
Y. Ogawa
Faculty of Pharmaceutical Sciences, Doshisha Women`s College of Liberal Arts, Kyoto, Japan
A. Minamizawa
Faculty of Pharmaceutical Sciences, Doshisha Women`s College of Liberal Arts, Kyoto, Japan
S. Tada
Faculty of Pharmaceutical Sciences, Doshisha Women`s College of Liberal Arts, Kyoto, Japan
T. Konishi
Faculty of Pharmaceutical Sciences, Doshisha Women`s College of Liberal Arts, Kyoto, Japan
Research Journal of Phytochemistry
Year: 2013 | Volume: 7 | Issue: 1 | Page No.: 10-17







ABSTRACT
Oxypinnatanine, a unique derivative of glutamic acid or glutamine with a furfuryl group increased the total time of NREM sleep by an oral administration. However the mechanism by which oxypinnatanine promotes sleep remains to be clarified. In this study, the effect of oxypinnatanine on heat release of freely moving mice from peripheral blood vessel was demonstrated by using thermography. Oxypinnatanine given at 30 mg kg-1 indicated the transient skin temperature elevation from 2 min after its administration but the skin temperature was decreased from 32 min after. Therefore, it is possible that the thermal release associated with the dilation of peripheral vessels is one of the mechanisms of oxypinnatanine about its promoting sleep. In addition, the effects of amino acid amides (30 mg kg-1) which isolated from Hemerocallis fulva var. sempervirens was also examined. As a result, pinnatanine and kwansonine A decreased the temperature, while longitubanine A, longitubanine B and kwansonine B did not. As changes in body temperature are generally associated with concomitant changes in sleep propensity and the effects of sleep promoting substances are related to changes in thermoregulation. Therefore, it is possible that the monitoring on heart release from peripheral blood vessel with the use of thermography might enable the search for substances which induce sleep.
PDF Abstract XML References Citation
Received: February 21, 2013;
Accepted: April 08, 2013;
Published: June 29, 2013
How to cite this article
Y. Ogawa, A. Minamizawa, S. Tada and T. Konishi, 2013. Variation of Body Temperature after Administration of Amino Acid Amides. Research Journal of Phytochemistry, 7: 10-17.
URL: https://scialert.net/abstract/?doi=rjphyto.2013.10.17
URL: https://scialert.net/abstract/?doi=rjphyto.2013.10.17
INTRODUCTION
The importance of sleep has recently been emphasized throughout the world as there are many people who have sleep problems in the modern world. For example, a meta-analysis has revealed that approximately 30% of the general population present with insomnia symptoms (Ohayon, 2002). Therefore the amelioration of insomnia is very important for society. There are many causes of insomnia such as poor physical condition, changing environment, mental stress and alcohol or medicine side effects, to name a few. Though there are several possible treatments according to the cause, one of the treatments, controlling body temperature, has been attracting attention recently. For example, many people who have poor blood circulation may have a sleeping problem or those who have menopausal troubles may not be able to sleep because of feeling very flushed. Sleep and body temperature is significantly correlated. The body temperature includes core body temperature and skin temperature and changes in core body temperature are generally interrelated with sleep (Zulley et al., 1984). The core body temperature drops with the onset of sleep, continues to decrease during sleep and gradually rises with the awakening from sleep (Czeisler et al., 1980; Zulley et al.,1984; Barrett et al., 1999; Foret et al., 1993). Therefore, changes of core body temperature mediate the changes in sleepiness.
