
J.G. Ray
Research Laboratory of Ecology and Ecotechnology, School of Biosciences, Mahatma Gandhi University, Kottayam, Kerala, 686560, India
T. Binoy Thomas
Environment Science Research Lab, Postgraduate Department of Botany, St Berchmans College, Changanacherry, Kerala, 686 103 India
Research Journal of Soil Biology
Year: 2012 | Volume: 4 | Issue: 3 | Page No.: 42-68







ABSTRACT
This is the outcome of a pioneer comprehensive investigation of green-algae of Oxic Dystrustepts soils in the Western Ghats of South India, one of the biodiversity hot-spots of the world exposing very many new species. Seventeen percent of the total species observed are quite new to science. Taxonomical characterization of algae in relation to soil and vegetation is carried out systematically from field soils, 'growth slides' and artificial culture of soil samples in specific media. Physico-chemical soil parameters examined were soil-moisture, pH, total carbon, conductivity, total/Kjeldahl nitrogen, plant available potassium and phosphorus. These are correlated to species richness, species diversity index and relative abundance of all green-algae observed. Desmids are found to be the most dominant group of green-algae in these soils. Since green-algae in general are significant biotic component of all ecosystems search of them in these soils is also worthy. Identification and understanding the ecology of them becomes highly relevant from an economic and conservation point of view of algae as well as sustainable management of the fertility of natural soils. Major goal of this investigation was to assess the biodiversity of green-algae in relation to certain physico-chemical characteristics of soils and three different kinds of vegetation such as natural forests, teak and rubber plantations. Results emphasize the worth of immediate exploratory studies throughout the wet tropical soils especially that in the Western Ghats to account the precious biodiversity of green-algae before they get further affected by soil degradations from diverse factors including global climate change.
PDF Abstract XML References Citation
Received: September 22, 2012;
Accepted: January 22, 2013;
Published: February 13, 2013
How to cite this article
J.G. Ray and T. Binoy Thomas, 2012. Ecology and Diversity of Green-algae of Tropical Oxic Dystrustepts Soils in Relation to Different Soil Parameters and Vegetation. Research Journal of Soil Biology, 4: 42-68.
DOI: 10.3923/rjsb.2012.42.68
URL: https://scialert.net/abstract/?doi=rjsb.2012.42.68
DOI: 10.3923/rjsb.2012.42.68
URL: https://scialert.net/abstract/?doi=rjsb.2012.42.68
INTRODUCTION
Algae are common to all kinds of soil ecosystems (Patova and Dorokhova, 2008; Soare and Dobrescu, 2010) and are visible in all tropical soils during favourable seasons. Green-algae are the major groups of algae (Messyasz, 2006) of tropical soils and land manipulations very much affect them (Patova and Dorokhova, 2008). However, green-algal diversity in soils of the Western Ghats, a biodiversity hot-spot of the world remains quite unknown. Algae in soils depend on certain ecological factors (El-Gamal et al., 2008; Mansour and Shaaban, 2010) including vegetation (Neustupa, 2001) and local climate (Chun-Xiang and Yong-Ding, 2003). Significant ecological roles of soil algae include nutrient conservation in soils (Tirkey and Adhikary, 2005), supporting of natural soil formation process (Hoffman et al., 2007), improving of texture and structure (Issa et al., 2007; Soare and Dobrescu, 2010) and acting as partners of numerous food webs (Lukesova and Frouz, 2007). Exact contribution of terrestrial algae to the general carbon pool of soils (Hunt et al., 1979; Hoffman et al., 2007) is yet to be explored. Algae influence germination and growth of seedlings (Singh and Chaudhary, 2011) and yield of crops (Singh et al., 2008). The biologically active secondary metabolites of algae affect soil animals (Metting, 1981) and plants (Schlichting, 1973). They are indicators of soil fertility (Novakovskaya and Patova, 2008; Zancan et al., 2006) and pollution bio-diagnostics (Freystein et al., 2008; Temraleeva et al., 2011; Yoshida et al., 2006). Knowledge of algal community in soils has applications towards conservation of soil and its biota. Study of algae and their ecological roles in soils is highly relevant especially in the current doom of climate change that triggers endangerment and extinction of sensitive species. Therefore, exploration of the biodiversity of them in soils in relation to environment parameters has applications in sustainable management of soil fertility.
Chlorophyta of topsoil are large and diverse and they perform valuable services both in the natural and cultivated soils (Bongale, 1981; Metting, 1981). Among the different groups of terrestrial micro-algae (Rindi et al., 2009), green-algae or Chlorophyta predominate in acidic soils and Cyanophyta in neutral and alkaline soils (Metting, 1981). Therefore, a specific inquiry into the biodiversity and ecology of them is carried out in a slightly acidic tropical wet soil of the Western Ghats (Chandran et al., 2005). Explanation of the population dynamics of them in relation to different kinds of vegetation, seasons and certain physico-chemical soil characteristics of Oxic Dystrustepts of the Western Ghats would serve as basics to the ecological information of tropical soils in general as well as a model for further investigations in this regard.
MATERIALS AND METHODS
Study area description: Algae are collected from ten different study sites of three different kinds of vegetation that included forest soils (F 1-F 10), teak plantation soils (TP 1-TP 10) and rubber plantation soils (RP 1-RP 10), distributed in the mid-upland and high-land regions of Western Ghats (70-350 m msL), Pathanamthitta district (Fig. 1) of Kerala State (9°4’-9 °29’N; 76°28’-77°18’E), South India (Soil Survey Organisation, 2007). Soil sample of each sampling site (1-2 acre land) of a vegetation type was collected from ten different random plots of about 10 m2 size. Climatic and other details of these sites are given in ecological descriptions of soils of the same region (Ray and Binoy, 2012). Among the three kinds of vegetation, the rubber plantations alone receive chemical fertilizer mixture of urea, rock phosphate and muriate of potash (10:10:10) at about 300 kg h-1 twice every year; the first doze during the onset of southwest monsoon and the second during the onset of the northeast monsoon season and the other kinds of vegetation receive no regular cultural or manurial support. Soil sampling for physico-chemical analyses (Jackson, 1973) and the results are already discussed (Ray and Binoy, 2012). Electrical conductivity (E.C) and total/Kjeldal Nitrogen (N) were the additional soil parameters analysed (Jackson, 1973) along with algal studies.
Isolation of algae from soil and the culture: Isolation and culture of soil algae were carried out as per (Mansour and Shaaban, 2010; Zancan et al., 2006) in Modified Bold’s Basal medium (MBBM) prepared as per (Pena-Castro et al., 2004). Solid medium in Petri plates and liquid medium in 250 mL conical flask were incubated at (25±2)°C under continuous light intensity of 4000 lux conditions and allowed to grow for 24-28 days prior to counting and identification process.
 | |
| Fig. 1: | Map of pathanamthitta district |
Biodiversity characterisation: Microscopic study of field soil samples, uni-algal culture of algae from soils in MBBM media (Neustupa, 2001) and ‘growth-slide’ (Zancan et al., 2006) in different seasons from three different kinds of vegetation were used to identify the total biodiversity of algae. Photographs of algae were taken using SONY Digital Camera W 310 attached to the microscope. Algal characterisation and classification were carried out in accordance with the Guiry and Guiry (2011). In addition to that, systematic keys of Iyangar and Desikachary (1981), Philipose (1967), Prescott (1951), Ralfs (1848), Randhawa (1959), Ramanathan (1964) and Pham et al. (2011) were also used. All the algae were identified up to the species/variety.
Environment relationships and population dynamics of algae: Correlations of algae to environmental factors such as vegetation, season and physico-chemical soil parameters are described as per Dey et al. (2010). Relative-abundance, species-richness and diversity-index (Shannon-Wiener Index) of all species of green-algae in each season were worked out.
Diversity Index and species richness: The Shannon-Wiener Index for green-algal diversity has been studied as per Shannon and Weaver (1949) using the formula:
where, Hs is diversity in a sample of S species or kinds, S is the number of species in the sample, Pi is the relative abundance of ith species or kinds, N is total number of individuals of all kinds, ni is number of individuals of ith species, ln is log base 2.
Species richness was calculated (Whittaker, 1977) with the following formula:
where, n is the number of different species in the sample and N is the total number of individual organisms in the sample.
