
E.A. Adebayo
Department of Pure and Applied Biology, Ladoke Akintola University of Technology, P.M.B. 4000, Ogbomoso, Nigeria
J.K. Oloke
Department of Pure and Applied Biology, Ladoke Akintola University of Technology, P.M.B. 4000, Ogbomoso, Nigeria
A.K. Bordoloi
North East Institute of Science and Technology, CSIR, Jorhat-785006, Assam, India
M. Barooah
Assam Agricultural University Jorhat-13, Assam, India
T.C. Bora
North East Institute of Science and Technology, CSIR, Jorhat-785006, Assam, India
Research Journal of Mutagenesis
Year: 2014 | Volume: 4 | Issue: 1 | Page No.: 1-13







ABSTRACT
High production and good quality are always the principal goals for agriculturally important crops, without the exception of mushrooms. P. pulmonarius is one of the commercially important edible mushrooms throughout the world. The present study was designed to improve the potential of P. pulmonarius LAU 09 for yields production. The improvement of P. pulmonarius LAU 09 was carried out by UV light exposure at 210 nm for 3 h, at 30 min interval. The highest rate of spawn ramification (0.889 cm day-1) and percentage spawn productivity (47.64%) was obtained by LAU 90. The primordia of mushroom sporophore recorded at 11th day of cultivation by LAU 60 and LAU 90 while recorded on 15th day in LAU 09. The highest biological efficiency (97.57%) and production rate (3.9%) obtained by LAU 90 while the least of 41.43 and 1.34%, respectively was obtained by LAU 09. The morphological and molecular characterization of the mutant strains established their true variation from their wild type. The wider evolutionary lineages of LAU 60 (mutant) from LAU 09 (wild) was shown, with LAU 09 located at outgroup root of phylogenetic tree and LAU 60 formed a phylogenetic cluster with another strains of P. pulmonarius (AY450349) from GenBank database, supported with 85% bootstrap value, while LAU 90 (mutant) match no strain at GenBank database collection. The result obtained shows the improved performance of the mutant strains over the wild (parent) strains, especially the mutant at 90 min exposure (LAU 90).
PDF Abstract XML References Citation
Received: June 12, 2012;
Accepted: March 09, 2013;
Published: April 15, 2014
How to cite this article
E.A. Adebayo, J.K. Oloke, A.K. Bordoloi, M. Barooah and T.C. Bora, 2014. Improvement of Pleurotus pulmonarius Lau 09 Through Mutation for
Yield Performance. Research Journal of Mutagenesis, 4: 1-13.
DOI: 10.3923/rjmutag.2014.1.13
URL: https://scialert.net/abstract/?doi=rjmutag.2014.1.13
DOI: 10.3923/rjmutag.2014.1.13
URL: https://scialert.net/abstract/?doi=rjmutag.2014.1.13
INTRODUCTION
The development and improvement of edible mushroom strains should be an active topic for continuous research within the field of mushroom cultivation owing to their enormous importance to humans, especially Pleurotus species. Pleurotus species are regarded as one of the most important commercially cultivated edible mushrooms throughout the world (Kibar and Perksen, 2008). One of the prominent species is P. pulmonarius, an edible mushroom with high protein content and having medicinal value with anti-tumor, anti-inflammatory and antimicrobial properties (Jose et al., 2002; Stajic et al., 2007). To date, strain improvement of P. pulmonarius among the Pleurotus species has been a difficult task, due to its intersterility behavior (Bao et al., 2004) and ecologolical specificity (Kuo, 2009), factors responsible for low performance of P. pulmonarius in several region, especially in cold weather condition (Kuo, 2009).
