
Sumpam Tangjang
Department of Botany, Rajiv Gandhi University, Rono Hills, Itanagar-791111, India
Kusum Arunachalam
Department of Forestry, NERIST, Nirjuli-791109, India
Ayyanadar Arunachalam
Department of Forestry, NERIST, Nirjuli-791109, India
Awadhesh Kumar Shukla
Department of Botany, Rajiv Gandhi University, Rono Hills, Itanagar-791111, India
Research Journal of Soil Biology
Year: 2009 | Volume: 1 | Issue: 1 | Page No.: 1-7







ABSTRACT
Seasonal and depth wise variation in bacterial and fungal populations was studied in three arecanut-based traditional agroforestry systems practiced by two ethnic communities viz., the Kalitas and Nyishis. Arecanut palm was abundantly found in Harmutty and this site also recorded greater species diversity. Soil was sandy loam in all sites. Soil organic C, total N and available P decreased with increasing soil depth. Bacterial population was highest during spring and that of fungi during autumn. Nonetheless, the highest microbial counts were recorded in the topsoil (0-10 cm) layer except during the rainy season when the population was greater in the subsurface (10-20 cm) layer. Altogether, 29 soil micro-fungal forms were recorded from three sites. Aspergillus and Penicillium were the abundant genera in the sites. Soil organic C and total N concentration had correlation with the microbial colony forming units. It was noticed that plant residues, added organic matter, vegetation, plant species composition and soil mineral nutrients altered the microbial population as well as their species composition under traditional agroforestry system.
PDF Abstract XML References Citation
How to cite this article
Sumpam Tangjang, Kusum Arunachalam, Ayyanadar Arunachalam and Awadhesh Kumar Shukla, 2009. Microbial Population Dynamics of Soil Under Traditional Agroforestry Systems in Northeast India. Research Journal of Soil Biology, 1: 1-7.
DOI: 10.3923/rjsb.2009.1.7
URL: https://scialert.net/abstract/?doi=rjsb.2009.1.7
DOI: 10.3923/rjsb.2009.1.7
URL: https://scialert.net/abstract/?doi=rjsb.2009.1.7
INTRODUCTION
Soil is a complex ecosystem, delimited by physico-chemical parameters that hold enormous numbers of diverse living organisms. Nevertheless, microbes are the least unstated mechanism of soil by both agronomists and the soil practitioners on the farm. Several soil organisms offer benefits to crop growing in an ecosystem, but are not well understood. The soil microbes decompose the plant and animal residues entering the soil and convert them into soil organic matter, which influences on soil physical, chemical and biological properties and on creating a complimentary medium for biological reactions and life support in the soil environment. Nonetheless, enhanced site-specific diversity typically results in higher levels of belowground microbial diversity and production (Olson et al., 2000).
Large quantities of readily decomposable organic matter are added to agricultural soils every year as crop residues or animal wastes and have a significant outcome on soil microbial commotion. The plant species growing on the soil also equally influence the population and species composition of the soil fungi (Hackl et al., 2000). Fungi and bacteria play a focal role in nutrient cycling by regulating soil biological activity (Arunachalam et al., 1997). However, the rate at which organic matter is decomposed by the microbes is interrelated to the chemical composition of the substrate as well as environmental conditions.
Multi-strata agroforestry systems are extensive in the humid and sub-humid tropics of the world, counting lowland and mountainous regions (Nair and Muschler, 1993) that posses major challenge for
study of plant-soil interactions. There have been a number of studies on the distribution of forest soil microfungi in terrestrial ecosystem. Some studies dealt with the influence of plant community (Chung et al., 2007; Carney and Matson, 2006), some with depth effects (Arunachalam et al., 1997) and others attempted to examine seasonal trends (Kennedy et al., 2005). Conversely, an integrated approach to the dynamics of microbial population in the traditional agroforestry system in and around Arunachal Pradesh is deficient, although such a challenge may give an insight into the microbial population dynamics as influenced by soil organic matter and nutrient build up. This study deals with the seasonal and depth wise variations in soil bacterial and fungal populations in relation to the micro-environmental and soil nutrient variability in one of the least studied traditional agroforestry systems in the northeast India. To go about the aforementioned approach, data on the seasonal and depth wise variations in bacterial and fungal populations in traditional agroforestry have been consummated and discussed in this paper to derive some conclusions regarding the influence in relation to micro-environmental and soil nutrient variability on the microbial population dynamics.
