
Meeta Jain
School of Biochemistry, Devi Ahilya Vishwavidyalaya, Takshashila Campus, Khandwa Road, 452 017 Indore (M.P.), India
LiveDNA: 91.14319
Mahija Panwar
School of Biochemistry, Devi Ahilya Vishwavidyalaya, Takshashila Campus, Khandwa Road, 452 017 Indore (M.P.), India
Rekha Gadre
School of Biochemistry, Devi Ahilya Vishwavidyalaya, Takshashila Campus, Khandwa Road, 452 017 Indore (M.P.), India
Research Journal of Phytochemistry
Year: 2017 | Volume: 11 | Issue: 3 | Page No.: 111-117







ABSTRACT
Background and Objective: Chlorophylls are essential tetrapyrroles responsible for harvesting solar energy, charge separation and electron transport in photosynthesis. δ-Aminolevulinic acid (ALA) is the common precursor of tetrapyrroles and its synthesis is a key regulatory step in chlorophyll biosynthetic pathway. Selenium is an essential micronutrient for animals and humans but its nutritional requirement in plants is controversial. The effect of Se on ALA synthesis has not been worked out. Thus, the present study was aimed to investigate the effect of Se on ALA synthesizing activity in etiolated maize leaf segments during greening. Materials and Methods: ALA content was estimated by modified Ehrlich’s reagent. ALA content measured after treatment with levulinic acid represents ALA formation. Total chlorophylls were extracted in 80% acetone and then quantitated spectrophotometrically. Student’s t-test was applied to test the significance of difference at probability levels p<0.05, p<0.01and p<0.001. Results: Supply of 0.1 and 0.5 mM Na2SeO3 decreased the ALA content in greening maize leaf segments; however, 0.05 mM Se slightly enhanced its contents. Decline in chlorophyll content was more substantial than decrease in ALA content and carotenoids. Concentration dependent decrease in ALA accumulation was observed with increasing concentration of selenite in light and dark. Substantial difference in ALA accumulation (L-D) was noticed at 0.5 mM Se. Amongst the precursors tested, glutamate and succinate caused more ALA accumulation in -Se tissue in light, while in +Se treated tissue, the increase was prominent by glycine and succinate. When leaf segments were incubated with levulinic acid in light, the ALA formation was increased by GSH in absence of Se; however, it was increased by NADP+ in presence of Se. In dark, GSH stimulated ALA formation, both in the absence and presence of Se. Conclusion: More prominent decrease in total chlorophylls than ALA content/formation with the supply of Se indicates the involvement of other steps of chlorophyll biosynthesis in addition to ALA synthesis, being affected by Se. Substantial decrease in L-D value at higher concentrations of Se suggests that chloroplastic ALA synthesizing activity is more sensitive towards Se.
PDF Abstract XML References Citation
Received: June 29, 2017;
Accepted: August 01, 2017;
Published: September 23, 2017
How to cite this article
Meeta Jain, Mahija Panwar and Rekha Gadre, 2017. Influence of Selenium Supplementation on δ-Aminolevulinic Acid Formation in Greening Maize Leaf Segments. Research Journal of Phytochemistry, 11: 111-117.
URL: https://scialert.net/abstract/?doi=rjphyto.2017.111.117
URL: https://scialert.net/abstract/?doi=rjphyto.2017.111.117
INTRODUCTION
Plants synthesize a wide variety of tetrapyrroles including chlorophylls, hemes, siroheme and bilins, which participate in many cellular processes. Chlorophyll biosynthesis is subjected to regulation mainly at three steps: The synthesis of δ-Aminolevulinic acid (ALA), Mg chelation and phototransformation of protochlorophyllide. Delta-aminolevulinic acid is the universal precursor of tetrapyrroles and all carbon and nitrogen atoms of tetrapyrroles are derived from it1. ALA is synthesized by two different pathways. In fungi, some bacteria and animals, a pyridoxal phosphate dependent enzyme ALA synthase catalyses the condensation of glycine and succinyl CoA to give ALA through Shemin pathway2. In contrast, in majority of bacteria, higher plants and algae, the intact 5-carbon skeleton of glutamate is converted into ALA by a multienzyme system through Beale pathway1. Biosynthesis begins with the activation of glutamate by glutamyl-t RNA synthetase to form glutamyl-t RNA. The activated glutamate is then reduced to Glutamate Semi Aldehyde (GSA) by NADPH dependant glutamyl-t RNA reductase. Finally, conversion of GSA to ALA is catalysed by GSA aminotransferase enzyme, requiring pyridoxal phosphate or pyridoxamine phosphate as a cofactor. ALA formation has been shown to be sharply decreased by various stresses3,4. Further, exogenous application of ALA has been reported to regulate antioxidant enzyme activities and thereby increased resistance of plants to different stresses, such as, cold5, chilling6, low irradiance7 and salinity8. Furthermore, a variety of medical applications including tumour diagnosis and cancer treatment for ALA have also been reported9. Hence, using levulinic acid (LA), a competitive inhibitor of ALA dehydratase enzyme, microbial production of ALA in photosynthetic bacteria has been identified10.