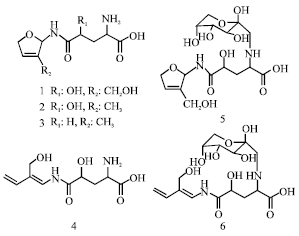 | |
| Fig. 1: | Structures of isolated compounds from Hemerocallis fulva var. sempervirens |
The core body temperature decreases because of the release of heat from a peripheral site and it develops the following process. (i) The peripheral blood vessel dilates and the heat transfer occurs as a result of increased blood flow (ii) the skin temperature rises with the transfer of heat (iii) the core body temperature and skin temperature decrease in conjunction with the increase of the amount of thermal release. Then, it might be possible to monitor this thermal release in sleep promoting stage by measuring of the skin temperature using thermography. Moreover, the monitoring of it might be useful when searching for substances that can induce sleep. Here, the thermal release in an initial stage of the sleep reaction was investigated by using a thermography.
In the continuing study for sleep promoting substances from natural resources, it was found that the flowers and leaves of Hemerocallis fulva L. var. sempervirens (Araki) M. Hotta (Liliaceae) are known to be taken as an aid for sleeping in Okinawa, Japan (Uezu, 1998). Previously, four known amino acid amides, oxypinnatanine (1), longitubanine A (2) and longitubanine B (3), pinnatanine (4) and two novel amino acid amides, kwansonine A (5) and kwansonine B (6), were isolated from the leaves of this plant (Ogawa and Konishi, 2009). These structures are shown in Fig. 1. Moreover, it was demonstrated by electroencephalographic analyses that an oral administration of oxypinnatanine (1) to mice increased non-rapid eye movement (non-REM, NREM) sleep in a dose-dependent manner (Ogawa et al., 2013). However, the mechanism by which oxypinnatanine promotes sleep remains to be clarified.
In the present study, the mechanism of oxypinnatanine (1)-induced sleep was investigated, using the monitoring of thermal release with dilatation of peripheral vessels. Furthermore, the effects of amino acid amides, which were isolated from H. fulva var. sempervirens were also investigated.
MATERIALS AND METHODS
Materials: Diazepam (cercine) (Qiu et al., 2009) and ramelteon (rozerem) (Miyamoto, 2006) were from Takeda Pharmaceutical Co., Ltd. Zolpidem (myslee) (Vinkers et al., 2010) was from Astellas Pharma Inc. Diphenhydramine (restamin kowa tablet) (Matsuoka and Kubota, 1897) was from Kowa Co., Ltd. These agents were dissolved in water. Glycine (Bannai and Kawai, 2012) was purchased from Nacalai Tesque; capsaicin (Kawabata et al., 2009) was from Wako Co., Ltd. Amino acid amides (compounds 1-6) were extracted and purified from H. fulva var. sempervirens. Separation of these compounds was carried out according to the method described previously (Ogawa and Konishi, 2009).
Animals: Male ddY mice, 18-28 g (4-6 weeks old), were used in this study. They were housed at an ambient temperature of 24±2°C on an automatically controlled 12: 12 h light-dark cycle (light on at 08:00, illumination intensity of approximately 100 lux). The experimental protocols were approved by the Animal Care Committee of Doshisha Women’s College of Liberal Arts.
Skin temperature measurement: Temperature were continuously monitored at 24±2°C. Each mouse was placed in an individualized cage for 90 min prior to the experiment and throughout the measurements. The skin temperature of mice was monitored with a calibrated IR camera (FLIR E60 Compact Thermal Imaging, 320x240 pixels IR Resolution). During IR recordings, both the mice and the camera were kept away from external IR sources and the distance between mice and the camera was 80 cm. The skin temperature of unanesthetized mice were tracked (n = 5). The normal skin temperature was measured for 30 min before the experiment. The temperature was measured for 80 min from administration of water or test substances. Average temperature on the back of mice was subsequently computed for every IR image using FLIR R and D software from FLIR Systems. All results were expressed as mean±SE of the difference of the normal skin temperature of each mouse due to large individual differences in skin temperatures.
Statistical analysis: Two-way analysis of variance (ANOVA) was used to test for statistical differences. When significant differences (p<0.05) were identified, the data were further analyzed by Student’s t-test.
RESULTS AND DISCUSSION
The sleeping drugs: Diazepam (benzodiazepines group), zolpidem (not benzodiazepines group) and ramelteon (agonist of melatonin receptors) were used with the monitoring of the change of the skin temperature.