MATERIALS AND METHODS
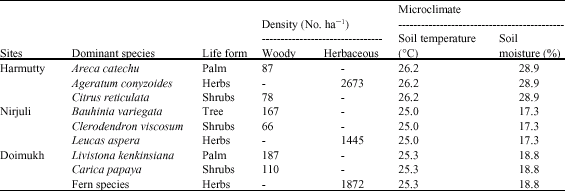
Study Sites: The study was conducted during 2003 January to 2004 November in traditional agroforestry systems of two villages in Arunachal Pradesh (27°60´N latitude and 94°21´E longitude) namely, Doimukh and Nirjuli (126 m above MSL) of Papum Pare district and Harmutty village (120 m above MSL) of North Lakhimpur district of Assam (26°46´N latitude and 93°50´E longitude), bordering Arunachal Pradesh, India. The Kalitas dominated Harmutty while Nirjuli and Doimukh were inhabited by the Nyishis, one of the major tribes of this area. Three traditional agroforestry systems were selected in each of the sites. The average area of the agroforestry plot varied between 200-400 m2 in Nirjuli and Doimukh and 320-490 m2 in Harmutty. The agroforestry were multi-storied consisting of trees, shrubs, herbs and climbers (Table 1) and consisted of species that are chosen according to the requirements of the household (Tangjang et al., 2004).
The sites are characterized by a climate with most rainfall occurring during the summer months (May-July) with relatively a little or scanty during the winter months. Total annual rainfall is typically 1100-1600 mm in all the sites and ambient temperatures average from minimum 12°C to a maximum of 37°C in all the study sites experiencing a humid tropical climate. Soils at the site were sandy loam to loamy sand. Nirjuli and Doimukh area are composed of newer alluvium (newer terrace deposits) represented by valley fill deposits comprising of sediments, while Harmutty area is composed of alluvium belonging to Pleistocene and recent times (Kumar, 1997). Over all, soil was sandy loam in all sites and slightly acidic (Table 2).
| Table 1: | Vegetation and microclimate in the three agroforestry systems (n = 12) |
 | |
| Table 2: | Soil properties in the traditional agroforestry systems |
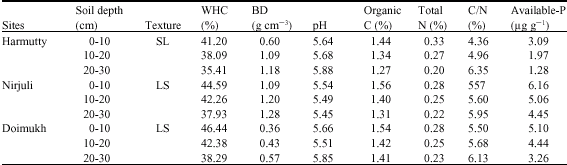 | |
| SL: Sandy Loam; LS: Loamy Sand; WHC: Water Holding Capacity; BD: Bulk Density | |
Soil Sampling and Analysis: Soil from three depths (viz., 0-10, 10-20 and 20-30 cm) were sampled during January, March, May and November that are the representative months of winter, spring, rainy and autumn seasons, during when the sampling was done. In each stand, ten replicates of soil samples were collected aseptically in sterilized polythene bags using a steel corer (6.5 cm inner diameter) and were used for the isolation of bacteria and fungi within 24 h. The remaining soil samples were air-dried and used for the determination of physico-chemical properties (Anderson and Ingram, 1993). Soil moisture content was determined gravimetrically by oven drying 10 g of fresh sieved soil for 24 h at 105°C.
Isolation of Bacterial and Fungal Population: Soil bacterial population was estimated by Waksman`s (1952) method using the nutrient agar medium at 105 dilutions. Fungal population was estimated by dilution plate method (Johnson and Curl 1972) using Martin`s Rose Bengal agar medium at 103 dilutions in water. The inoculated Petri-dishes were incubated at 30±1°C for 24 h and 25±1°C for 5 days for bacteria and fungi respectively. To calculate the populations of bacteria and fungi, colonies developed on Petri dishes were counted with the help of digital colony counter and expressed as number of colony forming units (cfu) g-1 dry soil. Representative isolates of fungi were identified under microscope with the help of standard manuals (Domsch et al., 1980; Barnett and Hunter, 1972).