RESULTS AND DISCUSSION
Diversity of Green-algae in relation to vegetations and seasons: Identification on the basis of morphological characteristics revealed the presence of altogether thirty six species of green-algae in these soils which belong to twenty three genera, ten orders and the two Divisions-Chlorophyta and Charophyta Table 1. Among them five are found to have unique characters to be reported as new species and another one as a new variety of already known species.
| Table 1: | Seasonal distribution of soil Green-algae (from the field samples) in Oxic Dystrustepts under the different vegetations |
 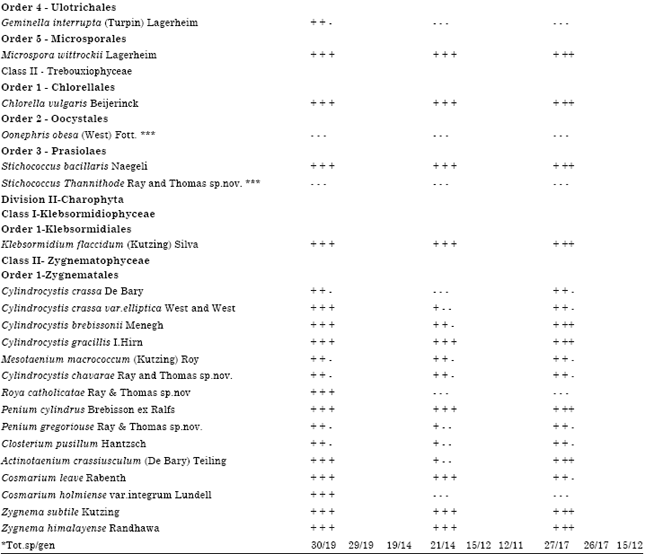 | |
| N.F: Natural Forest, T.P: Teak Plantation, R.P: Rubber Plantation, 1- South West Monsoon, 2-North East Monsoon, 3- Summer, ‘+’: Found in a soil/in a season-Not found in a soil/season, *** Species observed in culture conditions only; *Total species per genera in soils of each vegetation during seasons1,2,3 | |
Discovery that about 17% of algae in a representative region in the Western Ghats of south India are morphologically quite unique and qualified to be called as hitherto unreported new species/varieties emphasizes the worth of immediate exploring of the complete biodiversity of soil algae of the entire region. It may also be noted that the Western Ghats in South India is now considered as one of the ‘hottest’ of the ‘hot-spots’ in the world and is designated as a natural world heritage zone as per the recent declaration of the UNESCO. Even though the exact identity of the new species could not be established using molecular biological means, the unique morphological characteristics observed are still valid in the identification of new species and varieties of algae (Flint and Williamson, 2010). Green-algae represent only a part of the actual biodiversity of the soil algal community and the details of the other algal groups from this area are under the process of description and identification. It has become crystal clear that Oxic Dystrustepts in the Western Ghats has high diversity of green-algae, quite unlike the temperate soils, where algal diversity is poor (Lukesova, 2001).
Since, the biogeography of terrestrial algae, its diversity and ecology still remain rudimentary and deserve great attention (Neustupa and Skaloud, 2010; Rindi, 2007; Rindi et al., 2009), the present discovery of green-algal community of this ecologically sensitive tropical soil is highly relevant new knowledge. This is especially important because the ecology and diversity of soil algae of tropics is quite scarce (Sharma et al., 2006) while that of temperate soil environments is well-known (Neustupa, 2001; Neustupa and Skaloud, 2008). The current accounting of algae of soils on the basis of field studies, growth-slides and soil-culture enabled complete exploration of algae in a natural environment (Mansour, 2008; Rindi et al., 2009). Moreover, the mandatory frequent and regular soil sampling over one year period (Broady, 1979) analyzing the influences of many soil parameters on green-algae was comprehensive. Overall, this is a significant representative model record of algal diversity of wet tropics in general.
Soil algae are highly variable in morpho-taxonomic characteristics under diverse environmental influences (Ohtani et al., 2000). The highly undulating topographic terrains of the Western Ghats (Soil Survey Organisation, 2007) are quite conducive for different kinds of physico-chemical environmental conditions of the region. The high complexities of diverse abiotic regimes in the zone lead to existence of highly diverse ecological niches which contributes to the ‘hot-spot’ status of the region. The Western Ghats is known for high degree of ‘α’, ‘β’ and ‘γ’ kinds of biodiversity. The present report of high degree of green-algal diversity from Pathanamthitta District, a small representative area of the zone agrees with this kind of an expectation in the biodiversity aspect of the region. Details of the algal diversity observed are discussed below.
Division I-chlorophyta: Altogether twenty species of Chlorophyta belonging to fourteen genera of two classes Chlorophyceae and Trebouxiophyceae were observed in these soils; two were found to be new species and one a new variety. They belonged to the:
Class 1-chlorophyceae: There were sixteen species of this class in these soils which belong to five orders and seven families.
Volvocales: All the three species of this order were from the family Chlamydomonadaceae. One of them is a common species and two of them are quite new species/variety. The most common species identified as per Prescott (1951) from this soil was Chlamydomonas globosa Snow (Fig. 2A(a)-2B(b)) which was found in soils from natural forest and rubber plantations in all seasons but found only in the southwest monsoon season in teak plantation. However, the species appeared in large numbers in the cultures of soils from forest, teak and rubber plantations in all seasons. The new species found out was Chlamydomonas ovoidae Ray and Thomas sp. nov. (Fig. 2A(b); Silva, 2012). It was observed directly from the field soil samples of natural forest and rubber plantations and also could be cultured (MBBM) from soils from those field samples. It was found in field samples during all seasons in the natural forest, during the southwest monsoon and summer in rubber plantations but it was never found in the field soils or its culture from teak plantations.
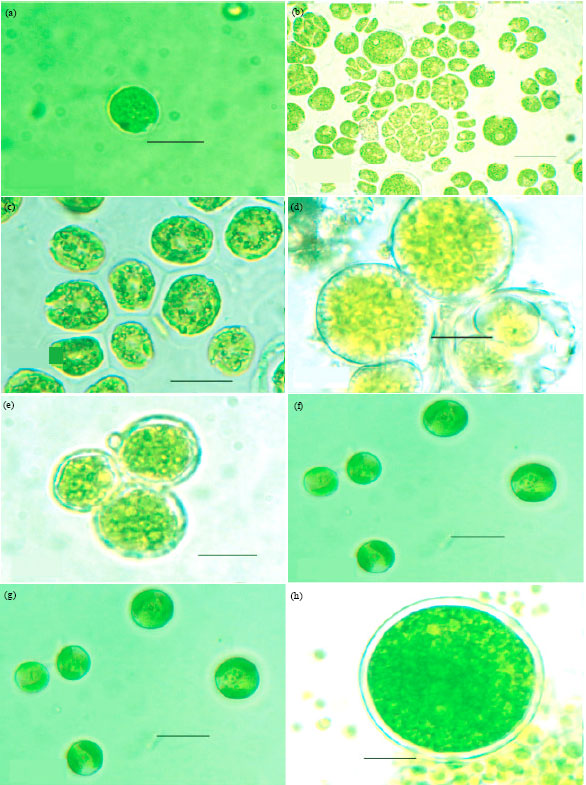 | |
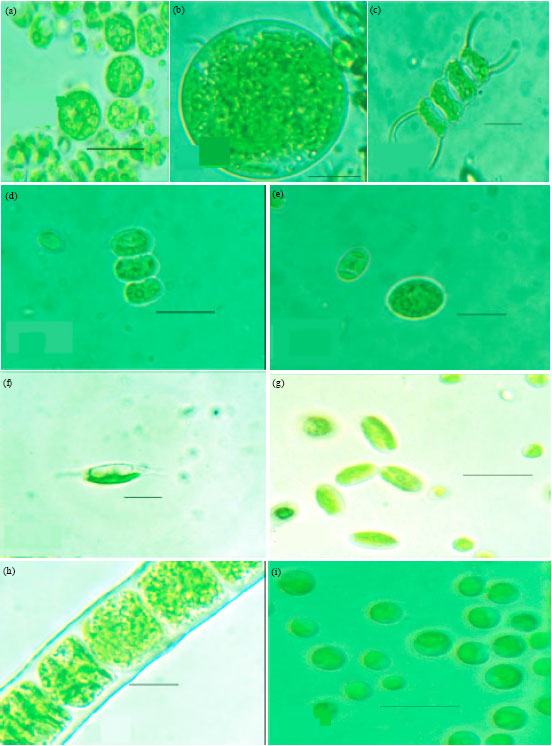
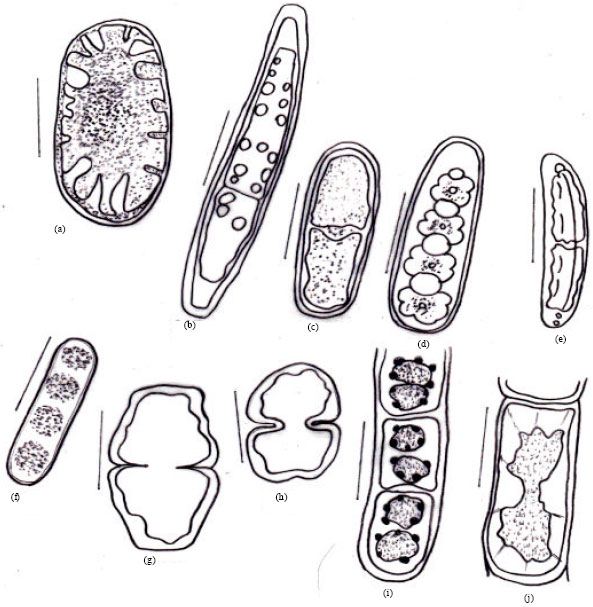
| Fig. 2A(a-h): | Microphotographs: (a) Chlamydomonas globosa (b) Chlamydomonas ovoidae (c) Chlamydomonas deasonii var. micro (d) Chlorococcum infusionum (e) Chlorococcum echinozygotum (f) Chlorococcum humicola (g) Myrmecia bisecta (h) Dictyochloropsis splendida |
This species collected from the current soils showed the typical characteristics such as unicellular of 5-10 μm wide, 7-8 μm long, spherical, green coloured and cup shaped single chloroplast with a single thick pyrenoid located at the posterior half of the cell body, eye spot at the anterior of cells embedded in chloroplast, no papilla, a thick sheath surrounding the cell as well as the zoosporangia, cell division producing 8-16 zoospores and the zoospores arranged in groups of four.