Selenium is an essential micronutrient for animals and humans nutrition because it is an integral part of glutathione peroxidase, a seleno enzyme that prevents oxidative damage to body tissues11. The effect of Se in plants is not well recognized and its role in the plant metabolism is still controversial. Several studies demonstrate that at low concentrations it may exert diverse beneficial effects, including growth promoting activity12, protection against abiotic stresses, such as, low temperature13, drought stress14, aluminium toxicity15, arsenic toxicity16 and salt stress17. Looking to the mitigating effects of Se against heavy metals, approach of Se-fertilization method has been used against cadmium18. An elevated level of Se is toxic to most plants, due to its non-specific incorporation into sulphur of amino acids. Seleno-amino acids incorporated into protein cause replacement of S-S bonds by the less stable Se-Se bonds, thus change the biological activity of proteins19. On the basis of the food chain, plants collect Se from the soil and provide it to higher trophic levels. Using this principle, plants have been used to remove Se from natural or polluted Se-rich areas by the process called phytoremediation and also as a food source to alleviate Se deficiency in humans or animals by bio fortification20. Decrease in total chlorophylls and ALA contents by arsenic has been reported in greening maize leaf segments21. However, the effect of Se on δ-ALA synthesis has not been worked out. This investigation will be helpful in understanding the mechanistic details of the toxic effect of Se on a vital physiological process, chlorophyll biosynthesis in maize leaves and further in developing Se resistant crops. The present study was carried out to investigate the effect of Se on δ-Aminolevulinic acid synthesis, a key regulatory step in chlorophyll biosynthesis, in etiolated maize leaf segments during greening.
MATERIALS AND METHODS
This investigation was performed as factorial experiment under Completely Randomized Design (CRD) with six replications during 2015-16 at School of Biochemistry, Devi Ahilya University, Indore. All the chemicals and solutions used in the study were of analytical grade.
Seed material, plant culture and tissue treatment: Seeds of Zea mays L.cv. Ganga Safed-2 were surface sterilized with 0.1% HgCl2 for 1-2 min and washed with tap water and then distilled water. Seeds were sown in plastic buckets containing acid washed sand, at 26±2°C for 7-8 days in continuous darkness. They were watered with ½ strength Hoagland’s solution without nitrogen. For various experiments the first and fully elongated leaves were taken. Leaves were cut into small segments (0.5 cm2) and were treated with varying concentrations of Se; 0.05 mM, 0.1 mM and 0.5 mM in continuous light intensity of 40 W m–1 at 25± 2°C for 24 h in light chamber. One fourth strength Hoagland solution was used as a control. At the end of treatment, leaf segments were thoroughly washed with distilled water prior to analysis.
Estimations: For estimating ALA content, treated leaf segments were hand homogenized in 1.0 M Na acetate buffer, pH 4.6. The extract was centrifuged at 10,000 rpm at 4°C for 20 min and the supernatant was used to quantify ALA content after condensing into porphobilinogen using ethylacetoacetate. Porphobilinogen was estimated using modified Ehrlich reagent according to the method of Tewari and Tripathy22. For determining ALA synthesizing activity, treated leaf segments were pre-incubated in 50 mM phosphate buffer containing 60 mM levulinic acid (pH 6.0) for 3 h in light and darkness, followed by estimation of ALA content. Aminolevulinic acid was extracted and estimated by the method of Tewari and Tripathy22. Total chlorophylls were extracted in 80% acetone and quantitated according to the method of Strain and Svec23.