In obtaining mutants of fungi strain, the use of ultraviolet (UV) light is better than that of X-rays, because of its simplicity, inexpensive and readily available of germicidal UV lamp. The germicidal effect and the absorption maximum of nucleic acids at 260 nm is related (i.e; 1 nanometer = 10 angstroms, symbolized Å) (Royse and May, 1993), which is included in the wavelengths delivered by the germicidal lamp. Actually, most germicidal lamps deliver a broader range of wavelengths, but experimentation has demonstrated that 260 nm is the effective wavelength. This wavelength has also been shown to increase mutation frequency. The formation of covalent bonds between adjacent pyrimidine nucleotides, generally thymine but sometimes cytosine usually resulted to germicidal (lethal) effect and the mutagenic effect. These linked pyrimidine nucleotides are referred to as dimers, either thymine dimers or cytosine dimers, depending on the specific nucleotides that have bonded together (Elliott and Langton, 1981). The consequence of dimer formation is an inhibition of normal DNA synthesis. It has also been demonstrated that there is a mechanism that can repair the UV-induced damage. The mechanism requires exposure to wavelengths in the approximate range of 360 to 480 nm. Light in this range has the effect of activating an enzyme that splits the dimer, thus permitting the return to normal DNA synthesis and lack of mutagenicity. This light repair is called photoreactivation (Murakami and Takemam, 1975). In experimental work in which UV is being used to obtain mutants, it is important that cells not be exposed to the wavelengths inducing photoreactivation. This can be accomplished by illuminating the work area with yellow light, which does not transmit wavelengths causing photoreactivation (Pukkila, 1993; Royse and May, 1993). Mutations are caused by UV through dimer formation, which is followed by replication of DNA. The dimers are responsible for gaps in the strands of DNA that are synthesized. This method of mutagenesis is different from the mode of action of ionizing radiation, e.g., X-rays, which cause alteration in the bases of the DNA and breaks in the DNA strands.
Ultra violent radiation (UV light) has been reported as one of the best physical methods for strain improvement for better yield performance (Kang et al., 1999). This method has been employed in improving enzyme production in Aspergillus niger (Kang et al., 1999) and Rhizopus oryzea (Suntornsuk and Hang, 1994), as well as mycelia cell and sporophore production in P. florida and P. sajor-caju (Ravishankar et al., 2006). However, no report is available on strains improvement of P. pulmonarius using UV light radiation exposure.
The present investigation was undertaken to improve the yield production (in term of mycelia growth and sporophore) of the P. pulmonarius LAU 09 strain through exposure to UV light radiation.
MATERIALS AND METHODS
This study was carried between the spaces of fourteen Months (June, 2009 to August, 2011). The dikaryotic mycelium of P. pulmonarius LAU 09 was isolated from fruit bodies, characterized to the species level and registered at the GenBank database. The strain was maintained on Potato Dextrose Agar (PDA) slant at 4°C, until further uses.
Mutation induction: The mutants were obtained through the exposure of an actively growing culture (7 days old) to UV-light at 210 nm for 3 h, at 30 min interval. The mutants were subcultured and incubated at 25°C for 7 days with wild type as control. The diameters of mycelia growth were taken into consideration.
Temperature tolerance of the mutants: The temperature optima for mycelium linear growth were assessed on PDA with 5% of YEA over a range of 15°C -35°C at 5°C increment. Experiments were conducted on Petri dishes (Zervakis et al., 2001), incubated for 7days and linear growth rates were determined by:
Y = KrX+C
where, Y is the distance, X is time, with diameter of mycelia growth.
The optima pH for mycelia growth: The pH optima for mycelia growth were determined using Potato Dextrose Broth (PDB) with 5% yeast extract powder (YE) over a range of 4 to 9, adjusted with (0.1 M) HCl and NaOH. Experiments were carried out in 250 mL flasks containing 100 mL substrate. The flasks were inoculated with a plug (6 mm) and incubated at 25°C for 7 days at 150 rpm. The mycelia mats were harvested and quantified (Oloke et al., 2009).
Morphological examination: The actively growing cultures (5 days old) were stained with Lactophenol blue and examined under the light microscope (OLYMPUS CX41).
Spawn production: Paddy grains (100 g bottle-1) were washed four times, boiled for 45 min and dried. The dried grains were mixed with 1%w/w of Calcium carbonate (CaCO3), dispensed in bottles and sterilized at 121°C for 30 min. The sterile grains were inoculated with 6 plugs (6 mm) of actively growing culture, incubated at 23±2°C and mycelia running were recorded at three days interval. The following parameters were determined; rate of mycelia ramification, ramification days, weight of ramified mycelia and spawn productivity.
Sporophore/fruit body production (rice straw as substrate: Paddy straw was shredded and soaked for 24 h (1 straw: 3 water). The straw was boiled for 3 h (pasteurization), dried in sterile condition and packed into sterile bags (1.4 kg bag-1), inoculated with spawn (5-10%w/w) and carefully shaken down to allow even spawn distribution throughout the bag. The bags were incubated in the dark at 25±3°C, until fully colonized. The colonized bags were transferred into a mushroom house and watering was carried out periodically for sporophore production.
Amino acids Sequence and Composition analysis: The nucleotides sequences of wild and mutant strains were translated into the encoded amino acid compositions to determine the degree of variation among strains.