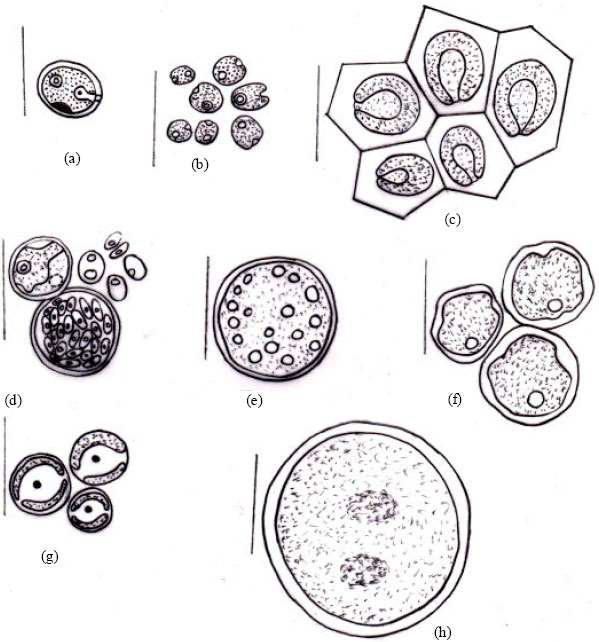 | |
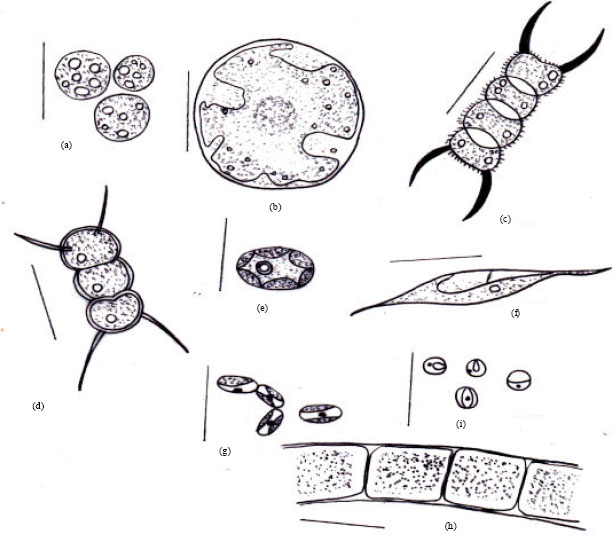
| Fig. 2B(a-h): | Diagrams: (a) Chlamydomonas globosa (b) Chlamydomonas ovoidae (c) Chlamydomonas deasonii var. micro (d) Chlorococcum infusionum (e) Chlorococcum echinozygotum (f) Chlorococcum humicola (g) Myrmecia bisecta (h) Dictyochloropsis splendida |
Unlike the closely related species of Chlamydomonas debaryana Goroschankin (Guiry and Guiry, 2011), the current species has some unique characters (Fig. 2B(b)), such as spherical to ovoid shape (versus ellipsoid or sub spherical), large basal thick ovoid pyrenoid (versus single small pyrenoid) and with rudimentary papilla (versus long papilla). Therefore, this species is named as C. ovoidae to recognize the ovoid shape of both the vegetative cell as well as its pyrenoid.
 | |
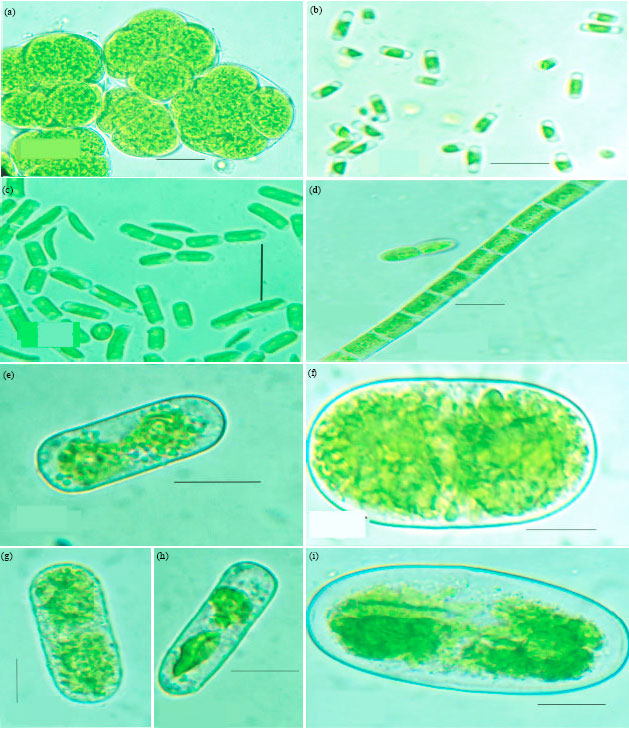
| Fig. 3A(a-i): | Microphotographs: (a) Neospongiococcum vacuolatum (b) Radiosphaera dissecta (c) Scenedesmus perforates (d) Scenedesmus quadricauda var. quadrispina (e) Coelastrella terrestris (f) Keratococcus bicaudatus (g) Geminella interrupta (h) Microspora wittrockii (i) Chlorella vulgaris |
The new variety found out was Chlamydomonas deasonii var. micro Ray and Thomas var. nov. (Fig. 2A(c); Silva, 2012). It was observed directly from the field soil samples but could not be cultured (MBBM) from field soil samples. This species was found in soils of natural forest and rubber plantations alone and not in soils of teak plantations.
 | |
| Fig. 3B(a-h): | Diagrams: (a) Neospongiococcum vacuolatum (b) Radiosphaera dissecta (c) Scenedesmus perforates (d) Scenedesmus quadricauda var. quadrispina (d) Coelastrella terrestris (e) Keratococcus bicaudatus (f) Geminella interrupta (g) Microspora wittrockii (h) Chlorella vulgaris |
This alga was observed only occasionally and that too in the two wet seasons alone but never found in the summer. This species showed typical characteristic features like unicellular cells which are closely packed and embedded in rigid mucilage of polygonal shape. The cells are 10-16 μm in diameter and 10-11 μm in long, cells are semi-spherical, dark-green colour, with anterior conical and posterior broad ends, wide apical opening, conical papilla present, many pyrenoids are distributed along the margins of the cup shaped chloroplast, eye spot located at the anterior side. This variety differs from a closely related species Chlamydomonas deasonii Ettl Guiry and Guiry (2011) by having smaller dimensions (versus bigger dimensions; 6-23 μm wide, 16-27 μm long), semi spherical (versus ellipsoidal shape), cells closely packed and embedded in a rigid mucilage of polygonal shape (versus thin mucilage of polygonal outer structure) wide apical opening (versus narrow opening). These unique morphological features (Fig. 2B(c)) suggest that this is a new variety and named as C. deasonii var. micro.
 | |
| Fig. 4A(a-i): | Microphotographs: (a) Oonephris obesa (b) Stichococcus bacillaris (c) Stichococcus Thannithode (d) Klebsormidium flaccidum (e) Cylindrocystis crassa (e) Cylindrocystis crassa var. elliptica (f) Cylindrocystis brebissonii (g) Cylindrocystis gracillis (h) Mesotaenium macrococcum |
Chlorococcales: There were seven species from this order and all belonged to the family Chlorococcaceae. The seven known species of this family were (1) Chlorococcum (Schrank) Meneghini (Fig. 2A(d)-2B(d)) identified as per Skaloud (2009).