Statistical analysis: The data presented in the text are the average values of at least three replicate experiments with ±standard error. Student’s t-test was applied to test the significance difference at probability levels p<0.05, p<0.01 and p<0.00124.
RESULTS
ALA content, total chlorophylls and carotenoids: When leaf segments were treated with 0.05 mM Na2SeO3, the ALA content increased marginally, however, 0.1 and 0.5 mM Na2SeO3 decreased the ALA content of the leaf tissue. (Table 1). Chlorophyll and carotenoid contents also decreased with the supply of 0.5 mM Se, where decline was found to be more severe for chlorophyll than ALA and carotenoid contents (Table 1).
ALA formation in light and dark: Incubation of maize leaf segments with phosphate buffer containing levulinic acid, in light and dark after treatment with sodium selenite decreased the ALA content in a concentration dependent manner (Table 2). Further, decrease in ALA content was almost to the same extent in both light and dark at each concentration of sodium selenite. There was a slight increase in the L-D value at 0.05 mM Na2SeO3; however, it decreased considerably at 0.1 and 0.5 mM concentration (Table 2).
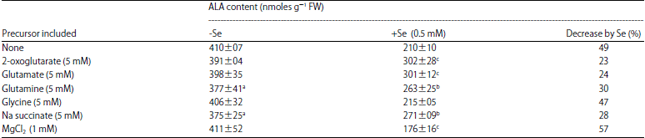
Effect of supply of precursors of chlorophyll on ALA content in the absence and presence of Na2SeO3: Treatment of excised maize leaf segments with 5.0 mM each of 2-oxoglutarate, glutamate, glutamine and Na succinate increased the ALA content in the presence of 0.5 mM Na2SeO3, however, almost same content of ALA was obtained in the leaf tissue without selenite treatment. (Table 3). In the presence of 1 mM MgCl2, the ALA content decreased in the presence of sodium selenite. Inclusion of glycine did not affect the ALA content in both the absence as well as in presence of Na2SeO3 (Table 3).
Effect of precursors of chlorophyll on ALA formation in light using Se treated and untreated leaf segments: For determining ALA formation activity, treated leaf segments were incubated in 50 mM phosphate buffer containing 60 mM levulinic acid (pH 6.0) for 3 h in light and darkness. It was observed that treatment with 5.0 mM each of 2-oxoglutarate and glycine as well as 1.0 mM MgCl2, decreased the ALA content in light, where the effect was more substantial with glycine (Table 4). The effect of these precursors was found to be different in the presence of sodium selenite. 2-Oxoglutarate decreased the ALA content marginally, while glycine and MgCl2 increased it, the increase being almost double by glycine and slightly by MgCl2. Due to these effects, percentage decrease by Se remains unaffected with 2-oxoglutarate, reduced by MgCl2 and greatly reduced by glycine. Supply of 5 mM each of glutamate and Na succinate increased the ALA content in light, in the absence of Se, this increase being more substantial with glutamate. These precursors increased the ALA content in the presence of sodium selenite also, however, the increase was more pronounced for Na succinate than glutamate (Table 4). With the inclusion of glutamine, ALA content remained same in light, in the absence of Se, while, there was slight increase in the presence of Se.
Effect of precursors of chlorophyll on ALA formation in dark using Se treated and untreated leaf segments: Treatment of maize leaf segments with 5 mM each of 2-oxoglutarate, glutamine and glycine and 1 mM MgCl2, in the absence of Na2SeO3, followed by incubation with phosphate buffer containing 60 mM levulinic acid in dark decreased the ALA content (Table 5). However, increase was obtained with these precursors in the presence of Na2SeO3 and due to these effects percentage decrease by Se was reduced. Supply of 5 mM each of glutamate and Na succinate increased the ALA content in the absence of selenite in dark, the increase being more pronounced with latter than the former. However, reverse trend of increase was found with these precursors in the presence of sodium selenite in dark, i.e., more pronounced increase with glutamate than Na succinate (Table 5).