RESULTS
Bacterial and fungal counts were greater in the surface (0-10 cm) layer of the soil as compared to others (Table 3). Maximum bacterial population was recorded in spring season in all the sites and minimum during winter. In contrast, fungal counts were higher during autumn in all the sites. During rainy season, maximum microbial count was, however, recorded in the subsoil (10-20 cm) layer. Both bacterial and fungal cfus were more in Harmutty than in Nirjuli and Doimukh sites. Quantitatively, bacterial counts were always high as compared to fungal population in all the sites. In general, similar trend was observed in the distribution of soil microorganisms in all the sites.
Altogether, 29 forms of fungi were isolated in the three sites. List of isolated fungi during the study is shown in Table 4. The relative proportion of different fungal taxonomic groups was almost identical in all the three sites. Penicillium and Aspergillus were the most abundant group of species in all the three study sites. However, some species were restricted to a particular site. Aspergillus fumigatus, Trichoderma viride and Syncephalus sp. were exclusively found in Nirjuli, while Trichoderma sp. was encountered only in Harmutty. Nevertheless, the number of fungal genera encountered in the three sites was similar.
| Table 3: | Seasonal variations in bacterial cfu (x105 g-1 dry soil) and fungal cfu (x103 g-1 dry soil) at three soil depths |
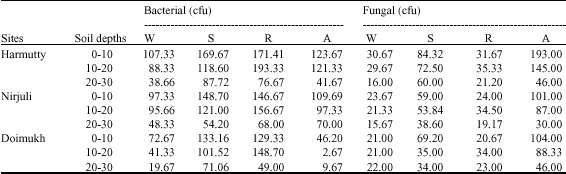 | |
| W: Winter; S: Summer; R: Spring; A: Autumn | |
| Table 4: | Microfungi isolated from three soil depths in the three traditional agroforestry systems |
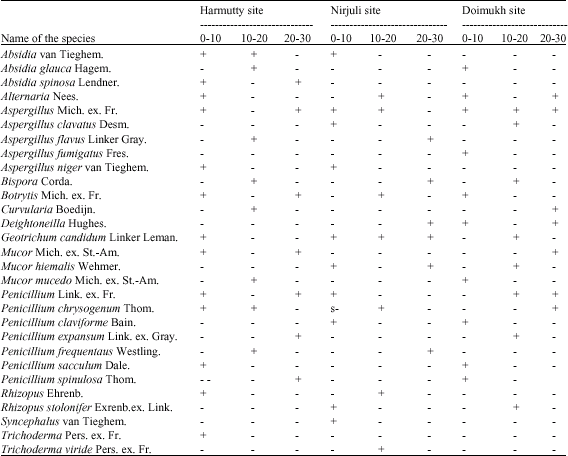 | |
| - and +: indicates absence and presence, respectively | |
| Table 5: | Correlation coefficient (R) for the relationships between bacterial and fungal cfu and soil physico-chemical properties |
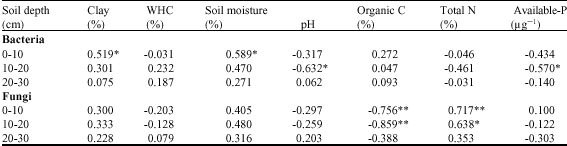 | |
| df = 17, *: p<0.05; p<0.01 | |
Among the soil physico-chemical properties, clay content and soil moisture show significant positive correlations with bacterial and fungal cfu (<0.05); water holding capacity and soil pH showed negative correlations, while organic C and total N showed positive correlations for bacteria and negative correlations with that of fungi (Table 5).