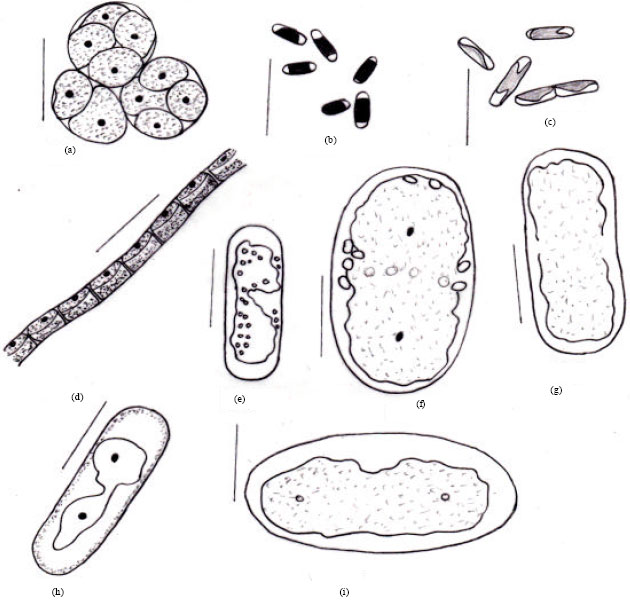 | |
| Fig. 4B(a-i): | Diagrams: (a) Oonephris obesa (b) Stichococcus bacillaris (c) Stichococcus Thannithode (d) Klebsormidium flaccidum (e) Cylindrocystis crassa (f) Cylindrocystis crassa var. elliptica (g) Cylindrocystis brebissonii (h) Cylindrocystis gracillis (i) Mesotaenium macrococcum |
This is found in field soil samples as well as in the cultures of those samples (MBBM) from natural forest and rubber plantation but never in the teak plantations. This was one of the very abundant soil algal species of forests during the southwest monsoon season. (2) Chlorococcum echinozygotum Starr (Fig. 2A(e)-2B(e)) identified as per Guiry and Guiry (2011). This is found in field samples of all vegetations including forest, teak and rubber in both the monsoon seasons and also in the culture (MBBM) of soil samples of all seasons. This is a very common species found in the soil as well as on other terrestrial platforms like stones, bricks, cement walls and the like, in the region. (3) Chlorococcum humicola (Naegeli) Rabenhorst (Fig. 2A(f), 2B(f)) identified as per Guiry and Guiry (2011).
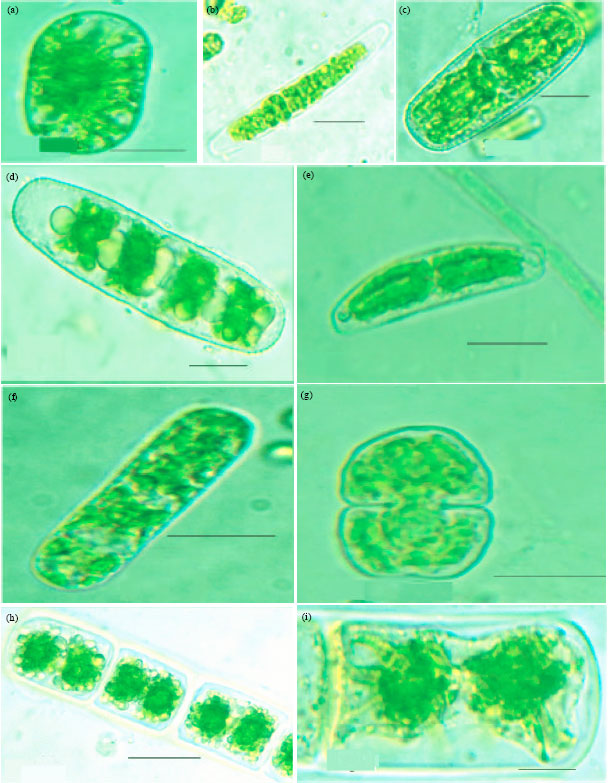 | |
| Fig. 5A(a-i): | Microphotographs: (a) Cylindrocystis chavarae (b) Roya catholicatae (c) Penium cylindrus (d) Penium gregoriouse (e) Closterium pusillum (f) Actinotaenium crassiusculum (g) Cosmarium leave (h) Cosmarium holmiense var. integrum (i) Zygnema subtile (j) Zygnema himalayense |
This is found in field soil samples as well as the cultures of those samples (MBBM) from natural forest (field observation in southwest monsoon only) and rubber plantation (field observation in southwest and northeast monsoons) and never in field soils or its cultures from teak plantations.
 | |
| Fig. 5B(a-j): | Cylindrocystis chavarae (a) Roya catholicatae (b) Penium cylindrus (c) Penium gregoriouse (d) Closterium pusillum (e) Actinotaenium crassiusculum (f) Cosmarium leave (g) Cosmarium holmiense var. integrum (h) Zygnema subtile (i) Zygnema himalayense |
This was one of the very abundant soil algal species of forests during the southwest monsoon season (4) Myrmecia bisecta Reisigl (Fig. 2A(g), 2B(g)) identified as per Khaybullina et al. (2010). This is a species never found in field samples but appeared in cultures (MBBM) from all the field samples of all seasons (5) Dictyochloropsis splendida Geitler var. splendida (Fig. 2A(h), 2B(h)) identified as per Khaybullina et al. (2010) and Skaloud (2009). This was found in field soil samples from all vegetations and also in the culture (MBBM) of them in all seasons (6) Neospongiococcum vacuolatum Deason and E.R. Cox (Fig. 3A(a), 3B(a)) identified as per Guiry and Guiry (2011). This was found in field soil samples from all vegetations and also in the cultures (MBBM) of them in all seasons. (7) Radiosphaera dissecta (Korschikov) Starr (Fig. 3A(b), 3B(b)) identified as per Guiry and Guiry (2011). This species never occurred in field samples but always found in the culture (MBBM) of the field soil samples from all seasons.
Sphaeropleales: Altogether four species were observed from this order, three of which were from the family Scenedesmaceae and one from Ankistrodesmaceae. The three species from Scenedesmaceae family were: (1) Scenedesmus perforates Lemmermann (Fig. 3A(c), 3B(c)) identified as per Guiry and Guiry (2011). This alga was never observed in field soil samples but appeared in the cultures (MBBM) of field samples from forest, teak and rubber plantations during all the seasons in large numbers. (2) Scenedesmus quadricauda var. quadrispina (Chodat) G.M. Smith (Fig. 3A(d), 3B(d)) identified as per Guiry and Guiry (2011). This species appeared in cultures (MBBM) of field samples from all the three vegetations during all the seasons in large numbers but never observed in field samples. (3) Coelastrella terrestris Reisigl Hegewald et Hangata (Fig. 3A(e), 3B(e)) identified as per Tschaikner et al. (2007). This species was found in soils from natural forest and rubber plantations in monsoon seasons and also in the cultures of these soil samples. A species from Ankistrodesmaceae family found out was Keratococcus bicaudatus (A. Braun ex Rabenhorst) J. B. Petersen (Fig. 3A(f), 3B(f)) identified as per Zidarova (2008) was the only species belonging to this family observed very common in these tropical soils. It was found in the soil samples of this area from all the vegetations during all seasons and also in the cultures (MBBM) of those soil samples.
Ulotrichales: There was only a single species from this order, belonging to the family Gloeotilaceae found in these soils. The species observed was Geminella interrupta (Turpin) Lagerheim (Fig. 3A(g), 3A(g)) identified as per Skaloud (2009). This is observed in field samples of forest soils alone and that too during the monsoons only. It also appeared in the culture (MBBM) of field soil samples of forest from all seasons but never appeared in the culture of soils from both the plantations.
Microsporales: There was only a single species from this order, belonging to the family Microsporaceae found in these soils. It was Microspora wittrockii Lagerheim (Fig. 3A(h), 3B(h)) identified as per Zarina et al. (2006). This is one of the very common species found in all the field samples from natural forest, rubber and teak plantations and also in the culture (MBBM) from these field samples during all seasons.
Class 2-Trebouxiophyceae: There were four species of this class in these soils which belonged to the three orders and three families.
Chlorellales: There was only one species observed from this order, belonging to the family Chlorellaceae, Chlorella vulgaris Beijerinck (Fig. 3A(i)) identified as per Guiry and Guiry (2011). This is a cosmopolitan species found in all soil samples from all the three vegetations during all seasons and also in the cultures (MBBM) of these soil samples.
Ooocystales: There was only one species observed from this order, belonging to the family Oocystaceae which was Oonephris obesa (West) Fott. (Fig. 4A(a), 4B(a)) identified as per Guiry and Guiry (2011). It was never observed in the field soils but always found in the cultures (MBBM) of soils from all vegetations and seasons.
