
Pramod Kumar Singh
School of Applied Sciences, Babu Banarasi Das University, 226028, Lucknow, India
Neelam Singh Verma
Department of Botany, DAV College, Kanpur, India
Neetu Pandey
School of Applied Sciences, Babu Banarasi Das University, 226028, Lucknow, India
Prashant Singh
Department of Chemistry, BBDNIIT, 226028, Lucknow, India
Research Journal of Medicinal Plants
Year: 2015 | Volume: 9 | Issue: 8 | Page No.: 375-394







ABSTRACT
To assess the salt tolerance mechanism in aromatic plants, a soil pot experiment was conducted on Ocimum sanctum (shyama tulsi). The plants were raised on natural sodic soils having different levels of Exchangeable Sodium Percentage (ESP) i.e.,: 4.37 (control), 15.7 (low), 35.6 (medium) and 54.5 (high) ESP levels in complete randomized design. Results indicated that increased in ESP significantly decreases vegetative growth and dry matter yields of the plant. The pH and EC of cell sap and proline content were increased than the control while water potential decreases (became more negative). Activities of enzymatic and some non-enzymatic anti-oxidant like catalase (CAT), peroxidase (POD), superoxide dismutase (SOD) and total phenol content were increased while Glutathione Reductase (GR) and H2O2 content were reduces that imply oxidative stress as a consequences increased soil ESP. The concentration of chloroplastic pigments like chlorophylls, carotenoids and carotenoids/chlorophyll ratio were decreased on increasing ESP. The reducing sugars and total carbohydrate were also decreased on increasing ESP. The protein content and essential oil content were increased on increasing the ESP levels. The Na concentration in root and stem of plant were increased while in leaf it was decreased. The K concentration was decreased on increasing the soil sodicity in root while in stem and leaf it was increased. The Na/K ratio was decreased on increasing the soil sodicity in stem and root while in leaf it was decreased. The concentration of iron (Fe), manganese (Mn), zinc (Zn) copper (Cu) and lead (Pb) were observed maximum in leaves which increases in low and medium ESP than the control while in high ESP these were decreases except Zn. Thus Ocimum plant showed tolerance mechanism in sodic stress environment.
PDF Abstract XML References Citation
Received: August 04, 2015;
Accepted: September 22, 2015;
Published: October 10, 2015
How to cite this article
Pramod Kumar Singh, Neelam Singh Verma, Neetu Pandey and Prashant Singh, 2015. Soil Sodicity Induced Changes in Aromatic Plants: Effects on Growth, Water Relation, Photosynthetic Pigments, Antioxidative Enzymes, Cations Concentration and Quality of Ocimum sanctum. Research Journal of Medicinal Plants, 9: 375-394.
URL: https://scialert.net/abstract/?doi=rjmp.2015.375.394
URL: https://scialert.net/abstract/?doi=rjmp.2015.375.394
INTRODUCTION
Exposure of plants to abiotic stresses such as high salinity and sodicity, drought, extreme light and temperature leads to major loss in crop worldwide. Among the various abiotic stress factors, soil salinization is the biggest threat to inland agriculture. In arid and semi arid parts of the world the problem of soil salinity and sodicity is prevalent to considerable extent (Yadav, 1993). A study on global land use pattern reveals that 7% of the world’s land area, amounting to 1000 million ha, has become saline (Tester and Davenport, 2003). In the present scenario human use of poor- quality irrigation systems is a major concern for scientists around the world. Therefore apart from the need for proper irrigation practices a concerted effort to understand the effect of salinity on plants, development of genetically engineered crop varieties and superior tolerant cultivars are essential to combat the world’s salinization problems (Rengasamy, 2006). The detrimental effects of alkalinity and salinity on growth, dry matter production and grain yield of different crops have been reported by several workers (Bhatnagar and Yadav, 1980; Singh and Abrol, 1985; Abrol, 1986; Manchanda et al., 1985; Singh and Singh, 1990; Chodhary et al., 1996; Dharmendra and Bijendra, 1997; Khanduja et al., 1980). Salinity stress affects plant growth, as well as development process such as seed germination, seedling growth and vigour, vegetative growth, flowering and fruit set (Sairam and Tyagi, 2004). In salt affected plants the result is primarily an ionic imbalance and hyper osmotic stress. The effect of this imbalance or disruption in homeostasis occurs at the cell level as well as at the whole-plant level. Massive change in ionic and water balance cause molecular damage and growth arrest. Finally in extreme saline conditions, this leads to tissue death and ultimately death of plant (Zhu et al., 1997; Xiong and Zhu, 2002). It is evident from the growing body of experimental evidence that salt stress affects the integrity of cellular membranes, activities of enzymes and the functioning of the plant photosynthetic apparatus (Serrano et al., 1999). An important cause of this damage is production of Reactive Oxygen Species (ROS) (Smirnoff, 1993). The ROS is routinely generated during normal plant metabolic processes. Chloroplast, mitochondria and peroxysomes are the most important intracellular generators of ROS like O2 and H2O2. Amongst these, chloroplast-mediated O2 and H2O2 production remains the most unavoidable consequence of an oxygen-enriched atmosphere. In chloroplasts, O2 and H2O2 are mainly produced by the electron acceptor of photosystem I, where as singlet oxygen is generated by the transfer of the electron from an excited chlorophyll molecules to molecular oxygen (Asada and Takahashi, 1987; Kikuzaki and Nakatani, 1993).
