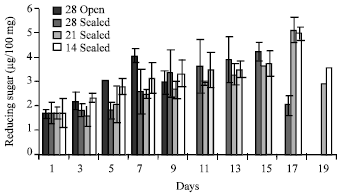 | |
| Fig. 5: | Reducing Sugar in the seeds of Hopea ponga |
Highest lipid peroxidation was observed in the seeds stored at 28±2°C in open environment. However, there was only slight variation in the lipid peroxidation activity in the seeds stored at low temperature.
There was an increase in the phenolic compounds from first day onwards (Fig. 6). A rapid increase in phenolics contents was observed in case of seeds stored at open condition up to 7th day and later decreased. The phenolic content in the seeds stored in 21±2°C increased up to 16.185 μg mL-1 by 17th day and later decreased. There was a steady increase in the phenolic contents in seeds stored at 14±2°C up to 15th day and a slight decrease was seen afterwards. The phenolic contents increased gradually in the seeds stored at 28±2°C in sealed polythene bags.
The electrolyte leakage of the seeds of Hopea ponga is given in Table 3. A rapid increase in the electrolyte leakage was observed in the seeds stored at open condition up to 9th day. A gradual increase in electrolyte leakage up to 19th day was observed in the seeds stored at 28±2°C under sealed condition. The highest electrolyte leakage was observed in the seeds stored at 21±2°C on 17th day followed by a decrease on 19th day. There was not much variation in electrolyte leakage up to 9th day in seeds stored at 14±2°C, later on a slight increase in electrolyte leakage was observed.
The total free amino acids increased with aging of seeds (Table 4). The initial total free amino acids was 3.322 which increased rapidly in seeds kept under sealed condition at 28±2°C and it reached 17.588 on 15th day. In open condition, it was 5.904 on 9th day and in 14±2°C, it was 6.645 on 19th day.
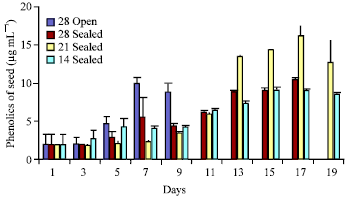 | |
| Fig. 6: | Total phenolics of seed exudates of Hopea ponga |
| Table 3: | Percentage of electrolyte leakage from the seeds of Hopea ponga |
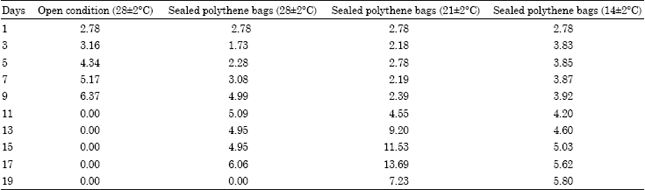 | |
| Table 4: | Total free amino acids μg/100 mL) in the seeds of Hopea ponga |
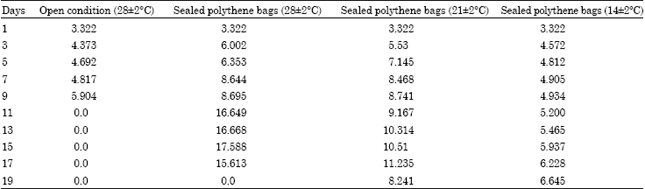 | |
| Table 5: | Pearson correlation table |
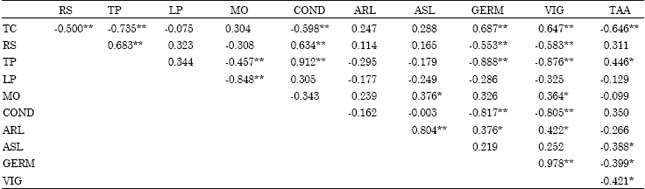 | |
| *: Significant at the 0.05 level (2-tailed). **: Significant at the 0.01 level (2-tailed).TC: Total carbohydrates, RS: Reducing sugar, TP: Total phenolics, LP: Lipid peroxidation, MO: Moisture content, COND: % electrolyte leakage, ARL: Average root length, ASL: Average shoot length GERM: % of germination, VIG: Vigour index, TAA: Total free amino acids | |
There was a positive correlation between the germination and total carbohydrates, total phenolics and reducing sugar, reducing sugar and conductivity, total phenolics and conductivity, ARL and ASL (Table 5).