Prasiolales: Altogether two species were observed from this order in these soils, both belonging to the family Prasiolaceae. One of these was found out to be a new species, possessing unique characters and the other was a well known species. The well known species from this family observed in these soils was Stichococcus bacillaris Naegeli (Fig. 4A(b), 4B(b)) identified as per Khaybullina et al. (2010) and Ohtani et al. (2000). It is a cosmopolitan species, found in all soil samples from all the three vegetations during all seasons and also in the culture (MBBM) of these soil samples. The new species with unique characters observed was Stichococcus Thannithode Ray and Thomas sp.nov. (Fig. 4A(c); Silva, 2012). This species appeared in the culture (MBBM) of all the soil samples but never appeared in the field samples. The typical characters of the species are pale green, unicellular, cells cylindrical with rounded ends, 6-7 μm long and 1-2 μm wide with a long parietal and folded plate-like chloroplast having smooth margins and one pyrenoid. Cells are arranged in a chain but unbranched. This species differs from the known Stichococcus bacillaris Naegeli (Guiry and Guiry, 2011) in features such as:
| • | Unlike 2-4 μm width in the known species, this species is of narrow cells of 1-2 μm width |
| • | Unlike parietal chloroplast in known species, this species has parietal and folded plate-like chloroplast |
| • | Presence of a pyrenoid which is quite absent in the known species (Fig. 4B(c)). Therefore, this was named as a new species Stichococcus Thannithode |
Division II-Charophyta: Altogether sixteen distinct species belonging to two classes, six families, two orders and nine genera were observed in these soils. Among them, three were found to be new species with unique features.
Class 1-Klebsormidiophyceae: There was only one species of this class from the order Klebsormidiales, belonging to the family Klebsormidiaceae, Klebsormidium flaccidum (Kutzing) Silva PC, Mattox KR and Blackwell (Fig. 4A(d), 4B(d)) identified as per Khaybullina et al. (2010) and Skaloud (2009). This is a cosmopolitan species, found in all field soil samples from all the three vegetations during all seasons and also in the culture (MBBM) of these soil samples.
Class 2-Zygnematophyceae: There were fifteen species of this class in these soils which belong to the five families of a single order Zygnematales.
Mesotaeniaceae: There were seven species from this family, of which two were identified as new species. The common well known species found in these soils were (1) Cylindrocystis crassa De Bary (Fig. 4A(e), 4B(e)) identified as per Guiry and Guiry (2011). This is found in field soil samples from natural forest and rubber plantations during the monsoons but never from teak plantations in any season. This alga also appeared in the cultures (MBBM) of soils from natural forest and rubber plantations during all seasons but never from soils of teak plantations. (2) Cylindrocystis crassa var.elliptica West and West (Fig. 4A(f), 4B(f)) identified as per Khondker et al. (2008). This is found in natural forest and rubber plantations in all seasons but in teak plantations during the southwest monsoon season alone. It appeared in the culture (MBBM) of soil samples from all the three vegetations during all the seasons. (3) Cylindrocystis brebissonii Menegh (Fig. 4A(g), 4B(g)) identified as per Khondker et al. (2008) and Opute (2000). This is found in field soil samples of all the seasons in natural forest and rubber plantations whereas, during the monsoons alone in teak plantation. It also appeared in cultures (MBBM) of field soil samples from all the three vegetations during all the seasons. (4) Cylindrocystis gracillis I. Hirn (Fig. 4A(h), 4B(h)) identified as per Guiry and Guiry (2011). This is found in all soil samples from all the three vegetations irrespective of the seasons but never found in cultures. (5) Mesotaenium macrococcum (Kutzing) Roy ET Bisset (Fig. 4A(i), 4B(i)) identified as per Khaybullina et al. (2010). This is one of the very common species found in field soil samples from all vegetations and also in the culture (MBBM) of them in both the monsoon seasons but absent in the field soils of summer and its culture.
The new species found out from this family was Cylindrocystis chavarae Ray and Thomas sp.nov. (Fig. 5A(a); Silva, 2012). This was one of the very common species found in the field samples from soils of all the three vegetations during the monsoon seasons and in the culture (MBBM) of soils from all seasons. This specimen was unicellular, cylindrical, unconstructed, loosely attached with each other to form filamentous colony, both ends rounded, cells 32-33 μm long, 17 μm wide. The length/breadth ratio of this specimen is 1.8. There are two chloroplasts which appeared as one, stellate with out pyrenoids. The nucleus is centrally placed.
This species was found quite distinct from the related species such as C. brebissonii and C. crassa (Guiry and Guiry, 2011) in the following features. (1) The nature of chloroplast-two of them appeared as one with very conspicuous 10-14 astral-strands quite unlike the two chloroplasts and few radiating prolongations which are slightly visible in the known species (2) length to breadth ratio below 2 unlike 2-4 in the known species (Fig. 5B(a)). Therefore, this alga is given the status of a new species Cylindrocystis chavarae.
The second new species found out from this family was Roya catholicatae Ray and Thomas sp. nov. (Fig. 5A(b); Silva, 2012). It was found in field soil samples of all seasons and also in the culture (MBBM) of those soil samples from natural forest but never in soils of teak or rubber plantations either in field soil samples or in its cultures. The typical characters of the species include unicellular and spindle-shaped with truncated ends, 50 μm long, 6 μm wide, length/ breadth ratio 8.3, single chloroplast, arranged in ribbon-like manner with many pyrenoids and centrally placed single nucleus. No central notch is found in the chloroplast. Vacuolated region is found at both apices of the cell (Fig. 5B(b)).
The quite distinct features of this alga from its closely related species such as Roya obtusa (Brebisson) West and West and Roya anglica G. S. West Guiry and Guiry (2011) include spindle shaped cell (versus slightly curved or straight with broadly rounded or truncated ends of the known species), single notch-less chloroplast per cell (versus two chloroplast with central notch of the known species), chloroplast not extended to the apices of the cell (versus elongated and ribbon shaped chloroplast extended to the apices of the known species). Because of these distinct features from the related known species, it was named as a new species R. catholicatae.
Peniaceae: Altogether two species of algae from this family were found out, one was a well known species and the other was found to be quite new to science. The one common species was Penium cylindrus Brebisson ex Ralfs (Fig. 5A(c), 5B(c)) identified as per Opute (2000) found in the soils of natural forest, teak and rubber plantations and also present in cultures (MBBM) of these soils in all seasons.
The new species found out was Penium gregoriouse Ray and Thomas sp. nov. (Fig. 5A(d); Silva, 2012). This species was found in the soil samples from all the vegetations of the monsoon seasons alone and also its culture (MBBM). The typical characters of this alga include unicellular cylindrical cells, 48-50 μm long, 12-14 μm wide, having distinct girdle bands in the cell wall, cell wall quite thick with regularly arranged granules or longitudinal ridges, five vacuoles and four chloroplasts with pyrenoids per cell (Fig. 5B(d)).
The unique features of this alga from the closely similar species Penium terrestre Jao (Guiry and Guiry, 2011) include (1) Four chloroplasts with five vacuoles-three in between and two at the end, without longitudinal ridges unlike two chloroplasts with longitudinal ridges in known species and (2) Multiple pyrenoids unlike 1-2 axial pyrenoids in known species. Therefore, this species is named as P. gregoriouse.
Closteriaceae: There was only a single species from this family in this soil, Closterium pusillum Hantzsch. (Fig. 5A(e), 5B(e)) identified as per Guiry and Guiry (2011). This was found in the field samples of natural forest, teak and rubber plantations during the monsoon seasons, also appeared in the cultures (MBBM) of soil samples of all the vegetations in the monsoon seasons.
Desmidiaceae: There were three well known species of algae from this family observed in this soil. (1) Actinotaenium crassiusculum (De Bary) Teiling (Fig. 5A(f), 5B(f)) identified as per Guiry and Guiry (2011). This was found in field soil samples of all the seasons in the natural forest and rubber plantations whereas during the monsoons alone in teak plantation; but it appeared in cultures (MBBM) of field samples from all the three vegetations during all the seasons in limited numbers. (2) Cosmarium leave Rabenth (Fig. 5A(g), 5B(g)) identified as per Flint and Williamson (2010), found in the field soil samples of all vegetations during all the seasons and also in the culture (MBBM) of all soil samples. (3) Cosmarium holmiense var.integrum Lundell (Fig. 5A(h), 5B(h)) identified as per Guiry and Guiry (2011), found in field soil sample of only forests, in all seasons and also in its culture (MBBM).
Zygnemataceae: Two species of desmids belonging to this family were observed in this soil. Both of them are well known species (1) Zygnema subtile Kutzing (Fig. 5A(i), 5B(i)) identified as per Guiry and Guiry (2011) and (2) Zygnema himalayense Randhawa (Fig. 5A(j), 5B(j)) identified as per Zarina et al. (2006) were found to be cosmopolitan species, found in all field samples from all vegetations of all seasons and also in the cultures (MBBM) of all those soil samples.
Relative abundance, species richness and diversity index
Relative abundance: Relative abundance of a species was calculated by the formula:
where, X is total number of samples collected, Y is No. of samples from which soil algae was isolated Dey et al. (2010).