Diazepam is one of the most famous sleeping drugs and it has been the mainstay of therapy for insomnia and anxiety. Since diazepam was discovered in the 1950s, a large number of benzodiazepines analogs have been synthesized to obtain a superior risk/benefit ratio. Compared with the water group, diazepam at 12 mg kg-1 immediately and significantly decreased the skin temperature 2 min after administration by 0.7°C and 30 min after by 1.7°C (Fig. 2a). This result was consistent with the time when the mice were promoted into sleep.
Zolpidem is a widely used hypnotic drug which potentiates GABAergic neurotransmission by acting at the benzodiazepine site of GABAA receptors (Langer et al., 1992). Zolpidem at 10 mg kg-1 decreased the skin temperature 0.5°C to 8 min and 1.3°C to 22 min after its administration, as compared with the water administrated group (Fig. 2b).
Ramelteon is a selective agonist to melatonin MT1/MT2 receptors in the suprachiasmatic nucleus; SCN (Kato et al., 2005) and has circadian (Richardson et al., 2008) and sedative effects (Roth et al., 2005). When ramelteon at 10 mg kg-1 was administrated, the skin temperature was decreased 8 min after its administration statistically significantly and -1.0°C was indicated at 22 min as compared with the water group (Fig. 2c). These suggested that the skin temperature is markedly decreased by administration of sleeping drugs.
Diphenhydramine is an antagonist to histamine H1 receptor and clinical use of H1 receptor promotes sleepiness as a side effect (Nicholson et al., 1985). Diphenhydramine given at 50 mg kg-1 significantly decreased the skin temperature by 1.5°C like sleeping drugs from 20 min (Fig. 2d).
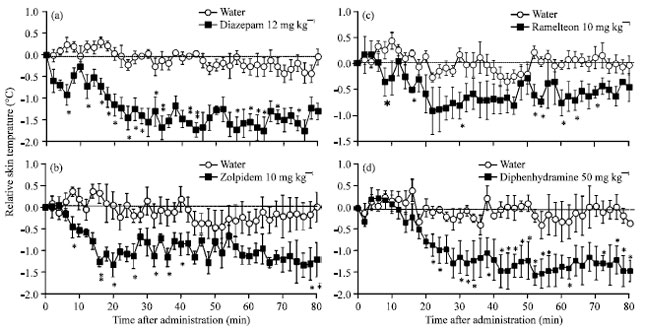 | |
| Fig. 2(a-d): | The effect of sleeping drugs on the skin temperature of mice, administration group (n = 5). Each circle represents the mean±SE of the difference of the normal skin temperature of each mouse. *p<0.05, **p<0.01 versus water group by student’s t-test after two-way analysis of variance was used to test for statistical differences (p<0.05) |
Therefore, it is possible to monitor thermal releases in sleep promoting stage by measurement of the skin temperature using thermography. There have been many reports on the relationship of core body temperature or skin temperature and sleep but this is the first report using thermography. Then, this assay method was used about a sleep promoting substances.
Several natural compounds have been shown to improve insomnia in mice; for example glycine (Yamadera et al., 2007), verbenalin (Makino et al., 2009), crocin (Masaki et al., 2012) and ornithine (Omori et al., 2012) were reported to be effective in promoting NREM sleep. Especially, glycine (2 g kg-1, p.o.) improve sleep by decreasing core body temperature with dilatation of peripheral vessels (Bannai and Kawai, 2012). Then the effect of glycine on the skin temperature was demonstrated. As a result, Glycine at a dose of 2 g kg-1 was risen the transient skin temperature elevation (+0.5°C) after its administration, because of dilatation of peripheral vessels and it was decreased (-1.2°C) from 30 min after its administration (Fig. 3a). This result was consistent with the literature data or the sleep-onset time (Brooks and Peever, 2011). Therefore it is possible to make a measurement of the transient temperature elevation with dilatation of peripheral vessels by using thermography measurement. Next, the effect of a substance which has a vasodilator action was demonstrated.