DNA extraction and ITS amplification: Mycelia were grown on potato dextrose agar, harvested using a scalpel and transferred into Epperdorf tubes. Small amount of autoclaved refined sand (Sigma) were added to each tube and the mycelia ground to a fine paste with a pestle-like stick (High Media), 400 μL of DNA Extraction buffer pH 8 (1M Tris-Cl pH 8.0; 1M NaCl; 200 mM EDTA pH 8.0; 10% SDS; 0.1%β-Mercaptoethanol) was added and the solution centrifuged at 4°C (12000 g) for 10 min. To the collected supernatant 300 μL Phenol and 300 μL Chloroform: Isoamylalcohol (24:1) were added and mixed gently. This was centrifuged (12000 g, 4°C for 10 min) and the aqueous phase was collected and 500 μL chilled Isopropanol was added and incubated at -20°C overnight. After the incubation, it was centrifuged (12000 g, 4°C for 10 min) and the pellet was washed with chilled 70% ethanol centrifuged for 5 min. The dried pellet was resuspended in 50 μL of Tris EDTA (10 Mm Tris and 1 mM EDTA, pH 8.0) buffer.
Amplification of the ITS region of the rRNA gene was carried out with a modified method of Gardes and Bruns (1993), using primers ITS1-F and ITS4-B (White et al., 1990). Each sample was used as template in a 25 μL PCR reaction volume consisting of; 200 μM each of dATP, dCTP, dGTP and dTTP, 2.5 mM MgCl2, 10X Taq. DNA Polymerase and 20 Pico mole of each of the two primers (Banglore Genei). The PCR profiles included an initial denaturation step of 94°C for 85 sec followed by 25 amplification cycles of denaturation, annealing and extension. The temperature and times for these steps were 95 OC for 35 sec, 55 OC and 55 sec and 72 OC for 2 min with a final incubation at 72°C for 10 min. The amplified PCR products were resolved on a 1.2% agarose gel and stained with Ethidium bromide. A 1kb ladder DNA marker (GeneRulerTM ) was used as a size standard.
Sequencing and phylogenetic analysis: The PCR products were purified using Exonuclease I and Shrimp Alkaline phosphatas in buffer (EXOSAP Kits). Both strands of the amplified region were sequenced using fluorescent dye terminator chemistry and were run on ABI 3130 (4 capillary) or 3730XI (96 capillary) Automated Sequencer (Perkin Elmer Applied Biosystems, Foster City, CA), following the manufacturer’s protocols. Sequencing primers were ITS1-F, 5.8S, 5.8SR and ITS4-B. Oligonucleotide sequences for primers 5.8S and 5.8 SR were given by Vilgalys and Hester (1990). Sequence contigs were assembled and edited using Sequencer 3.0 software (Gene codes Corporation, Ann Arbor, MI).
Phylogenetic trees were constructed by using all cloned sequences together with all nonredundant large subunit (nLSU) sequences of named Pleurotus species obtained from GenBank. The multiple alignments of all the sequences were performed using CLUSTAL W (http://www.ebi.ac.uk/Tools/msa/clustalw2/), followed by manual adjustments. The phylogenetic analyses was carried out using sequence data of ITS 5.8s and 28s ribosomal RNA gene from LAU 09 (wild) and LAU 60 (mutant) of P. pulmonarius and corresponding GenBank data of related species. The identical sequences were merged into one input sequence when running the computer programs to generate the phylogenetic trees constructed by UPGMA, Neighbor-joining (NJ) and parsimony methods. The bootstrap test for estimating the reliability of phylogenetic tree topology was performed using 100 replications by the SEQBOOT program (Felsenstein, 1989). The consensus tree was obtained by running the consense program (Felsenstein, 1989).
Statistical analysis: ANOVA for the mycelia yield production under different conditions of growth factors was performed using SPSS software (Version 16) with significantly different at 0.05 probability level according to Duncan Multiple Range Test. The data used were in triplicate. The accession means for the mycelia yield measured were to compare the performance of the strains. Accession differences were determined by Duncan Multiple Range Test (DMRT).
RESULTS
The improved mutants strain were obtained between 30 to 120 min of UV-light exposure (LAU 30, LAU 60, LAU 90 and LAU 120), with highest mycelia growth at LAU 90 and the least at LAU 120 in comparison with the parent strain (LAU 09), while LAU 150 and LAU 180 produced significantly less mycelia yields (Fig. 1).