Out of the 90 samples collected from three different vegetations during different seasons, the percentage of field occurrence of each and every species was calculated for all the thirty six diverse species. The green-alga with the highest relative abundance in this soil was Chlorella vulgaris (63.33%) followed by Coelastrella terrestris (62.22%), Dictyochloropsis splendida (54.44%), Keratococcus bicaudatus (51.11%) and Zygnema himalayense (51.11%).The least abundant soil alga was Roya catholicatae (6.6%) (Table 2).
| Table 2: | Relative abundance (%) of occurrence in field samples only) of soil green-algae in Oxic Dystrustepts under the three different vegetations |
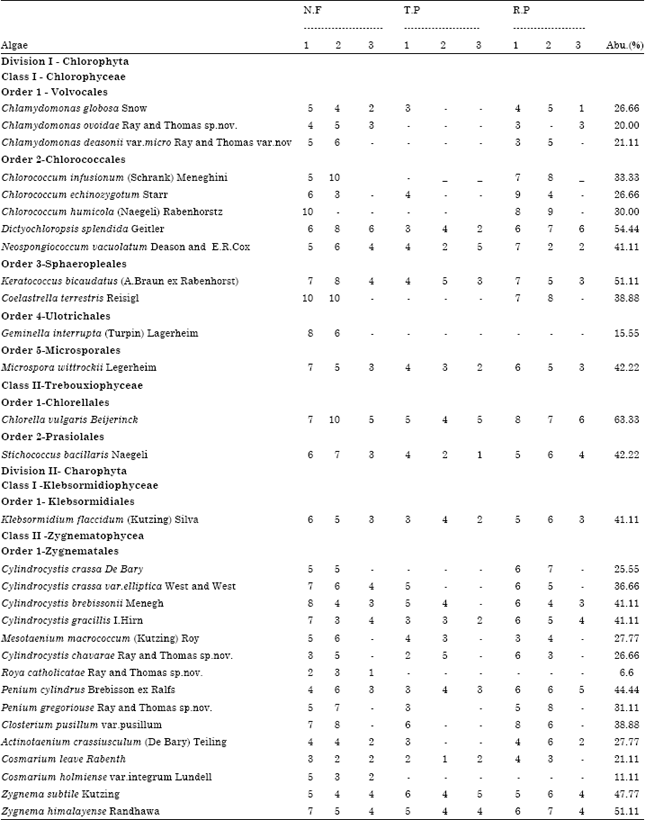 | |
| N.F: Natural Forest, T.P: Teak Plantation, R.P: Rubber Plantation, 1- South West Monsoon, 2-North East Monsoon, Abu - Abundance | |
| Table 3: | Species richness and diversity index of soil Green-algae in different seasons and vegetations |
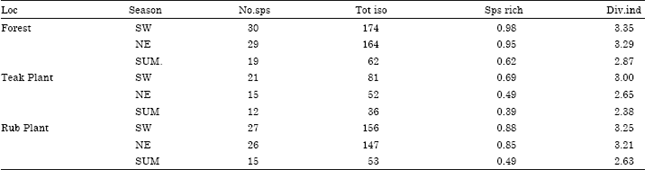 | |
| Loc - Location, No. sps-Number of species, Tot iso-Total isolate, Sps rich- Species richness, Div. Index-Diversity index (Shannon-Wiener Index), S.W: South West Monsoon, N.E : North East Monsoon, SUM-Summer, Teak Plant-Teak Plantation, Rub Plant - rubber plantation | |
| Table 4: | Average physico-chemical properties of Oxic Dystrustepts soils from different vegetations |
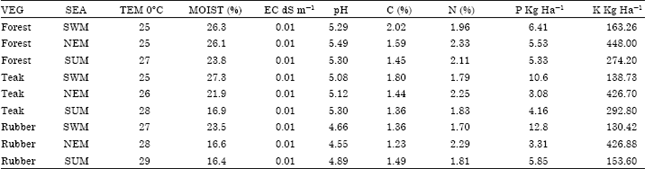 | |
| Abbreviations: VEG-Vegetation, SEA-Season, TEM-Temperature, MOIST-Moisture, E.C-Electrical conductivity; SWM-South West Monsoon, NEM-North East Monsoon, SUM-Summer | |
In all seasons, natural forest showed the highest abundance of algal flora than teak and rubber plantations. Soil in all the three vegetations yielded the maximum number of isolates of diverse species during the monsoon seasons. In general, forest vegetations had the highest number of algal isolates than both the plantations in all seasons.
Species diversity index and Species richness (Table 3): The range of variation of the diversity index of green algae from summer to south west monsoon seasons in different soils was 2.87-3.35 in forest, 2.38-3.00 in teak plantation and 2.63-3.25 in rubber plantation. Among the three vegetations, forest had the highest green algal diversity index than the teak and rubber plantations in all the three seasons. The diversity of specimens was highest in the southwest monsoon season in all the three vegetations. Soil algal diversity was the least in the summer season under forest, teak and rubber. Species richness of natural forest (0.85) was higher than rubber plantation (0.74) and teak plantation (0.52).
Physico-chemical properties of soils: Average physico-chemical characteristics of the filed soil samples from different vegetations are given in Table 4. Except that of soil nitrogen, all the other parameters in this table are as per Ray and Binoy (2012).
Environmental correlations: Correlations of green-algal Diversity Index (DI) to soil characteristics such as pH, C, N, P, K and Moisture at different sampling sites and at different seasons using Pearson correlation coefficient were determined. A significant positive correlation exists between DI-P, Moisture (p<0.01) and C (p<0.05) in all the three sampling sites. However, no significant correlation was observed between DI and the other soil characteristics such as N, K and pH.
Physico-chemical factors of soils have been considered as significant factors in determining soil algal distribution (Fujita and Nakahara, 2006; Starks and Shubert, 1982). However, in the current observation, apart from C, P and soil moisture, no significant correlations were found between the other individual soil parameters such as soil N, K and pH to green-algal diversity index in different vegetation. According to Neustupa and Skaloud (2008), it is light that mainly influences the diversity of algae. But in teak plantations where trees remain leafless and the soil getting more exposed to sun light during summer, a corresponding increase in soil algal diversity was not observed. Regardless of sporadic summer rains received, summer is more or less a dry season in this part of Western Ghats, without sufficient soil moisture. This observation point to the fact that soil moisture conditions and temperature also affect soil algal communities under different vegetation, especially when light is not a limiting factor. Many algae in soils are adapted to live without much water during their vegetative phase but their reproductive stages are completed in presence of water (Lewis and McCourt, 2004). A decrease in soil moisture content in teak plantations from that of rubber plantations and natural forests may also be attributed to the general decrease of species diversity of green-algae of soils in the former kinds of vegetation.
The annual fluctuations in soil pH observed were 4.55-5.49 but the seasonal changes at any particular soil were quite negligible. Influence of pH on soil algae is well known (Nayak and Prasanna, 2007; Zancan et al., 2006). In general, pH and EC affect the availability of soil nutrients and in turn affect the biodiversity of soil algae (Mansour and Shaaban, 2010). Green-algae prefer acidic soils than alkaline and neutral soils (Schlichting, 1973) and the high diversity of green-algae observed in the present zone may be attributed to this fact. Green-algae are considered to be the most tolerant group of algae resistant to adverse soil conditions like high acidity, loss of structure, water logging and poor nutrient conditions of soils (Hoffman et al., 2007; Zancan et al., 2006). Unlike the very low diversity of algae found in highly acidic podozolic soils (Novakovskaya and Patova, 2008), a high diversity of green-algae was observed in the slightly acidic Oxic Dystrustepts under all the three different vegetations in the Western Ghats.
One of the very important environmental parameters in relation to algal community observed in the current investigation was that of vegetation. According to Bohlen et al. (2001) and Bongale (1981), in a particular soil type of a specific climate condition, algal community structure is influenced by specific vegetation of the locality. The most influential ecological factors on the growth and development of green-algae must be microclimatic conditions (Sharma et al., 2006) determined by light and humidity (Neustupa and Skaloud, 2010), temperature and moisture (Shimmel and Darley, 1985), all of which are highly influenced by vegetation. The whole zone of current investigation belonged to a single soil type (Oxic Dystrustepts) under three different vegetations. The significant interrelationships observed in the current investigations between vegetation and abundance of green-algal community reveal the negative impact of monoculture plantations on green-algal biodiversity in wet tropical soils. It may be noted that teak plantations have more negative influence on algal biodiversity than rubber plantations, even though the soils under the latter vegetation are subjected to more intensive cultural and manurial practices. This is in agreement with the observation of Zancan et al. (2006) that a regular application of fertilizer increases the diversity and number of soil algae in the field, even though intensive application leads to the reduction of species diversity. However, a general decrease in the biodiversity of green-algal community in plantations from that of the forests point to the significant positive influence of virgin natural vegetation on green-algal community structure in tropical soils.