Capsaicin is the major ingredient in hot peppers of the plant genus Capsicum. It is used extensively in foods and is also utilized as a traditional medicine worldwide for the treatment of various disorders. Capsaicin (10 mg kg-1, p.o.), which is known as a vasodilating agent (Salem and Dunbar, 2002) significantly elevated the skin temperature immediately after its administration; and the effect was statistically significant from 14 min after the administration (Fig. 3b). However it did not decrease the body temperature compared with the water administration and it did not change the amount of sleep time (Surh et al., 1995). Thus, only vasodilating does not seem to induce sleep but it seems to be more important for the introduction of sleep through its decrease of body temperature with thermal release.
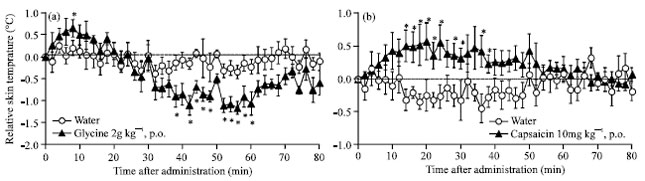 | |
| Fig. 3(a-b): | The effect of glycine or capsaicin on the skin temperature of mice, administration group (n = 5). Each circle represents the mean±SE of the difference of the normal skin temperature of each mouse. *p<0.05 versus water group by student’s t-test after two-way analysis of variance was used to test for statistical differences (p<0.05) |
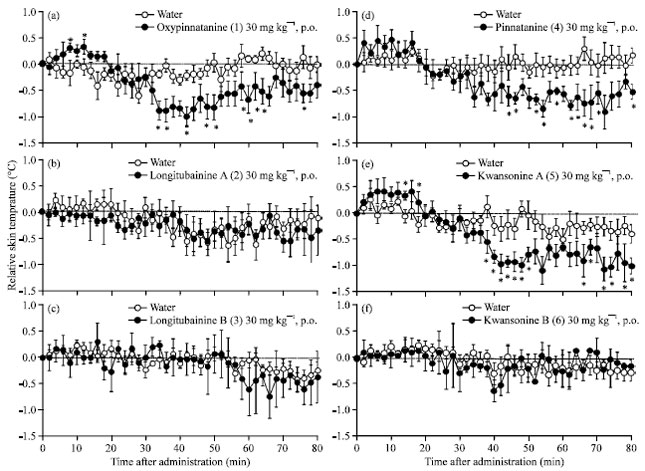 | |
| Fig. 4(a-f): | The effect of amino acid amides (compounds 1-6) isolated from H. fulva var. sempervirens, administration groups (n = 5). Each circle represents the mean±SE of the difference of the normal skin temperature of each mouse (n = 5). *p<0.05 versus water group by student’s t-test after two-way analysis of variance was used to test for statistical differences (p<0.05) |
There the effect of oxypinnatanine (1) which was isolated from H. fulva var. sempervirens was examined. As a result, 1 given at 30 mg kg-1 indicated the transient skin temperature elevation (+0.3°C) after its administration but it was decreased (-1.0°C) from 32 min after its administration (Fig. 4a). Because this result showed the same tendency with glycine, it is suggested that the thermal release associated with the dilation of peripheral vessels is enumerated in one of the mechanisms of 1 on promoting sleep. These results indicate that the monitoring system of skin temperature could be utilized as one of the assay methods to search for substances that promote the sleep.