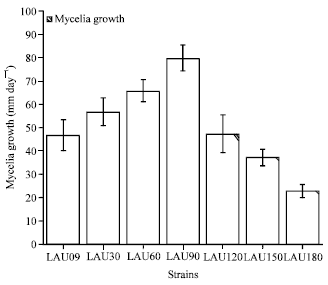 | |
| Fig. 1: | Mycelia growth (mm day-1) of wild and mutant strains of Pleurotus pulmonarius after 7 days incubation |
| Table 1: | Linear growth rates (mm day-1) of wild and mutant strains of P. pulmonarius at different temperature ranges |
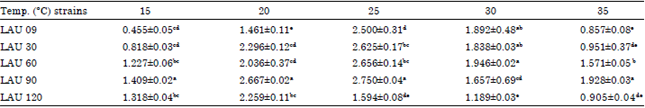 | |
| The mean values on the same column followed by different letters are significantly different at 0.05 probability level according to Duncan multiple range test | |
| Table 2: | Weight of dried mycelia yield of wild and mutant strains of P. pulmonarius at different pH values |
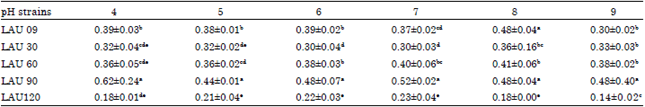 | |
| The mean values on the same column followed by different letters are significantly at 0.05 probability level according to Duncan multiple test | |
The Linear growth rates of wild and mutant strains of P. pulmonarius at different temperature ranges are shown in Table 1, with satisfactory growth obtained at 20°C and optima growth temperature at 25°C for all the strains, while LAU 90 (mutant) produced high yield at all evaluated temperature values. The means of mycelia yield obtained by LAU 90 (mutant) were significantly different (p<0.05) from all other strains in all evaluated temperature values except at 30°C. The highest mean value of linear growth rates (2.750 mm day-1) was obtained by LAU 90 at 25°C, while the lowest mean value of linear growth rates (0.455 mm day-1) was obtained by LAU 09 at 15°C. The highest means of dried mycelia weight (0.62 g) was obtained by LAU 90 (mutant) at pH 6 and lowest mycelia yield (0.14 g) was obtained at pH 9 by LAU 120 (mutant) (Table 2). The means value of mycelia yield by LAU 90 were significantly different (p<0.05) at pH 4, 7 and 9.
| Table 3: | Spawn ramification rates (cm day-1ay) of wild, mutant and hybrid strains of P. pulmonarius |
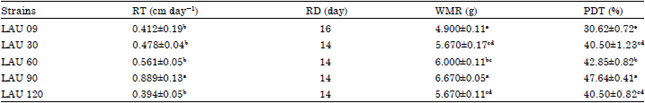 | |
| The mean values on the same column followed by different letters are significantly at 0.05 probability level according to Duncan multiple test, RT: Ramification rate, RD: Ramification days, WMR: Weight of mycelia ramification, PDT: Spawn productivity | |
| Table 4: | Distribution of the total fresh mushroom production (g) over three harvest by wild and mutants strain of P. pulmonarius on paddy straw |
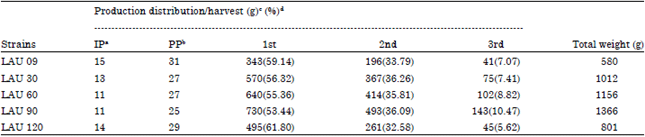 | |
| IP: Incubation period (required days for formation of primordia), PP: Production period, starting with the formation of primordia till third harvest, Weight of mushrooms obtained in 5 replicates, Percent production for the strains during each harvest | |
| Table 5: | Biological efficiency (%) and Production rate (%/d) of the wild and mutant strains of P. pulmonarius |
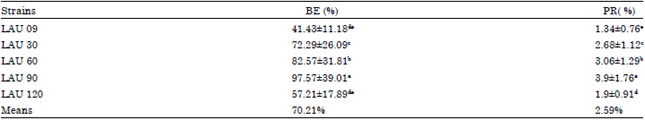 | |
The mean values on the same column followed by different letters are significantly at 0.05 probability level according to Duncan multiple test, BE: Biological efficiency (productivity) of the each strain, PR: Production rate of the each strain | |
The highest mean of spawn ramification rates (0.887 cm day-1), highest means of weight of mycelia ramification (6.670 g) and highest means of percentage spawn productivity (47.64%) with shortest days of ramification (12 days) were obtained by LAU 90 (mutant). The lowest means of spawn ramification rates (0.394 cm day-1) was obtained by LAU 120 and lowest means of percentage spawn productivity (30.62%), lowest values of weight of mycelia ramification (4.900 g) and longest days of ramification (16 days) obtained by LAU 09 (wild) (Table 3).