DISCUSSION
Recalcitrant seeds are those which will germinate immediately soon after they are separated from parent plants. It is widely believed that the seeds of Dipterocarpaceae are generally short lived and sensitive to desiccation (Ashton, 1982; Gunn, 1988; Adjers and Otsamo, 1996). According to Tompsett, (1987) there are two types of seeds in dipterocarps; one type of seed which could not be dried below 45% moisture content without causing damage while the 2nd type could be safely dried to 10-15% moisture content. Dayal and Kaveriappa (2000) reported that the critical moisture contents for Hopea parviflora and Hopea ponga as 26 and 27%, respectively below which they fail to germinate. They have reported that these seeds fail to germinate below respective critical moisture level. The seeds of Hopea ponga stopped germination when moisture level decreased beyond 26%. However, in sealed polythene bags moisture level did not decrease beyond 40% under same temperature of 28±2°C which indicate that there are other factors responsible for viability other than seed moisture. The moisture content in the present study decreased from first day to 26% on the ninth day in the seeds incubated at 28±2°C in an open environment which corroborates with other studies (Dayal and Kaveriappa, 2000). There was a slight decrease in moisture content in seeds incubated at lower temperature. However, the germination percentage declined in the seeds incubated at low temperature. This may be due to the chilling effect on the seeds. Many recalcitrant seeds of Dipterocarps such as Drybalanops aromatica (Jensen, 1971) Shorea curtesii, S. platycladus (Tang, 1971) Shorea ovalis (Sasaki, 1976) and Hopea odorata (Tang and Tamari, 1973) are reported to suffer from chilling injury. According to Roberts (1972), freezing damage in moist seed is presumably associated with the formation of ice crystals.
The reason for the deleterious effect of sub ambient temperature could be due to protein denaturation (Simon et al., 1976) or change in membrane thickness and permeability (Wolfe, 1978). The lower temperature resulted in the decreased in germination percentage in the present study also. The germination percentage decreased rapidly in the seeds incubated at 28±2°C in open condition. The germination percentage increased with decreasing temperature under sealed conditions. The seeds incubated under 14±2°C remained viable till 21st day. Khosh-Khui and Bonyanpour (2006) also observed the maximum germination percentage at 15°C in Cuminum cyminum. Siddique and Wright (2003) reported the effect of high drying temperature on the moisture content and seed quality of Pea seeds. However, there was no effect of temperature and light on the germination of seeds of Ulmus glabra and U. laevis (Cicek and Tilki, 2006).
Saha et al. (1992) reported the sharp decline in the levels of soluble carbohydrates during the desiccation. Sulieman et al. (2007), observed a reduction of some storage nutrients in the lentil seeds during germination. Nautiyal and Purohit (1985) also reported the lower level of soluble proteins in non viable seeds. There was a decline in the total carbohydrates during desiccation. Similar trend was obtained in the present study also.
The reducing sugar content increased with lapse of time and decreasing temperature. There was an increment in total free amino acids during the course of incubation. The increased amino acid content indicated the breaking of proteins. The leaching of soluble sugars and amino acids during the course of incubation was observed by Gupta and Aneja (2004) in soybean varieties. They also observed less leakage of sugars and amino acids in cold condition. The similar results were observed in the present study also. The lower level of soluble proteins in non viable seeds was also observed by Saha et al. (1992) in Shorea robusta.
Lipid peroxidation is considered to be the primary cause for seed deterioration. The rapid increase in lipid peroxidation in seeds stored in open condition at 28±2°C was noticed by other workers in other seeds (Pinzino et al., 1999; AL-Maskri et al., 2003; Salim and Shereena, 2006). Increased in lipid peroxidation in Daucus carota associated with aging was noticed by AL-Maskri et al. (2003). They also observed an increase in electrolyte conductivity with increasing aging time. In the present study, the leakage of electrolyte increased with aging of seeds. Khan et al. (2007) opined that EC test has the potential to be developed as Vigour test in ranking the seed lot quality and to predict the seedling performance in Wheat. The increased lipid peroxidation is considered to be a main factor in deterioration of plasma membrane leading to leaching more electrolytes from the cell followed by protein modification as evidenced by increase in amino acid content in the present study.
CONCLUSION
The germination percentage of Hopea ponga seeds were decreased with laps of time and decreasing temperature. There was a declining trend of carbohydrates and increase in the levels of reducing sugars and total phenolics. The total free amino acids increased with laps of time. The synergistic effect of reduction in moisture, increased lipid peroxidation, protein modification together responsible in reducing the viability of the seeds of Hopea ponga.
ACKNOWLEDGMENTS
The authors are grateful to the Department of Applied Botany, Mangalore University for providing the facilities. The financial support in the form of a major research project sanctioned by U.G.C. is gratefully acknowledged.
REFERENCES
- Aguilar, R., L. Ashworth, L. Galetto and M.A. Aizen, 2006. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett., 9: 968-980.
PubMed - Al-Maskri, A.Y., M.M. Khan, I.A. Khan and K. Al-Habsi, 2003. Effect of accelerated aging on viability, vigor (RGR), lipid peroxidation and leakage in carrot (Daucus carota L.) seeds. Int. J. Agric. Biol., 5: 580-584.