The effects of compounds (2)-(6) which were isolated from H. fulva var. sempervirens were also examined. Compounds (2)-(4) have been isolated from only a few plants, such as Euscaphis japonica (Grove et al., 1973) and Honkenya peploides (Cerantola et al., 2005), besides the Hemerocallis species (Inoue et al., 1990). Moreover, there has been no study about the bioactivity of these compounds. In this study, 4 and 5 at a dose of 30 mg kg-1 showed the same tendency of glycine or 1. In contrast, compounds 2, 3 and 6 showed no effect on skin temperature of the mice (Fig. 4b-f). Therefore, it was suggested that compounds 4 or 5 might have a sleep promoting effect and, 2, 3 or 6 have no effect. Further study is being done on the sleep promoting activity of compounds (2)-(6).
CONCLUSION
In this study, it is indicated that the thermal release in sleep promoting stage can be monitored by using a thermography. Moreover, oxypinnatanine (1) indicated the transient skin temperature elevation after it’s administration and that temperature was decreased after the transient elevation. Thus, the thermal release associated with the dilation of peripheral vessels is one of the mechanisms of 1 about its promoting sleep.
ACKNOWLEDGMENTS
This research was financially supported by a Grant-in-Aid for Scientific Research (C) (No. 22590025) from the Japan Society for the Promotion of Science.
REFERENCES
- Bannai, M. and N. Kawai, 2012. New therapeutic strategy for amino acid medicine: Glycine improves the quality of sleep. J. Pharmacol. Sci., 118: 145-148.
CrossRef - Barrett, J., L. Lack and M. Morris, 1999. The sleep-evoked decrease of body temperature. Sleep, 16: 93-99.
PubMedDirect Link - Brooks, P.L. and J.H. Peever, 2011. Impaired GABA and glycine transmission triggers cardinal features of rapid eye movement sleep behavior disorder in mice. J. Neurosci., 31: 7111-7121.
CrossRefDirect Link - Cerantola, S., M.A. Bessieres, C. Magne and E. Deslandes, 2005. Occurrence of the unusual amino acid N5-(hydroxymethyl-2,5-dihydro-2-furyl)-L-allo-γ-hydroxyglutamine in Honkenya peploides (L.) Ehrh. Biochem. Syst. Ecol., 33: 1187-1189.
Direct Link - Czeisler, C.A., E. Weitzman, M.C. Moore-Ede, J.C. Zimmerman, R.S. Knauer, 1980. Human sleep: Its duration and organization depend on its circadian phase. Science, 210: 1264-1267.
PubMedDirect Link - Foret, J., A. Aguirre, Y. Touitou, M. Clodore and O. Benoit, 1993. Effect of morning bright light on body temperature, plasma cortisol and wrist motility measured during 24 hour of constant conditions. Neurosci. Lett., 155: 155-158.
CrossRef - Grove, M.D., D. Weisleder and M.E. Daxenbichler, 1973. Pinnatanine and oxypinnatanine, novel amino acid amides from Staphylea pinnata L. Tetrahedron, 29: 2715-2719.
CrossRef - Inoue, T., K. Iwagoe, T. Konishi, S. Kiyosawa and Y. Fujiwara, 1990. Novel 2,5-dihydrofuryl-γ-lactam derivatives from Hemerocallis fulva L. var. Kwanzo Regel. Chem. Pharm. Bull., 38: 3187-3189.
Direct Link - Kato, K., K. Hirai, K. Nishiyama, O. Uchikawa and K. Fukatsu et al., 2005. Neurochemical properties of ramelteon (TAK-375), a selective MT1/MT2 receptor agonist. Neuropharmacolgy, 48: 301-310.
Direct Link - Kawabata, F., N. Inoue, Y. Masamoto, S. Matsumura and W. Kimura et al., 2009. Non-pungent capsaicin analogs (capsinoids) increase metabolic rate and enhance thermogenesis via gastrointestinal TRPV1 in mice. Biosci. Biotechnol. Biochem., 73: 2690-2697.
CrossRefDirect Link - Makino, Y., S. Kondo, Y. Nishimura, Y. Tsukamoto, Z.L. Huang and Y. Urade, 2009. Hastatoside and verbenalin are sleep-promoting components in Verbena officinalis. Sleep Biol. Rhythms, 7: 211-217.