Table 4 and 5 showed the number of flushes, Biological Efficiency (BE) and Production Rate (PR). Strains required 11 days (LAU 60 and LAU 90) to 15 days (LAU 09) after inoculation to produce their first fruiting bodies. The time required for the culture cycles to reach three flushes was shortest for strains LAU 90 and LAU 60 at 25 and 27 days, respectively, while the parental strain (LAU 09), at 31 days, had the longest culture cycle (Table 4). Total fresh mushroom production was 580 g (LAU 09) to 1366g (LAU 90), the distribution pattern of production by harvest was similar for all strains, with more than 80% of the total mushroom yield produced in the first two harvests. The highest BE and PR values were recorded for strains LAU 90 (97.57, 3.9%), LAU 60 (82.57, 3.06%), LAU 30 (72.29, 2.68%) and LAU 120 (57.21, 1.97%), while least values of 41.43% (BE) and 1.34% (PR) were recorded for LAU 09 (Table 5).
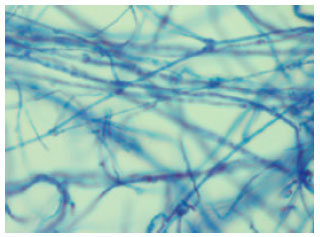 | |
| Fig. 2: | Hyphal arrangement of LAU 09 (wild) |
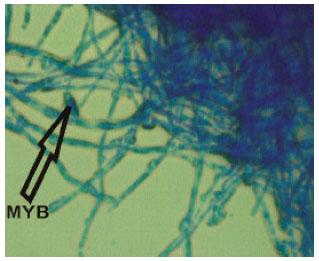 | |
| Fig. 3: | Hyphal arrangement of LAU 90 (mutant) with a prominent mycelia bud (MYB) |
| Table 6: | The differences in amino acid compositions between wild and mutant strains of P. pulmonarius |
 | |
| TAP: Type of amino acids produced, NA: No. of amino acids produced, IA: Initial amino acid, TA: Terminal amino acid, D: Aspartic acid, N: Asparagine, E: Glutamic acid | |
The morphological arrangements of the wild and mutant hypha of P. pulmonarius are shown in Fig. 2 and 3. Comparing wild and mutant strains, LAU 90 (mutant) has significant features that differentiate it from LAU 09 (wild), such features are a prominent mycelia bud structure, appendage filament and hyphal density (thickness). Table 6 shows differences in amino acid composition obtained by nucleotides sequence translation between wild and mutant strains of P. pulmonarius. The type of amino acids produced did not vary among the strains, but there is variation in the number of amino acids produced. The highest number (56) of amino acid was obtained by LAU 09 (wild), while the least number of amino acids (44) was produced by LAU 90 (mutant).
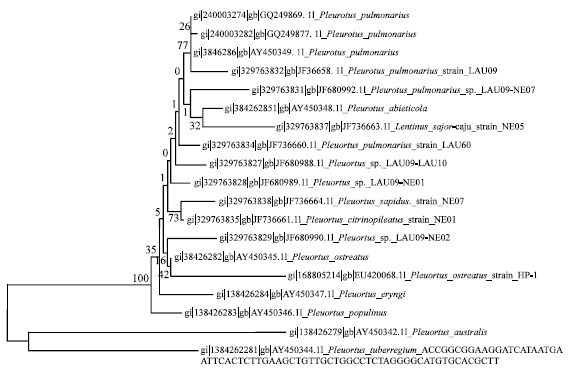 | |
| Fig. 4: | Phylogenetic tree of LAU 09 (wild) constructed by UPGMA, with boostrap values from 100 replicates showing maximum similarity with LAU 09 (wild) and the homogeneous strains of Pleurotus species |
The initial amino acid (IA) is aspartic acid (D) for LAU 09 and LAU 60, while LAU 90 (mutant) has glutamic acid (E) as the initial residue. The terminal amino acids obtained for LAU 09 and LAU 60 is the same (Asparagine-N), while LAU 90 (mutant) has D (aspartic acid).
The phylogenetic analyses using sequence data of ITS 5.8 and 28 sec ribosomal RNA gene from LAU 09 (wild) and LAU 60 (mutant) of P. pulmonarius and the corresponding GenBank data of related species are shown in Fig. 4 and 5. The outgroup arising from the root of the phylogenetic tree for P. pulmonarius (accession number: JF736658) for phylogenetic evaluation is P. tuberregium (AY450344) from the GenBank database. P. pulmonarius LAU 09 formed a phylogenetic cluster with P. pulmonarius complex at the top of the phylogenetic tree with a bootstrap value of 77%, which confirmed the close evolutionary lineage of the strain (Fig. 4). P. pulmonarius LAU 09 (accession number: JF736658) was used as an outgroup to root the trees. The rooted UPGMA tree shows two phylogenetic lineages (i.e two clusters of genes) arising from the root. One leads to a terminal node containing a strain of Lentinus sajor-caju from GenBank database (accession number: JF736663) and the other subdivides further into eight terminal branches leading to P. pulmonarius LAU 60 (mutant), which is at subcluster eight from the root. The bootstraps (100% support) values of the LAU 60 is 85% to the outgroup strain (LAU 09), which is the parent strain (Fig. 5).