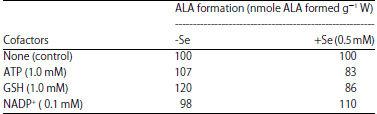
Effect of supply of cofactors on ALA formation in light and dark: Addition of 1 mM ATP and 0.1 mM NADP+ along with levulinic acid in the incubation mixture did not affect the ALA content in light in -Se treated leaf segments, however, 1 mM glutathione increased it (Table 6). In the leaf segments treated with Na2SeO3, the ALA content increased with NADP+ inclusion in the incubation mixture in light, while, it decreased with ATP and GSH (Table 6).
| Table 1: | Effect of supplementation of Na2SeO3 on ALA content, total chlorophylls and carotenoids in excised maize leaf segments during greening |
 | |
| Table 2: | Effect of supplementation of Na2SeO3 on ALA formation in light and dark |
 | |
±: Standard error, Values in round brackets are relative values in comparison to control to represent degree of inhibition. Level of significance: p<0.01b, p<0.001c | |
| Table 3: | Effect of supply of precursors of chlorophyll on ALA content in the absence and presence of Na2SeO3 in excised maize leaf segments during greening |
 | |
| ±: Standard error, Level of significance: p<0.05a, p<0.01b, p<0.001c | |
| Table 4: | Effect of precursors of chlorophyll on ALA formation in light using Se treated and untreated leaf segments |
 | |
| ±: Standard error, Level of significance: p<0.05a, p<0.01b, p<0.001c | |
| Table 5: | Effect of precursors of chlorophyll on ALA formation in dark using Se treated and untreated leaf segments |
 | |
| ±: Standard error, Level of significance: p<0.01b, p<0.001c | |
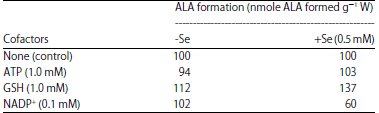
Addition of 1 mM ATP along with levulinic acid in the incubation mixture slightly reduced the ALA content in dark in -Se treated leaf segments, however, 1 mM GSH increased it slightly (Table 7). With the supply of 0.1 mM NADP+, ALA content remained same in-Se treated leaf segments. Glutathione, 1 mM and NADP+, 0.1 mM, increased and decreased the ALA content respectively in +Se treated leaf segments in dark, however, with 1 mM ATP, ALA content remained same (Table 7).
| Table 6: | Effect of supply of cofactors on ALA formation in light |
 | |
| Table 7: | Effect of supply of cofactors on ALA formation in dark |
 | |
DISCUSSION
The results of present investigation demonstrate a decline in ALA content with 0.1 and 0.5 mM Na2SeO3 supplementation; marginal increase was observed with 0.05 mM sodium selenite (Table 1). Dose dependent effect of selenate on physiological and biochemical responses of rice seedlings, with low concentration of selenate inducing a stimulatory effect on growth and metabolism against toxic effect of high concentration has been shown by Das et al.25. Further, decrease in chlorophyll content is more substantial than decrease in ALA and carotenoid contents (Table 1). This implies that apart from ALA synthesizing capacity, other steps of chlorophyll biosynthetic pathway are also being affected by Se. Inhibition of δ-Aminolevulinic acid dehydratase (δ-ALAD) activity by selenium has been reported by Jain and Gadre26 in maize leaves. Similarly As and Hg have been shown to inhibit ALAD activity in maize leaf segments27,28. Decrease in the level of carotenoids by Se may be due to effect of the metalloid on photosynthetic apparatus. Selenate toxicity at a concentration of >30 μM has been shown to decrease chlorophylls and carotenoids29.
A concentration dependent decrease in ALA content with the supply of 0.1 and 0.5 mM selenite was observed in both light and dark conditions, where the effect is prominent at higher concentration (Table 2). Possibly, selenium is promoting the catabolism of accumulated ALA in these organelles or decreasing its synthesis, resulting in lower values. Metabolism of ALA into amino and dicarboxylic acid has been reported in blue green algae30. The authors have also shown the catabolism of ALA to CO2 by etiolated barley leaves. Further, there is a growing body of evidence which indicates that ALA may be synthesized via nonporphyrin pathway in a number of organisms31-33. Aminolevulinic acid accumulation upon treatment with levulinic acid in light reflects mainly chloroplastic synthesis and in dark mainly mitochondrial synthesis. The higher value of L-D ALA content at lowest concentration of sodium selenite may be due to ineffectiveness towards this concentration of Se (Table 2).