DISCUSSION
Generally, topsoil contains high organic matter, which in the presence of adequate moisture supply is acted upon by the microorganisms to decompose the complex organic residues into simpler forms; hence, microbial counts are generally higher in the surface soil layer (Shamir and Steinberger, 2007) as compared to the lower depths. However, the distribution of soil microbial population is determined by a number of environmental factors like pH, moisture content and soil organic matter (Kennedy et al., 2005). Higher bacterial population in the topsoil (0-10 cm) layer during spring season in present study is in agreement with the observation of Jha et al. (1992a) who recorded higher populations during spring and post-rainy seasons. However, peak in bacterial population was recorded during rainy season may be attributed to favorable soil moisture and temperature conditions that coincide with greater microbial activity and decomposition. On the other hand, minimum population counts during winter in the present study sites may be due to low ambient temperature and greater physiological water stress which are otherwise critical for the growth and activity of microbes. Higher fungal population during autumn supported the findings of other workers (Arunachalam et al., 1997), which perhaps is due to prevailing favorable moisture and temperature setting during the post-monsoon periods and also that the litter and other plant residues are decomposed faster during rainy season and sufficient soil organic matter and humus accumulates that may have enhanced the colonization of the soil microbes in subsequent period.
Maximum population in the subsoil (10-20 cm) layer during rainy season in all the three sites studied corroborates to that of Classen et al. (2007) who pointed out that during hot summer months, the sub-layer of soil occasionally harbors more fungal populations caused by temperature and moisture regimes than the topsoil layer. Higher rate of infiltration in the loamy sand may also in part have contributed to this phenomenon. However, Shukla et al. (1989) and Arunachalam et al. (1997) found negligible differences in fungal population across depths. Low fungal population during rainy season (Table 3) in the topsoil may partly be linked to the run-off losses of fungal propagules along with the plant materials from the hill slope due to heavy rainfall in the region (Shukla et al., 1989). Further, overall reduction in microbial population in the lower soil depths was attributed to fewer amounts of minerals, low oxygen content and increased carbon-dioxide concentration (Shukla et al., 1989). Consequently, fungal flora noted in the deeper depths of soil was significantly low in all the three sites. During winter, low moisture content might have slowed down microbial activity and organic matter decomposition and thus resulted in a low microbial population.
A total of 29 soil microfungi were isolated (Table 4). However, only a few fungal species were found to be dominant and basically no marked variations in the composition of species were noticed in different seasons of the year across the sites. The species like Aspergillus, Botrytis, Geotrichum, Penicillium and Rhizopus were common to all sites. Some fungal species encountered were rare and restricted to particular site. Dominance of the genus Penicillium and Aspergillus in the present study sites may be due to their greater rate of spore production and dispersal and partly due to their resistance over extreme environmental conditions (Schimel, 1995). The fungal species richness recorded in the present study was higher than those (13 species) reported from subtropical humid forest soils in north-east India (Arunachalam et al., 1997) and 26, 21 and 27 species, respectively from soils of valley land, terrace and slopes in this region (Shukla and Mishra, 1992). Conversely, the present species number was slightly lower than those reported 33 fungal species from subtropical humid forest by Arunachalam et al. (1997) and was much lower as compared to 41 fungal species isolated from South Dakota grassland soil (Dennis and Christensen, 1981). Presence of one or a few dominant species in the present study was however in agreement with that of Jha et al. (1992b) who pointed out that for a given community, only a few species are numerically predominant and may strongly affect the environmental conditions for the other.
The topsoil layer (0-10 cm) had 22 species which was greater than the subsoil layers (10-20 and 20-30 cm). Dkhar (1983) suggested that fungi grow slowly in the deeper soil layers due to shortage of mineral nutrients and compaction of soil along depth. The rate of change in fungal population was attributed to the type of vegetation growing on a particular area (Entry and Emmingham, 1996), variation in physico-chemical characteristics of the soil and environmental complex of the locality (Bossio et al., 2005). Higher counts of bacterial and fungal population in Harmutty may be attributed to the dense growth of plants and greater availability of nutrients on account of greater accumulation of litter and may also be due to spreading of other biodegradable domestic wastes into the system by the traditional farmers. Contrarily, low microbial population in Nirjuli and Doimukh may be because addition of plant remains and household waste in these sites was comparatively poor that also resulted in low organic matter content in the soil. Many workers have also recorded a correlation between fungal species composition and the species composition of the aboveground vegetation (Chung et al., 2007). Ground floor of Harmutty site of present study was also dominated by fern species.