The edited nucleotide sequence of LAU 90 (mutant) show no similarity with any available data in the GenBank database after blasting with the national centre for Biotechnology information (NCBI) website. The nucleotide sequence of LAU 09 (wild) and LAU 90 (mutant) are shown in Fig. 6 and 7 with total number of sequence nucleotides very high (1150) in LAU 09 (wild) compare to 750 obtained in LAU 90 (mutant).
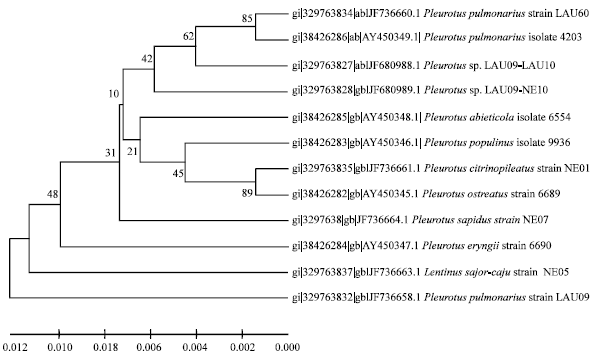 | |
| Fig. 5: | Phylogenetic tree of LAU 60 (mutant) constructed by UPGMA, with boostrap values from 100 replicates showing maximum similarity with LAU 09 (wild) and the homogeneous strains of Pleurotus species |
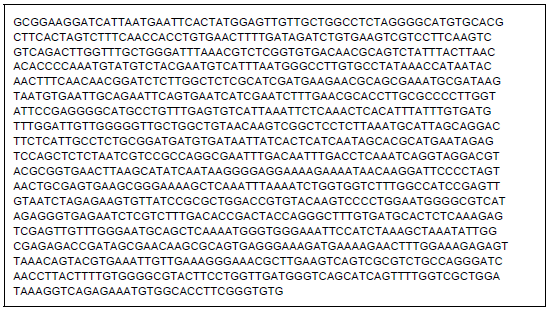 | |
| Fig. 6: | Total length (1150) of nucleotides sequence of LAU 09 (wild) |
DISCUSSION
The better survival rate obtained in mutant strains between 30 to 120 min exposure to UV-light with highest mycelia growth by LAU 90 (mutant) agree with Jaivel and Marimuthu (2010) previous report, which stated that an exposure of the parent strain of Aspergillus terreus to UV-light above 120 min reduced the survival rate of the organism, while exposure below 60mins might not really have effect on the morphological and genetic makeup of the organism.
 | |
| Fig. 7: | Total length (750) of nucleotides sequence of LAU 90 (mutant) |
In previous studies, the optimum temperature for mycelia growth rate for Pleurotus species was established at 25, -30°C, with no growth observed at 35°C (Zervakis et al., 2001; Kibar and Perksen, 2008). The considerable high yield of mycelia obtained at 35°C provides proof that the mutants strain has the ability to tolerate high range of temperature and subsequently increases the chances of the organism to perform better in tropical region. The broad ranges of optimal pH values (pH 3.5 to 6) were reported for fungal growth and products yield (Membre et al., 1999; Restaino et al., 1983). The highest yield obtained above the earlier established pH optima is an indication that the mutant strains can adapt to a high condition of salt concentration and enhanced better performance in salty environment.
The means of spawn ramification rates, weight of mycelia ramification and percentages spawn productivity showed significant differences (p<0.05) in LAU 90 from all other strains. This has established true strain improvement in comparison to wild strain (LAU 09) and a previous report. Gaitan-Hernandez and Salmones (2008) reported the average 20-30days for spawn ramification for the Pleurotus species. Zervakis et al. (2001) observed pinhead formation after an incubation period of 23 days in commercial cultures of P. ostreatus whereas Obodai et al. (2003) cited incubation periods of 34 days when using rice straw as substrate. This is commercially desirable since it represents a significant reduction in the total number of culture cycle days (Gaitan-Hernandez and Salmones, 2008).