Lemes-Da-Silva et al. (2010) observed a decrease in algal masses in forests soils covered by fallen leaves, twigs and decomposing residues. However, the overall diversity of algal species in the tropical forest was found to be higher than that of plantations in the current observations. This is in agreement with the observations of Fujita and Nakahara (2006), Metting (1981) and Parker (1961) that some photoautotrophic soil algae survive below the soil surface and some of which are adapted to survive heterotrophically below the soil surface. Such species are identified by culture of soils (El-Gamal et al., 2008) which also contributed to exploration of higher algal biodiversity in the current investigation.
Many species of algae observed in this study are economically highly significant. Stichococcus bacillaris that produce bio-diesel (Olivieri et al., 2011) was represented by two species in this soil. Similarly Scenedesmus that has high use in the cleanup of radioactive and metallic contaminants from soil and water through biotransformation (Diale et al., 2011) was also observed here. The potential of micro algae of soils include their utility in decomposition of organic residues and as indicators of pollution (Temraleeva et al., 2011), role in soil fertility and land reclamation process (Prasanna and Nayak, 2007) and prevention of soil erosion (Metting, 1981). Myrmecia, one of the genera of green-algae found in this soil, is a well known phycobiont of lichens Khaybullina et al. (2010). Many antibacterial, antifungal, antiviral, anti tumour and other bioactive compounds of many specific activities are extracted from micro algae (Singh et al., 2005) and hence the potential of the present inventory of soil algae is tremendous.
Zygnematales (Desmids) are the most dominant group of algae (42%) found in the Oxic Dystrustepts of Western Ghats in the current investigation. Desmids in general are cosmopolitan species of algae, considered as biological indicators of purity of water bodies and are found in environments of low electrical conductivity, low temperature, weakly acidic pH and limited nitrogen but a wide range of phosphorous (Stepankova et al., 2008). According to Ngearnpat et al. (2008), phosphorous deficiency leads to an obvious reduction in the growth of some desmids in peats. Acidification is known to be the cause of significant decrease of desmid diversity and changes in species composition (Stastny, 2010). The slightly acidic nature of the soils and non-degraded aspects of the soils with sufficient phosphorous content might be the reasons that supported a high diversity of desmids in these soils. Indirectly, the richness of desmid population in these soils indicates sufficient P in the soil which, may be harnessed into yield using appropriate mycorrhiza. Chlorococcales, a cosmopolitan group of algae (Coesel and Krienitz, 2007) was the second largest group of algae (19%) found in this soil. Growth of Chlorococcal green-algae are enhanced by the low pH in the soil (Metting, 1981). The high diversity of this algal group observed in these soils agrees with the observations of Neustupa (2001) that broad leaved forest soils contain high diversity of green-algae.
One of the most abundant genera found in this soil is Chlorella (relative abundance equals 63.33%) which is a ubiquitous species in wet tropics (Sharma et al., 2006). This is a well known metal binding genus used in bioremediation of contaminated soils (Yoshida et al., 2006). It is also considered as the most efficient algae in scavenging ammoniacal nitrogen and biodegradation of some carcinogenic dyes (El-Sheekh et al., 2009; Singh and Dhar, 2007). Reports of Lukesova (2001), Neustupa (2001) and Skaloud (2006) that Klebsormidium is a common alga of aero-terrestrial habitats is found true in the current zone of investigations as well. K. flaccidium is very tolerant to different types of stresses such as extreme dehydrations (Rindi, 2007). Khaybullina et al. (2010) also observed the species as common algae in the soils of forests. Stichococcus, considered as a common species in many soils (Lukesova, 2001; Neustupa, 2001; Zancan et al., 2006) is found common in these wet tropical soils as well. Chlorococcum, considered as common green-algae of temperate soils (Wehr and Sheath, 2003) is found common in this tropical soil as well. Chlorococcum was observed only in the in vitro culture of soils of forest, teak and rubber of the present study. Observation of Megharaj et al. (2000) that Chlorococcum as an indicator species of contaminated soils is not true as the genus was found common in soils from all the vegetations including that of virgin forest and also in the culture of all these soils. Scenedesmus, the most frequent algae in the liquid culture of all the soils of forest, rubber and teak of the present study, is one of the algae used for bio energy generation such as biodiesel and biohydrogen (Demirbas and Demirbas, 2011). Microspora is a frequent filamentous alga found in soils of all the three vegetations which is considered as a bioindicators (Novis, 2004).
Roya catholicatae and Cosmarium holmiense were species of green-algae found in the forest soils alone. Algae specific to both forest and rubber plantations were Chlamydomonas ovoidae, Chlamydomonas deasonii var. micro, Chlorococcum infusionum, Chlorococcum humicola, Coelastrella terrestris, Cylindrocystis crassa. All the species except Chlamydomonas ovoidae, Chlamydomonas deasonii var. micro, Chlorococcum infusionum, Chlorococcum humicolum, Coelastrella terrestris, Geminella interrupta, Cylindrocystis crassa, Roya catholicatae and Cosmarium holmiense found in teak plantations were found in the rubber plantations or forests as well.
CONCLUSION
Tropical Oxic Dystrustepts soils are quite rich in green-algal diversity which include hitherto unreported new species. Desmids are found to be the most dominant category of green-algae in the soil. Many of the green-algae are economically highly valuable species. Vegetation type was found to be the most significant factor that control green-algal diversity in these soils. Overall the report emphasizes the need of immediate accounting of the entire algal diversity of the Western Ghats of South India, a fast degrading natural soil ecosystem and a precious biodiversity hot-spot of the world.
ACKNOWLEDGMENTS
Financial support extended to the second author under FIP programme by the UGC, New Delhi, is gratefully acknowledged.
REFERENCES
- Bohlen, P.J., P.M. Groffman, C.T. Driscoll, T.J. Fahey and T.G. Siccama, 2001. Plant-soil-microbial interactions in a northern hardwood forest. Ecology, 82: 965-978.
Direct Link - Bongale, U.D., 1981. Abundance of algal species in relation to soil types and major crops in cultivated soils of Karnataka state (India). Plant Soil, 60: 289-293.
CrossRef - Chandran, P., S.K. Ray, T. Bhattacharyya, P. Srivastava, P. Krishnan and D.K. Pal, 2005. Lateritic soils of Kerala, India: Their mineralogy, genesis and taxonomy. Aust. J. Soil Res., 43: 839-852.
CrossRefDirect Link - Chun-Xiang, H.U. and L. Yong-Ding, 2003. Primary succession of algal community structure in Desert soil. Acta Bot. Sin., 45: 917-924.
Direct Link - Coesel, P.F.M. and L. Krienitz, 2007. Diversity and geographic distribution of desmids and other coccoid green algae. Biodivers. Conserv., 17: 381-392.
CrossRef - Demirbas, A. and M.F. Demirbas, 2011. Importance of algae oil as a source of biodiesel. Energy Convers. Manage., 52: 163-170.
CrossRefDirect Link - Dey, H.S., K. Tayung and A.K. Bastia, 2010. Occurrence of nitrogen-fixing cyanobacteria in local rice fields of Orissa, India. Ecoprint Int. J. Ecol., 17: 77-85.
CrossRefDirect Link - El-Gamal, A.D., N.A.E. Ghanem, E.Y. El-Ayouty and E.F. Shehata, 2008. Studies on soil algal flora in Kafr El-Sheikh governorate, Egypt. Egypt. J. Phycol., 9: 1-23.
Direct Link - El-Sheekh, M.M., M.M. Gharieb and G.W. Abou-El-Souod, 2009. Biodegradation of dyes by some green algae and cyanobacteria. Int. Biodeterioat. Biodegradat., 63: 609-704.
CrossRefDirect Link - Flint, E.A. and D.B. Williamson, 2010. Desmids (Chlorophyta), including two new species and three new varieties in two swamps, a lake and a tarn in the south Island of New Zealand. N. Z. J. Bot., 43: 285-300.
CrossRefDirect Link - Freystein, K., M. Salisch and W. Reisser, 2008. Algal biofilms on tree bark to monitor airborne pollutants. Biologia, 63: 866-872.
CrossRef - Fujita, Y. and H. Nakahara, 2006. Variations in micro algal structure in paddy soil in Osaka, Japan: Comparison between surface and subsurface soils. Limnology, 7: 83-91.
CrossRef - Hunt, M.E., G.L. Floyd and B.B. Stout, 1979. Soil algae in field and forest environments. Ecology, 60: 362-375.
Direct Link - Issa, O.M., C. Defarge, Y. Le Bissonnais, B. Marin and O. Duval et al., 2007. Effects of the inoculation of cyanobacteria on the microstructure and the structural stability of a tropical soil. Plant Soil, 290: 209-219.
CrossRef - Khaybullina, L.S., L.A. Gaysina, J.R. Johansen and M. Krautova, 2010. Examination of the terrestrial algae of the Great Smoky Mountains National Park, USA. Fottea, 10: 201-215.
Direct Link - Khondker, M., A. Alfasane and R.F. Bhuiyan, 2008. (Chlorophyta): A new record for Bangladesh. Bangladesh J. Bot., 37: 189-191.