The experiments suggest a protective role of 2-Oxoglutarate, glutamate, glutamine and Na succinate against decrease in the ALA formation by Se as these compounds increase the ALA content in presence of sodium selenite and thus, reduce the percentage decrease by Se (Table 3). Glutamate and 2-oxoglutarate being precursors of ALA might be involved in ALA synthesis. The 5-carbon skeleton of ALA has been demonstrated to be derived from 5-carbon compounds like, glutamate or α-ketoglutarate in greening leaves34. Succinate being an intermediate of TCA cycle may be involved in supplying glutamate for ALA formation. Further, magnesium seems to promote the inhibitory effect of selenium for ALA synthesis, may be through formation of Mg-Se complex. In the presence of light, percentage decrease by Se remains unaffected by the supply of 2-Oxoglutarate and glutamine, slightly decreased by Na succinate and MgCl2, however, greatly reduced by glycine. Glutamate supply increased the percentage decrease by selenite (Table 3). Glycine is suppressing the promoting effect of Se for catabolism of ALA in light; however, decrease in ALA content by glycine in absence of Se is not clear. Succinate also seems to protect the catabolism of ALA in presence of Se. The precursors of chlorophyll seem to be ineffective in causing ALA accumulation in the absence of selenite in dark. However, in the presence of Na2SeO3, all of them increase the ALA content, the effect being more pronounced with glutamate (Table 5). Hence, it is possible that the precursors of chlorophyll are acting as protectants against selenium in mitochondria for the synthesis of ALA.
Studies with cofactors indicate the enhancing role of GSH in causing accumulation of ALA in the absence of Na2SeO3 in light; however, ALA content is decreased in the presence of selenite (Table 6). Thus, GSH seems to act synergistically with levulinic acid in the absence of selenium than in its presence. Haem has been found to inhibit the formation of δ-ALA, which is enhanced by GSH in cell-free extracts of Chlorella35. ALA content remained almost same with the supply of ATP and NADP+ in the absence of selenite; however, they slightly decrease and increase the ALA content respectively in the presence of Se (Table 6). Glutathione accelerates the ALA formation in both the absence as well as presence of Se in dark, although effect is more prominent in the presence of metalloid than in its absence. Thus, mitochondrial ALA synthesis seems to be promoted by GSH. The effect of ATP and NADP+ is same in dark in the absence of Se as in light; however, in presence of Se, NADP+ substantially reduced the ALA content (Table 7).
CONCLUSION AND FUTURE RECOMMENDATIONS
This is the first report showing the effect of selenium, a metalloid on the key regulatory step of chlorophyll biosynthesis in etiolated maize leaf segments during greening. The results of the study demonstrate more prominent decrease in total chlorophylls than ALA content/formation with the supply of Se. This suggests the possibility of involvement of other steps of chlorophyll biosynthesis in addition to ALA synthesis, being affected by Se. Further, substantial decrease in L-D value at higher concentrations of Se suggests that chloroplastic ALA synthesizing activity is more sensitive towards Se. Thus, this study discovers the toxicity of selenium on chlorophyll biosynthesis in maize leaf segments through the inhibition of 1st regulatory step, i.e., formation of ALA. It will help the researchers to understand the mechanistic details on vital physiological process of chlorophyll biosynthesis.
SIGNIFICANCE STATEMENTS
This study discovers the possible site of toxicity of selenium on chlorophyll biosynthesis in maize leaf segments through the inhibition of 1st regulatory step, i.e., formation of ALA, that can be beneficial in understanding the mechanistic details on vital physiological process of chlorophyll biosynthesis. This is the maiden report showing effect of Se on very 1st step of chlorophyll biosynthesis, otherwise so far, only protective/antagonistic effects of Se against abiotic stresses and its use for bio fortification has been reported. This study will help the researchers to uncover the critical areas of regulation of chlorophyll metabolism that many researchers were not able to explore. Thus, a novel role of Selenium on inhibition of chlorophyll biosynthesis process was identified.