The values of BE and PR obtained in this study (41.43-97.57 and 1.34-3.9%), respectively were higher than the values reported by Membre et al. (1999), who observed BE values of 16.8-71.9% and PR values of 0.48-1.38%. The BE and PR values by Philippoussis et al. (2001) were lower than the reported values in the present study, while Gaitan-Hernandez and Salmones (2008) reported a range of means values of BE (84.17%) and PR (1.76%) close to those reported in the present study. The highest BE and PR reported in this study, especially in mutants strain has established an increased in yield performance of the mutated strains of P. pulmonarius LAU 09, which is evidence of strain improvement.
The wide variation obtained in the mutant strain (LAU 90), in terms of hyphal arrangement developed some features such as bud-like structure, appendages filament and hyphal density that differ from the wild type (LAU 09).This provides evidence of the diversity in morphological arrangement of mutant strains as a result of the mutation effect. Several different mutations have been reported in L. edodes, e.g. those inducing nutrient deficiency (Murakamu and Tsuneda, 1982), sporeless hymenium (Hasebe, 1991), haploplasic lethal factors (Hasebe, 1991) and abnormal colony or basidiocarp morphology (Murakami and Takemam, 1975; Hasebe, 1991).
The results obtained in this study might be as result of point mutations in the nucleotide sequences which has altered the amino acid sequence and eventually will affect the protein synthesis of the organisms.
The cladistic position of the P. pulmonarius (LAU 09) strain in the phylogenetic trees suggests that there is a common ancestor and that sub ancestors diverged majorly at early in evolution (Vilgalys et al., 1996). The phylogenetic analyses based on the PCR-RFLP data of the partial 26srDNA has also been reported (Bao et al., 2004) which revealed that 9 of the biological species, the P. cornucopiae complex, P. cystidiosus complex, P. salmoneos-tramineus, complex, P. dryinus, P. nebrodensis, P. smithii and P. ulmarius were congruent with independent phylogenetic lineages. The cladograms based on the sequence of the ITS 5.8s and 28s rDNA constructed by UPGMA, NJ and parsimony methods showed that the P. pulmonarius (LAU 60) and P. pulmonarius (LAU 09) diverged at an earlier stage of evolution (Vilgalys et al., 1996), but have a close origin with other strains of P. pulmonarius (AY450349) which came together in a single phylogenetic cluster (Bao et al., 2005). The implication is that the conserved region of the mutant strain (P. pulmonarius-LAU 60) has been altered which resulted in genetic diversity in the mutant strain and caused its genetic variation from the parent (P. pulmonarius-LAU 09). This assumption is supported by a bootstrap value of 85% in the parsimomy tree. Bao et al. (2005) used RFLP analysis of 26 sec rDNA with a combination of mt SSU rDNA sequence analysis to the Pleurotus test strain identification of biological species in the genus Pleurotus.
The length of nucleotides sequence and the arrangement of the nucleotides in the mutant strain (LAU 90) is widely altered. The aberration observed in the P. pulmonarius LAU 90 (mutant) is a result of induced mutation, such as the shorter length of nucleotides (750) in comparison with the wild type which has 1150 nucleotides, there are different point mutations such as additions or deletions of the genes, which could be responsible for the high rate of yields and products production. One potential difficulty with this type of alteration is that simple insertion of circular transforming DNA into a homologous region leads to a duplication of genomic sequences. In some fungi, such duplications have been shown to be unstable and subject to extensive methylation and point mutation (Selker et al., 1987; Cambareri et al., 1989). This could also be the reason of no resemblance in nature of the strain (LAU 90) to either parent (LAU 09) or any other strains from the GenBank databases. This could have resulted in a new or novel strain of genus the of Pleurotus.
CONCLUSION
The result obtained shows the improved performance of the mutant strains over the wild (parent) strains, especially the mutant at 90mins exposure (LAU 90). The mutant strains gave the highest yield in term of spawn productivity, total weight of fresh fruit body, biological efficiency and production rate. So also, the morphological, molecular variation and differences in amino acid composition with evolutionary divergence of the mutant strains from the wild type is true evidence of a novel strain of Pleurotus species developed from this study.
ACKNOWLEDGMENT
The Director of NEIST, India is gratefully acknowledged for granting facilities available to carry out this research, so also TWAS, Italy and CSIR for the award of Postgraduate Fellowship given to me and utilized at NEIST, CSIR, Jorhat, India and also to the authority of LAUTECH, Ogbomoso, Nigeria for granted the study leave to utilized the award.
REFERENCES
- Bao, D., S. Kinugasa and Y. Kitamoto, 2004. The biological species of Oyster mushrooms (Pleurotus spp.) from Asia based on mating compatibility tests. J. Wood Sci., 50: 162-168.