CONCLUSION
Over all, the present study concludes that the population counts of bacteria and fungi in the soils of traditional agroforestry is influenced by vegetation, density and species composition. However, the role of macro and micro-climatic seasonality and soil nutrient status can not be completely ruled out. It is also understood that the quality of plant residues accumulating in these homestead agroforestry are furthermore important and may play a vital role in soil nutrient management within the system through microbial decomposition however.
ACKNOWLEDGMENTS
We thank the villagers of Harmutty, Nirjuli and Doimukh for their cooperation during the field study. The authors are also thankful to Mr. Moharam Ingti, Laboratory Assistant for helping in the laboratory studies. This study is part of a research project funded by the Indian Council of Agricultural Research (ICAR), New Delhi. The first author also thanks the Council of Scientific and Industrial Research (CSIR), New Delhi for awarding Junior Research Fellowship.
REFERENCES
- Bossio, D.A., M.S. Girvan, L. Verchot, J. Bullimore, T. Borelli, A. Albrecht, K.M. Scow, A.S. Ball, J.N. Pretty and A.M. Osborn, 2005. Soil microbial community response to land use change in an agricultural landscape of Western Kenya. Microb. Ecol., 49: 50-62.
CrossRefDirect Link - Carney, K.M. and P.A. Matson, 2006. The influence of tropical plant diversity and composition on soil microbial communities. Microb. Ecol., 52: 226-238.
PubMed - Chung, H., D.R. Zak, P.B. Reich and D.S. Ellsworth, 2007. Plant species richness, elevated CO2 and atmospheric nitrogen deposition alter soil microbial community composition and function. Global Change Biol., 13: 980-989.
CrossRefDirect Link - Classen, A.T., S.T. Overby, S.C. Hart, G.W. Koch and T.G. Whitham, 2007. Season mediates herbivore effects on litter and soil microbial abundance and activity in a semi-arid woodland. Plant Soil, 295: 217-227.
Direct Link - Dennis, C. and M. Christensen, 1981. The soil microfungal community of a South Dakota grassland. Can. J. Bot., 59: 1950-1960.
CrossRefDirect Link - Domsch, K.H., W. Gams and T.H. Anderson, 1980. Compendium of Soil Fungi. Academic Press, London, United Kingdom, ISBN: 9780122204012, Pages: 1264.
Direct Link - Entry, J.A. and W.H. Emmingham, 1996. Influence of vegetation on microbial degradation of atrazine and 2,4-dichlorophenoxyacetic acid in riparian soils. Can. J. Soil Sci., 76: 101-106.
CrossRefDirect Link - Hackl, E., G. Bachmann and S. Zechmeister-Boltenstern, 2000. Soil microbial biomass and rhizosphere effects in natural forest stands. Phyton, 40: 83-90.
Direct Link - Jha, D.K., G.D. Sharma and R.R. Mishra, 1992. Soil microbial population numbers and enzyme activities in relation to altitude and forest degradation. Soil Biol. Biochem., 24: 761-767.
CrossRefDirect Link - Jha, D.K., G.D. Sharma and R.R. Mishra, 1992. Ecology of soil microflora and mycorrhizal symbionts in degraded forests at two altitudes. Biol. Fertil. Soils, 12: 272-278.
CrossRefDirect Link - Kennedy, N.M., D.E. Gleeson, J. Connolly and N.J.W. Clipson, 2005. Seasonal and management influences on bacterial community structure in an upland grassland soil. FEMS Microb. Ecol., 53: 329-337.
CrossRefDirect Link - Shamir, I. and Y. Steinberger, 2007. Vertical distribution and activity of soil microbial population in a sandy desert ecosystem. Microb. Ecol., 53: 340-347.
Direct Link - Shukla, A.K. and R.R. Mishra, 1992. Influence of soil management systems on the microfungal communities of potato field. Cryptogamie Mycol., 13: 135-144.
Direct Link - Tangjang, S., S. Deb, A. Arunachalam, U. Melkania, K. Arunachalam, K. Shrivastava and S. Brahma, 2004. Tribal communities and vegetational characteristics in traditional agroforestry systems of Northeast India. Indian J. Agrofor., 63: 73-80.
Direct Link