ACKNOWLEDGMENT
Research grant (ID MRP-MAJOR-BIOC-2013-6699) to Meeta Jain Jrom University Grants Commission (UGC), New Delhi, India as major research project is gratefully acknowledged.
REFERENCES
- Beale, S.I., 1978. δ-Aminolevulinic acid in plants: Its biosynthesis, regulation and role in plastid development. Annu. Rev. Plant Physiol., 29: 95-120.
CrossRefDirect Link - May, B.K., C.R. Bhasker, M.J. Bawden and T.C. Cox, 1990. Molecular regulation of 5-Aminolevulinate synthase. Diseases related to heme biosynthesis. Mol. Biol. Med., 7: 405-421.
PubMedDirect Link - Dalal, V.K. and B.C. Tripathy, 2012. Modulation of chlorophyll biosynthesis by water stress in rice seedlings during chloroplast biogenesis. Plant Cell Environ., 35: 1685-1703.
CrossRefDirect Link - Turan, S. and B.C. Tripathy, 2015. Salt-stress induced modulation of chlorophyll biosynthesis during de-etiolation of rice seedlings. Physiol. Plant., 153: 477-491.
CrossRefDirect Link - Balestrasse, K.B., M.L. Tomaro, A. Batlle and G.O. Noriega, 2010. The role of 5-Aminolevulinic acid in the response to cold stress in soybean plants. Phytochemistry, 71: 2038-2045.
CrossRefDirect Link - Korkmaz, A., Y. Korkmaz and A.R. Demirkıran, 2010. Enhancing chilling stress tolerance of pepper seedlings by exogenous application of 5-Aminolevulinic acid. Environ. Exp. Bot., 67: 495-501.
CrossRefDirect Link - Sun, Y.P., Z.P. Zhang and L.J. Wang, 2009. Promotion of 5-Aminolevulinic acid treatment on leaf photosynthesis is related with increase of antioxidant enzyme activity in watermelon seedlings grown under shade condition. Photosynthetica, 47: 347-354.
CrossRefDirect Link - Naeem, M.S., Z.L. Jin, G.L. Wan, D. Liu, H.B. Liu, K. Yoneyama and W.J. Zhou, 2010. 5-Aminolevulinic acid improves photosynthetic gas exchange capacity and ion uptake under salinity stress in oilseed rape (Brassica napus L.). Plant Soil, 332: 405-415.
CrossRefDirect Link - Ishizuka, M., F. Abe, Y. Sano, K. Takahashi and K. Inoue et al., 2011. Novel development of 5-Aminolevurinic acid (ALA) in cancer diagnoses and therapy. Int. Immunopharmacol., 11: 358-365.
CrossRefDirect Link - Sasaki, K., M. Watanabe, T. Tanaka and T. Tanaka, 2002. Biosynthesis, biotechnological production and applications of 5-Aminolevulinic acid. Applied Microbiol. Biotechnol., 58: 23-29.
CrossRefDirect Link - Lobanov, A.V., D.L. Hatfield and V.N. Gladyshev, 2008. Reduced reliance on the trace element selenium during evolution of mammals. Genome Biol., Vol. 9.
CrossRefDirect Link - Djanaguiraman, M., D.D. Devi, A.K. Shanker, A., Sheeba and U. Bangarusamy, 2005. Selenium-an antioxidative protectant in soybean during senescence. Plant Soil, 272: 77-86.
CrossRefDirect Link - Hawrylak-Nowak, B., R. Matraszek and M. Szymanska, 2010. Selenium modifies the effect of short-term chilling stress on cucumber plants. Biol. Trace Elem. Res., 138: 307-315.
CrossRefDirect Link - Valadabadi, S.A., A.H. Shiranirad and H.A. Farahani, 2010. Ecophysiological influences of zeolite and selenium on water deficit stress tolerance in different rapeseed cultivars. J. Ecol. Nat. Environ., 2: 154-159.
Direct Link - Cartes, P., A.A. Jara, L. Pinilla, A. Rosas and M.L. Mora, 2010. Selenium improves the antioxidant ability against aluminium-induced oxidative stress in ryegrass roots. Ann. Applied Biol., 156: 297-307.