CrossRef - Masaki, M., K. Aritake, H. Tanaka, Y. Shoyama, Z.L. Huang and Y. Urade, 2012. Crocin promotes non-rapid eye movement sleep in mice. Mol. Nutr. Food Res., 56: 304-308.
CrossRef - Matsuoka, Y. and K. Kubota, 1897. Characteristics of inflammation induced by fusarenon-X, a trichothecene mycotoxin from Fusarium species. Toxicol. Applied Pharmacol., 91: 333-340.
CrossRef - Nicholson, A.N., P.A. Pascoe and B.M. Stone, 1985. Histaminergic systems and sleep studies in man with H1 and H2 antagonists. Neuropharmacology, 24: 245-250.
PubMedDirect Link - Ogawa, Y. and T. Konishi, 2009. N-Glycosides of amino acid amides from Hemerocallis fulva var. sempervirens. Chem. Pharm. Bull. (Tokyo), 57: 1110-1112.
CrossRefPubMedDirect Link - Ogawa, Y., N. Uchiyama, T. Konishi and Y. Urade, 2013. Oxypinnatanine promotes non-rapid eye movement sleep in mice. Sleep Biol. Rhythms, 11: 45-50.
CrossRefDirect Link - Ohayon, M.M., 2002. Epidemiology of insomnia: What we know and what we still need to learn. Sleep Med. Rev., 6: 97-111.
CrossRefPubMedDirect Link - Omori, K., Y. Kagami, C. Yokoyama, T. Moriyama and N. Matsumoto et al., 2012. Promotion of non-rapid eye movement sleep in mice after oral administration of ornithine. Sleep Biol. Rhythms, 10: 38-45.
CrossRefDirect Link - Qiu, M.H., W.M. Qu, X.H. Xu, M.M. Yan, Y. Urade and Z.L. Huang, 2009. D1/D2 receptor-targeting L-stepholidine an active ingredient of the Chinese herb Stephonia induces non-rapid eye movement sleep in mice. Pharmacol. Biochem. Behav, 94: 16-23.
CrossRefPubMedDirect Link - Richardson, G.S., P.C. Zee, S. Wang-Weigand, L. Rodriguez and X. Peng, 2008. Circadian phase-shifting effects of repeated ramelteon administration in healthy adults. J. Clin. Sleep Med., 4: 456-461.
Direct Link - Roth, T., C. Stubbs and J.K. Walsh, 2005. Ramelteon (TAK-375) a selective MT1/MT2-receptoroagonist reduces latency to persistent sleep in a model of transient insomnia related to a novel sleep environment. Sleep, 28: 303-307.
Direct Link - Salem, N. and J.C. Dunbar, 2002. The insulin-mediated vascular and blood pressure responses are suppressed in CGRP-deficient normal and diabetic rats. Diabetes Metab. Res. Rev., 18: 238-244.
CrossRefPubMedDirect Link - Surh, Y.J., S.H. Ahn, K.C. Kim, J.B. Park, Y.W. Sigb and S.S. Lee, 1995. Metabolism of capsaicinoids: Evidence for aliphatic hydroxylation and its pharmacological implications. Life Sci., 56: PL305-PL311.
CrossRefDirect Link - Uezu, E., 1998. Effects of Hemerocallis on sleep in mice. Psychiatry Clin. Neurosci., 52: 136-137.
Direct Link - Vinkers, C.H., J.F. Cryan, B. Olivier and L. Groenink, 2010. Elucidating GABAA and GABAB receptor functions in anxiety using the stress-induced hyperthermia paradigm: A review. Open Pharm. J., 4: 1-14.
Direct Link - Yamadera, W., K. Inagawa, S. Chiba, M. Bannai, M. Takahashi and K. Nakayama, 2007. Glycine ingestion improves subjective sleep quality in human volunteers correlating with polysomnographic changes. Sleep Biol. Rhythms, 5: 126-131.
CrossRefDirect Link