Direct Link - Ashton, P.S., 1988. Dipterocarp biology as a window to the understanding of tropical forest structure. Ann. Rev. Ecol. Syst., 19: 347-370.
CrossRefDirect Link - Ashton, P.S., T.J. Givnish and S. Appanah, 1988. Staggered flowering in the Dipterocarpaceae: New insights into floral induction and evolution of mass fruiting in the aseasonal tropics. Am. Nat., 132: 44-66.
Direct Link - Brearley, F.Q., J. Proctor, L. Nagy, G. Dalrymple and B.C. Voysey, 2007. Reproductive phenology over a 10-year period in a lowland evergreen rain forest of Central Borneo. J. Ecol., 95: 828-839.
CrossRefDirect Link - Cicek, E. and F. Tilki, 2006. Effect of temperature, light and stoarage on seed germination of Ulmus glabra Huds. and U. laevis Pall. Pak. J. Biol. Sci., 9: 697-699.
Direct Link - Dayal, B.R. and K.M. Kaveriappa, 2000. Effect of desiccation and temperature on germination and Vigour of the seeds of Hopea Parviflora beddome and H. Ponga (Dennst.) Mabb. Seed Sci. Technol., 28: 497-506.
Direct Link - Khan, A.M., H. Khan, R. Khan and A. Aziz, 2007. Vigor tests used to rank seed lot quality and predict field emergence in wheat. Am. J. Plant Physiol., 2: 311-317.
CrossRefDirect Link - Khosh-Khui, M. and A.R. Bonyanpour, 2006. Effect of some variables on seed germination and seedling growth of Cumin (Cuminum cyminum L.). Int. J. Agric. Res., 1: 20-24.
Direct Link - Maycock, C.R., R.N. Thewlis, J. Ghazoul, R. Nilus and D. Burslem, 2005. Reproduction of dipterocarps during low intensity masting events in a Bornean rain forest. J. Veg. Sci., 16: 635-646.
CrossRef - Meher Homiji, V.M., 1979. Distribution of the Dipterocarpaceae: Some phytogeographic consideration on India. Phytocoenologia, 6: 85-93.
Direct Link - Muralikrishnan, H. and K.R. Chandrashekar, 1997. Regeneration of Hopea ponga: Influence of wing loading and viability of seeds. J. Trop. For. Sci., 10: 58-65.
Direct Link - Nautiyal, A.R. and A.N. Purohit, 1985. Seed viability in sal. II. Physiologoical and biochemical aspects of ageing in seeds of Shorea robusta. Seed Sci. Technol., 13: 69-76.
Direct Link - Paquette, A., J. Hawryshyn, A.V. Senikas and C. Potvin, 2009. Enrichment planting in secondary forests: A promising clean development mechanism to increase terrestrial carbon sinks. Ecol. Soc., 14: 31-31.
Direct Link - Pinzino, C., A. Capocchi, L. Galleschi, F. Saviozzi, B. Nanni and M. Zandomeneghi, 1999. Aging, free radicals and antioxidants in wheat seeds. J. Agric. Food Chem., 47: 1333-1339.
PubMed - Siddique, A.B. and D. Wright, 2003. Effect of different drying time and temperature on Moisture percentage and seed quality (viability and vigour) of pea seeds (Pisum sativum L.). Asian J. Plant Sci., 2: 978-982.
Direct Link - Salim, N. and J. Shereena, 2006. Chilling tolerance in Pisum sativum L. seeds: An ecological adaptation. Asian J. Plant Sci., 5: 1047-1050.
CrossRefDirect Link - Simon, E.W., A. Minchin, M.M. McMenamin and J.M. Smith, 1976. The low temperature limit for seed germination. New Phytol., 77: 301-311.
Direct Link - Stewart, R.R.C. and J.D. Bewley, 1980. Lipid peroxidation associated with accelerated ageing of soybean axes. Plant Physiol., 65: 245-248.
Direct Link - Syamsuwida, D. and J.N. Owens, 1997. Time and method of floral initiation and effect of paclobutrazol on flower and fruit development in Shorea stenoptera (Dipterocarpaceae). Tree physiol., 17: 211-219.
PubMed - Tompsett, P.B., 1987. Desiccation and storage studies on Dipterocarpus seeds. Ann. Applied Biol., 110: 371-379.
CrossRefDirect Link - Vasanthraj, B.K. and K.R. Chandrashekar, 2006. Analysis of the structure of Charmady reserve forest. Trop. Ecol., 47: 279-290.
Direct Link - Wolfe, J., 1978. Chilling injury in plants: The role of membrane lipid fluidity. Plant Cell Environ., 1: 241-247.
CrossRefDirect Link