CrossRefDirect Link - Malik, J.A., S. Goel, N. Kaur, S. Sharma, I. Singh and H. Nayyar, 2012. Selenium antagonises the toxic effects of arsenic on mungbean (Phaseolus aureus Roxb.) plants by restricting its uptake and enhancing the antioxidative and detoxification mechanisms. Environ. Exp. Bot., 77: 242-248.
CrossRefDirect Link - Hawrylak-Nowak, B., 2009. Beneficial effects of exogenous selenium in cucumber seedlings subjected to salt stress. Biol. Trace Elem. Res., 32: 259-269.
CrossRefDirect Link - Hu, Y., G.J. Norton, G. Duan, Y. Huang and Y. Liu, 2014. Effect of selenium fertilization on the accumulation of cadmium and lead in rice plants. Plant Soil, 384: 131-140.
CrossRefDirect Link - Hartikainen, H., T. Xue and V. Piironen, 2000. Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil, 225: 193-200.
CrossRefDirect Link - White, P.J. and M.R. Broadley, 2009. Biofortification of crops with seven mineral elements often lacking in human diets-iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol., 182: 49-84.
CrossRefDirect Link - Jain, M., T. Menka, P. Prachi, M. Seema, V. Jayesh and G. Rekha, 2015. Effect of arsenic on δ-Aminolevulinic acid formation in greening maize leaf segments. Indian J. Plant Physiol., 20: 191-196.
CrossRefDirect Link - Tewari, A.K. and B.C. Tripathy, 1998. Temperature-stress-induced impairment of chlorophyll biosynthetic reactions in cucumber and wheat. Plant Physiol., 117: 851-858.
CrossRefDirect Link - Jain, M. and R. Gadre, 1998. Inhibition of chlorophyll synthesis and enzymes of nitrogen assimilation by selenite in excised maize leaf segments during greening. Water Air Soil Pollut., 104: 161-166.
CrossRefDirect Link - Jain, M. and R.P. Gadre, 2004. Inhibition of 5-Aminolevulinic acid dehydratase activity by arsenic in excised etiolated maize leaf segments during greening. J. Plant Physiol., 161: 251-255.
CrossRefDirect Link - Gupta, P., S. Juliana, J. Meeta and G. Rekha, 2013. Inhibition of 5-Aminolevulinic acid dehydratase by mercury in excised greening maize leaf segments. Plant Physiol. Biochem., 62: 63-69.
CrossRefDirect Link - Hawrylak-Nowak, B., 2013. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul., 70: 149-157.
CrossRefDirect Link - Duggan, J.X., E. Meller and M.L. Gassman, 1982. Catabolism of 5-Aminolevulinic acid to CO2 by etiolated barley leaves. Plant Physiol., 69: 19-22.
CrossRefDirect Link - Troxler, R.F. and A.S. Brown, 1975. Metabolism of δ-Aminolevulinic acid in red and blue-green algae. Plant Physiol., 55: 463-467.
CrossRefDirect Link - Gassman, M.L., J.X. Duggan, L.C. Stillman, L.M. Vlcek, P.A. Castelfranco and B. Wezelman, 1978. Oxidation of Chlorophyll Precursors and its Relation to the Control of Greening. In: Chloroplast Development, Akoyunoglou, G. and J.H. Argyroudi-Akoyunoglou (Eds.). Elsevier/North Holland, Amsterdam, pp: 167-181.
- Owens, T.G., D.M. Riper and P.G. Falkowski, 1978. Studies of δ-Aminolevulinic acid dehydrase from Skeletonema costatum, a marine plankton Diatom. Plant Physiol., 62: 516-521.
CrossRefDirect Link - Beale, S.I., S.P. Gough and S. Granick, 1975. Biosynthesis of δ-Aminolevulinic acid from the intact carbon skeleton of glutamic acid in greening barley. Proc. Natl. Acad. Sci., 72: 2719-2723.
Direct Link - Weinstein, J.D., R.W. Howell, R.D. Leverette, S.Y. Grooms, P.S. Brignola, S.M. Mayer and S.I. Beale, 1993. Heme inhibition of δ-Aminolevulinic acid synthesis is enhanced by glutathione in cell-free extracts of Chlorella. Plant Physiol., 101: 657-665.
CrossRefDirect Link