The present endeavor is aimed to evaluate and screened out the sodicity tolerance mechanism of Ocimum sanctum L. (shyama tulsi) which is held sacred by Hindus and is used as medicinal plants in day to day practice in Indian homes for various ailments (Rajeshwari, 1992). The purpose of study to reduce ever rising pressure to produce more food, fiber, fodder and other agricultural commodities, it is difficult to spare fertile land for cultivation of aromatic crops. Several medicinal properties have been attributed to Ocimum sanctum L. (Batta and Santhakumari, 1971; Sarkar et al., 1994). Different parts of Tulsi plant e.g., leaves, flowers, stem, root, seeds etc. are known to possess therapeutic potentials and have been used, by traditional medical practitioners, as expectorant, analgesic, anticancer, antiasthmatic, antiemetic, diaphoretic, antidiabetic, antifertility, hepatoprotective, hypotensive, hypolipidemic and antistress agents. Tulsi has also been used in treatment of fever, bronchitis, arthritis, convulsions etc. Aqueous decoction of Tulsi leaves is given to patients suffering from gastric and hepatic disorders (Rajeshwari, 1992; Pandey and Anita, 1990). Herbal preparations containing Ocimum sanctum L have been suggested to shorten the course of illness, clinical symptoms and biochemical parameters in patients suffering from viral hepatitis (Rajeshwari, 1992). The leaf juice of Ocimum sanctum L. along with Triphala is used in Ayurvedic eye drop preparations recommended for glucoma, cataract, chronic conjunctivitis and other painful eye diseases. The juice of fresh leaves is also given to patients to treat chronic fever, dysentery, hemorrhage and dyspepsia (Pandey and Anita, 1990). A decoction of Tulsi leaves is a popular remedy for cold (Rajeshwari, 1992; Pandey and Anita, 1990). Tulsi leaves also check vomiting and has been as anthelmintic (Sen, 1993).
Thus, in present study Ocimum species were selected for the screening of tolerance limit and tolerant species of Ocimum on the basis of morpho-physiological changes, ions uptake and essential oil production for cultivation in sodic stress environment.
MATERIALS AND METHODS
Plant materials and experimentation: Among the plants known for medicinal value, the plants of genus Ocimum belonging to family Lamiacae are very important for their therapeutic potentials. Ocimum sanctum L. known as ‘Tulsi’ in Hindi and ‘Holy Basil’ in English, is an erect aromatic herb or under shrub found throughout India. Two types of Ocimum sanctum L. are met within cultivation: (i) Tulsi plants with green leaves known as Sri Tulsi and (ii) Tulsi plants with purple leaves known as Krishna Tulsi (Pandey and Anita, 1990).
The plants were raised on natural sodic soils having different levels of exchangeable sodium percentage (ESP) i.e., 4.37 (control), 15.7 (low), 35.6 (medium) and 54.5 (high) ESP levels with four replicates of each in complete randomized design. The study was conducted in earthen pots to know the tolerance limit of plant at different ESP levels at which plant can easily grow and reproduce. The soil were collected from the five sites of the Banthra village up to 15 cm depth (surface soils) only and kept it for drying. After drying the soils were thoroughly crushed and properly mixed. The soils were analyzed after processing and chemical properties of soil were indicated in the Table 1. The earthen clay pots were lined on their inner side alkathene sheets to check the leaching and contamination from the clay of the pots of 15 kg soils were filled in each pot, 5 plantlets of Ocimum sanctum (Shyama Tulsi) were transplanted in mid July and thinning as done after 30 days to allow 4 plants to grow in each pot. A does treatment of 200 mg N kg–1 soil as Ca (NO3)2 and 100 mg P kg–1 soils as KH2PO4 and 100 mg kg–1 soil as MgSO4 was applied to all plant in two split doses during crop growth.