Direct Link - Lemes-Da-Silva, N.M., L.H.Z. Branco and O. Nechi-Junior, 2010. Corticolous green algae from tropical forest remnants in the northwest region of Sao Paulo State, Brazil. Rev. Brasil Bot., 33: 215-226.
Direct Link - Lewis, L.A. and R.M. McCourt, 2004. Green algae and the origin of land plants. Am. J. Bot., 91: 1535-1556.
CrossRefDirect Link - Lukesova, A., 2001. Soil algae in brown Coal and Lignite Post-Mining Areas in Central Europe (Czech Republic and Germany). Rest. Ecol., 9: 341-350.
Direct Link - Mansour, H.A., 2008. Natural and propagated algal flora isolated from El-Fayum Governorate. Egypt. J. Bio., 10: 54-61.
Direct Link - Megharaj, M., D. Kantachote, I. Singleton and R. Naidu, 2000. Effects of long term contamination of DDT on soil microflora with special reference to soil algae and algal transformation of DDT. Environ. Pollut., 109: 35-42.
CrossRef - Nayak, S. and R. Prasanna, 2007. Soil pH and its role in cyanobacterial abundance and diversity in rice field soils. J. Applied Ecol. Environ. Res., 5: 103-113.
Direct Link - Neustupa, J., 2001. Soil algae from marlstone-substratum based biotopes in the nature park Dzban (Central Bohemia, Czech Republic) with special attention to the natural treeless localities. Algol. Stud., 101: 109-120.
Direct Link - Neustupa, J. and P. Skaloud, 2008. Diversity of subaerial algae and cyanobacteria on tree bark in tropical mountain habitats. Biologia, 63: 806-812.
CrossRefDirect Link - Neustupa, J. and P. Skaloud, 2010. Diversity of subaerial algae and cyanobacteria growing on bark and wood in the lowland tropical forests of Singapore. Plant Ecol. Evol., 143: 51-62.
CrossRefDirect Link - Ngearnpat, N., P.F.M. Coesel and Y. Peerapornpisal, 2008. Diversity of desmids in three Thai peat swamps. Biologia, 63: 901-906.
CrossRef - Novakovskaya, I.V. and E.N. Patova, 2008. Green algae in spruce forests in the north-east of European Russia. Biologia, 63: 836-842.
CrossRef - Novis, P.M., 2004. A taxonomic survey of Microspora (Chlorophyceae, Chlorophyta) in New Zealand. N. Z. J. Bot., 42: 153-165.
Direct Link - Ohtani, S., K. Suyama, H. Yamamoto, Y. Aridomi, R. Itoh and Y. Fukuoka, 2000. Distribution of soil algae at the monitoring sites in the vicinity of Syowa station between austral summers of 1992/1993 and 1997/1998. Polar Biosci., 13: 113-132.
Direct Link - Olivieri, G., A. Marzocchella, R. Andreozzi, G. Pinto and A. Pollio, 2011. Biodiesel production from Stichococcus strains at laboratory scale. J. Chem. Tech. Biotech., 86: 776-783.
CrossRef - Parker, B.C., 1961. Facultative heterotrophy in certain soil algae from the ecological viewpoint. Ecology, 42: 381-386.
CrossRefDirect Link - Patova, E.N. and M.F. Dorokhova, 2008. Green algae in tundra soils affected by coal mine pollutions. Biologia, 63: 831-835.
CrossRef - Pena-Castro, J.M., F. Martinez-Jeronimo, F. Esparza-Garcia and R.O. Canizares-Villanueva, 2004. Heavy metals removal by the microalga Scenedesmus incrassatulus in continuous cultures. Bioresour. Technol., 94: 219-222.
CrossRefDirect Link - Philipose, M.T., 1967. Chlorococcales. Indian Council of Agricultural Research, New Delhi, India, Pages: 365.
Direct Link - Prasanna, R. and S. Nayak, 2007. Influence of diverse rice soil ecologies on cyanobacterial diversity and abundance. Wetlands Ecol. Manage., 15: 127-134.
CrossRef - Ray, J.G. and T.T. Binoy, 2012. Fertility characteristics of oxic dystrustepts under natural forest, rubber and teak plantations in different seasons, Kerala, South India. Commun. Soil Sci. Plan. Anal., 43: 2247-2261.
CrossRefDirect Link - Rindi, F., H.A. Allali, D.W. Lam and J.M. Lopez-Bautista, 2009. An Overview of the Biodiversity and Biogeography of Terrestrial Green Algae. In: Biodiversity Hotspots, Rescigno, V. and S. Maletta (Eds.). Chapter 3, Nova Science Publishers Inc., Hauppauge, NY., USA., ISBN: 9781608764587, pp: 105-122.
- Sharma, N.K., S. Singh and A.K. Rai, 2006. Diversity and seasonal variation of viable algal particles in the atmosphere of a subtropical city in India. Environ. Res., 102: 252-259.
Direct Link - Shimmel, S.M. and W.M. Darley, 1985. Productivity and density of soil algae in an agricultural system. Ecology, 66: 1439-1447.
Direct Link - Singh, A.P. and B.R. Chaudhary, 2011. Allelopathic potential of algal weed Pithophora oedogonia (Mont.) ittrock on the germination and seedling growth of Oryza sativa L. Bot. Res. Int., 4: 36-40.
Direct Link - Singh, N.K. and D.W. Dhar, 2007. Nitrogen and phosphorous scavenging potential in microalgae. Indian J. Biotechnol., 6: 52-56.
Direct Link - Singh, P.K., S. Kishore, J. Prakash, S.K. Singh and M. Shukla, 2008. Cyanophycean algae inhabiting sodic soil exhibit diverse morphology: An adaptation to high exchangeable sodium. Ecoprint, 15: 15-21.
Direct Link - Singh, S., B.N. Kate and U.C. Banerjee, 2005. Bioactive compounds from cyanobacteria and micro algae: An overview. Crit. Rev. Biotechnol., 25: 73-95.
PubMed - Skaloud, P., 2006. Variation and taxonomic significance of some morphological features in European strains of Klebsormidium (Klebsormidiophyceae, Streptophyta). Nova Hedwigia, 83: 533-550.
CrossRefDirect Link - Skaloud, P., 2009. Species composition and diversity of aero-terrestrial algae and cyanobacteria of the Borec Hill ventaroles. Fottea, 9: 65-80.
Direct Link - Soare, L.C. and C.M. Dobrescu, 2010. Preliminary data on edaphic algae in the city of Pitesti (Romania). Fascicula Biol., 17: 186-189.
Direct Link - Starks, T.L. and L.E. Shubert, 1982. Colonization and succession of algae and soil-algal interactions associated with disturbed areas. J. Phycol., 18: 99-107.
CrossRef - Stastny, J., 2010. Desmids (Conjugatophyceae, Viridiplantae) from the Czech Republic: New and rare taxa, distribution, ecology. Fottea, 10: 1-74.
Direct Link - Stepankova, J., J. Vavruskova, P. Hasler, P. Mazalova and A. Poulickova, 2008. Diversity and ecology of desmids of peat bogs in the Jizerske hory Mts. Biologia, 63: 895-900.
CrossRefDirect Link - Temraleeva, A.D., D.L. Pinskii, E.N. Patova and E.V. Spirina, 2011. The use of algae-cyanobacterial communities for the assessment of lead pollution of gray forest soils. Eurasian Soil Sci., 44: 326-331.
CrossRef - Tirkey, J. and S.P. Adhikary, 2005. Cyanobacteria in biological soil crusts of India. Curr. Sci., 89: 515-521.
Direct Link - Tschaikner, A., E. Ingolic, M.P. Stoyneva and G. Gartner, 2007. Autosporulation in the soil alga Coelastrella terrestris (Chlorophyta, Scenedesmaceae, Scenedesmoideae). Phytol. Balcanica, 13: 29-34.
Direct Link - Yoshida, N., R. Ikeda and T. Okuno, 2006. Identification and characterization of heavy metal-resistant unicellular algae isolated from soil and its potential for phytoremediation. Bioresour. Technol., 97: 1843-1849.
CrossRef - Zancan, S., R. Trevisan and M.G. Paoletti, 2006. Soil algae composition under different agro-ecosystem in North-Eastern Italy. Agric. Ecosyst. Environ., 112: 1-12.
CrossRef - Zarina, A., Masu-ul-Hasan and M. Shameel, 2006. Taxonomic study of the class Ulvophyceae (Chlorophyta) from certain areas of the Punjab, Pakistan. Proc. Pak. Acad. Sci., 43: 229-240.
Direct Link - Zidarova, R.P., 2008. Algae from Livingston Island (S Shetland Islands): A checklist. Phytol. Balcanica, 14: 19-35.
Direct Link - Broady, P.A., 1979. Qualitative and quantitative observations on green and yellow green algae in some English soils. Eur. J. Phycol., 14: 151-160.
CrossRefDirect Link