CrossRef - Bao, D., T. Aimi and Y. Kitamoto, 2005. Cladistic relationship among the Pleurotus ostreatus complex, the Pleurotus pulmonarius complex and pleurotus emyngii based on the mitochondrial small submit ribosomal DNA sequences analysis. J. Wood Sci., 51: 77-82.
CrossRef - Cambareri, E.B., B.C. Jensen, E. Schabtach and E.U. Selker, 1989. Repeat-induced G-C to A-T mutations in Neurospora. Science, 244: 1571-1575.
CrossRef - Elliott, T.J. and F.A. Langton, 1981. Strain improvement in the cultivated mushroom Agaricus bisporus. Euphytica, 30: 175-182.
CrossRef - Felsenstein, J., 1989. PHYLIP-phylogeny inference package (Version 3.2). Cladistics., 5: 164-166.
Direct Link - Gaitan-Hernandez, R. and D. Salmones, 2008. Obtaining and characterizing Pleurotus ostreatus strains for commercial cultivation under warm environmental conditions. Sci. Hort., 118: 106-110.
CrossRef - Gardes, M. and T.D. Bruns, 1993. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol., 2: 113-118.
CrossRefDirect Link - Hasebe, K., 1991. Genetics studies on mutants and agronomic characters in shiitake, Lentinus edodes. Rep. Tottori Mycol. Inst., 29: 1-69.
Direct Link - Jaivel, N. and P. Marimuthu, 2010. Strain improvement of Aspergillus terreus for increased lovastatin production. Int. J. Eng. Sci. Tech., 2: 2612-2615.
Direct Link - Kang, S.W., E.H. Ko, J.S. Lee and S.W. Kim, 1999. Overproduction of beta-glucosidase by Aspergillus niger mutant from lignoocellulosic biomass. Biotechnol. Lett., 21: 647-650.
Direct Link - Kibar, B. and A. Peksen, 2008. Modelling the effects of temperature and light intensity on the development and yield of different Pleurotus species. Agric. Trop. Subtrop., 41: 68-73.
Direct Link - Stajic, M., J.B. Vukojevic and S. Duletic-Lausevic, 2007. Influence of the cultivation conditions on ligninolytic enzyme production in Pleurotus pulmonarius. Proc. Nat. Sci., 113: 303-312.
Direct Link - Membre, J.M., M. Kubaczka and C. Chene, 1999. Combined effects of pH and sugar on growth rate of Zygosaccharomyces rouxii, a bakery product spoilage yeast. Applied Environ. Microbiol., 65: 4921-4925.
Direct Link - Murakamu, S. and A. Tsuneda, 1982. Adenine-requiring mutant of Lentinus edodes, extremely susceptible to attack by Trichoderma species. Rep. Tottori Mycol. Inst., 20: 54-62.
Direct Link - Obodai, M., J. Cleland-Okine and K.A. Vowotor, 2003. Comparative study on the growth and yield of Pleurotus ostreatus mushroom on different lignocellulosic by-products. J. Ind. Microbiol. Biotechnol., 30: 146-149.
CrossRefPubMedDirect Link - Philippoussis, A., G. Zervakis and P. Diamantopoulou, 2001. Bioconversion of agricultural lignocellulosic wastes through the cultivation of the edible mushrooms Agrocybe aegerita, Volvariella volvacea and Pleurotus spp. World J. Microbiol. Biotechnol., 17: 191-200.
CrossRef - Ravishankar, S., M. Pandey, R.P. Tewari and V. Krishna, 2006. Development of sporeless/low sporing strains of Pleurotus through mutation. World J. Microbiol. Biotechnol., 22: 1021-1025.
CrossRef - Selker, E.U., E.B. Cambareri, B.C. Jensen and K.R. Haack, 1987. Rearrangement of duplicated DNA in specialized cells of Neurospora. Cell, 51: 741-752.
PubMed - Suntornsuk, W. and Y.D. Hang, 1994. Strain improvement of Rhizopus oryzae for production of l(+)-lactic acid and glucoamylase. Lett. Applied Microbiol., 19: 249-252.
CrossRef - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Vilgalys, R. and M. Hester, 1990. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol., 172: 4238-4246.
Direct Link - Zervakis, G., A. Philippoussis, S. Ioannidou and P. Diamantopoulou, 2001. Mycelium growth kinetics and optimal temperature conditions for the cultivation of edible mushroom species on lignocellulosic substrates. Folia Microbiol. (Praha), 46: 231-234.
CrossRefDirect Link