Growth, cell sap pH and EC, plant water relation: Data were recorded as plant height and dry matter yield (oven dried at 70°C for 24 h) and seed yield. Plant height was measured from soil level to top of inflorescence. The water status of leaf tissue was ascertained when plants were about 100 days old. At the same time cell sap pH and EC of leaf samples were determined using EC and pH meter after homogenization of one gram of fresh leaves in 20 mL of 0.025 M EDTA solution (Dwivedi et al., 1980). The water potential was measured hydrometrically in leaves using Wescor micro-voltmeter and C-52 leaf chambers when the plants were growing at field capacity. The Water Saturation Deficit (WSD) and Specific Water Content (SWC) were calculated following standard procedures (Barrs and Weatherley, 1962).
| Table 1: | Chemical properties of soil used in experiment |
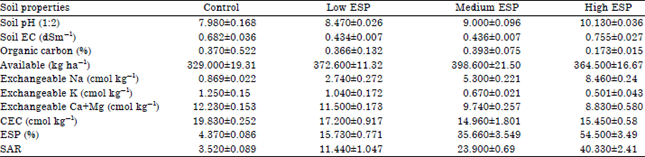 | |
| EC: Electrical conductivity, CEC: Cation exchange capacity, ESP: Exchange sodium percentage, SAR: Sodium absorption ratio | |
Yield and cations analysis: Yield was recorded at harvest. Harvested plants were washed and thoroughly separated into root, stem and leaves and dried in an oven at 70°C for 24 h. Tissue concentration of cations were measured in the solution after wet digestion (HNO3, HCLO4 10:1v/v mixture) of the oven dried plant material (Piper, 1967). The cations (Na and K) were measured on flame photometer while Fe, Mn, Zn, Cu and Pb were measured on atomic absorption spectrophotometer (Perkin Elmer Analyst 300).
Chloroplastic pigments: Concentration of chlorophylls and carotenoids were determined in 80% acetone extract of the young fully expanded fourth leaf by the method of Lichtehthaler (1987). The homogenate was centrifuged at 4000x g for 10 min to remove the residue. The color intensity of clear supernatants were measured at 663.2; 646.8 and 470 nm for chlorophyll a, chlorophyll b and carotenoids, respectively. Results have been expressed as mg chlorophyll or carotenoids g–1 fresh weight.
Enzyme extraction: Fresh leaf tissue (2.5 g) was homogenized in 10.0 mL of chilled 50 mM potassium phosphate buffer (pH 7.0) containing 1.0% insoluble Polyvinyl Pyrrolidone (PVP) using chilled pestle and mortar kept in ice bath. The homogenate was filtered with two-fold muslin cloth and centrifuged at 20000x g for 10 min in refrigerated centrifuge at 2°C. The supernatant was stored at 2°C and used for enzyme assays within 4 h.
Enzyme assays: The various enzymes were assayed in crude leaf extracts made in glass-distilled water. Finely chopped leaf material was ground with chilled acid washed silica sand in an ice-chilled pestle and mortar kept in an ice bath. One gram of leaf tissue was extracted in 10 mL of chilled GDW and diluted properly at the time of enzyme assay. The tissue extract was filtered through a double fold muslin cloth kept at 5°C in refrigerate till the time of enzymes assay.
Catalase (EC, 1.11.1.6): Catalase was assayed by the method of Bisht (1972), a modification of the method of Euler and Josephson. Ten milliliter of reaction mixture, standardized against 0.1 M potassium permagnate, containing 0.5 mM hydrogen peroxide and 1 mM phosphate buffer (pH 7.0), was taken in a test tube stabilized at 25°C. The reaction was initiated by adding 1 mL of suitably diluted enzyme extract and the contents were mixed thoroughly. The reaction was allowed to proceed for five minutes and then stopped by addition of 2 mL of 4 M H2SO4. Corresponding blanks in which sulphuric acid was added prior to the addition of enzyme extract, were run simultaneously. The final reaction mixture was then titrated against 0.1 M KMnO4. The H2O2 reduced was then calculated as the difference in titrate value of respective blank and sample. Enzyme activity is expressed as μmole H2O2 reduced unit fresh matter or protein weight.
Peroxidase (EC. 1.11.1.7): Modification of the method by Luck (1963) was used for peroxidase assay. Reaction mixture comprising of 5 mL 0.1 M phosphate buffer (pH 6.0) 1 mL 0.01% H2O2 and 1 mL 0.05% p-phenylene diamine was taken in a centrifuge tube and stabilized at 25°C. Reaction was initiated by adding 1 mL of diluted enzyme extract to the reaction and allowed to proceed for 5 min after which it was stopped by adding 2 mL of 4 N H2SO4. Corresponding blanks were maintained in which H2SO4 was added to the substrate mixture prior to the addition of enzyme extract. These tube kept in refrigerator for 20-30 min and then contents centrifuge at 4000x g for 15 min at room temperature. The color intensity of the supernatant was measured at 485 nm on spectrophotometer (Spectro-chem MK II Manufactures AIML). The enzyme activity has been expressed as units 100 mg–1 fresh weight protein, an enzyme unit being defined as the difference of 0.01 in the optical density between blank and sample per minute of reaction time.
Superoxide dismutase (EC 1.15.1.1): The activity of superoxide dismutase (SOD) was determined by measuring the ability to inhibit the photochemical reduction of Nitro-Blue Tetrazolium (NBT) in a 3 mL reaction mixture containing 50 mM phosphate buffer pH 7.8, 13 mM methionine, 75 μM NBT, 2 μM riboflavin, 0.1 mM EDTA and 0 to 50 μL enzyme extract. Riboflavin was added last and tubes were illuminated for 10 min. Blanks were not illuminated and the above reaction mixture without the enzyme extract developed the maximum colour at 560 nm. One unit of SOD represents the amount that inhibits the NBT reduction by 50%. The Cu/Zn SOD was determined by inhibiting the activity by 3 mM KCN and subtracting the value from total SOD. The enzyme has been quantified on the basis of % inhibition (Beauchamp and Fridovich, 1971).
Glutathione reductase (EC 1.6.4.2): Glutathione Reductase (GR) assay was performed in a 3 mL reaction mixture containing 100 mM phosphate buffer pH 7.0, 1 mM GSSG, 1 mM EDTA, 0.1 mM NADPH and 25-50 μL of the enzyme extract. The oxidation of NADPH was followed by monitoring the decrease in absorbance per min at 340 nm. The amount of NADPH oxidized was calculated using the extinction coefficient of 6.22 mM–1 cm–1 (Jablonski and Anderson, 1978).
Hydrogen peroxide: Hydrogen peroxide (H2O2) was estimated by method of Brennan and Frenkel (1977). Freshly chopped leaf tissues were ground in chilled pestle mortar in 100% chilled acetone and centrifuged at 10,000 g for 5 min. The pellet was discarded. The supernatant was mixed with water and shaken thoroughly. To the supernatant, titanium tetra chloride was added followed by chilled liquid ammonia. The precipitate was solubilized with the help of a thin glass rod and centrifuged. Residue was washed repeatedly with acetone to remove chlorophyll. The colorless residue was dissolved in 2 M H2SO4. The color intensity was read at 415 nm. The results have been expressed as μmol H2O2 g–1fresh weight.
Total phenol contents: Total Phenol content was determined using folin-cioltue reagent. The leaf extract was shaken thoroughly and made up to 10 mL using distilled water. The amount of total phenolic compound was determined colorimetrically with Folin Ciolcalteu (FC) reagent and 1.5 mL of 20% sodium carbonate according to earlier studies (Kumaran and Karunakaran, 2007). The absorbance of resulting solution was measured at 765 nm in UV-spectrophotometer.
Proline content: Proline was estimated by the method of Bates et al. (1973). Freshly chopped leaves were ground in sulphosalicylic acid. After filtration with Whatman filter paper No.1, a suitable aliquot was taken with ninhydrin reagent and glacial acetic acid and boiled for 1 h. The color was extracted in toluene and read at 520 nm.
Soluble protein: Determination of protein in tissue extracts was made to determine the specific activity of enzyme. Protein was precipitated in tissue extracts with an equal volume of 20% chilled Trichloro Acetic Acid (TCA) and tissue-TCA mixture was allowed to stand for 4 h at 4°C. The content were centrifuge and the residue washed with 5% TCA. Glass distilled water was finally used to removed TCA. This residue was then freed of the pigment by extracting thrice with acetone. The residue was then dried in vacuum. The dried residue so obtained was solubilized in 0.1 M NaOH at 80°C for 10 min on water bath and protein was estimated by folin Ciolcalteu reagent of Lowry et al. (1951). The color intensity was measured in spectronic colorimeter at 660 nm. The readings were referred to calibration curve prepared from crystalline serum albumin from Sigma chemical Co. St Louis USA. The results have been expressed on percent fresh weight basis.
Sugars: Sugars were determined colorimetrically by the method of Nelson (1944). A suitable aliquot drawn from the alcohol soluble fraction was treated with a saturated solution of lead acetate and filtered. The residual lead in the filtrate was removed by precipitation with supersaturated solution of potassium oxalate. The filtrate was made to volume and a suitable aliquot drawn from this was used for determination of the concentration of the reducing sugars. Total (reducing plus non-reducing) sugars were estimated as reducing sugars after carrying out the enzymatic hydrolysis of the non-reducing sugars (using invertase) for 24 h. The non-reducing sugars were calculated as the difference in the concentration of sugars before and after hydrolysis by invertase.
Starch: Starch was estimated by the method of Montgomery (1957). The alcohol insoluble material was first solubilized in distilled water in a boiling water bath for 15 min. It was macerated with the help of a glass rod in 25% perchloric acid (PCA). The supernatant was separated by centrifugation at 5,000 g for 10 min and collected in 50 mL of 15% PCA and centrifuged. The process was repeated with 10 and 5% PCA. Finally, the residue was rejected and all the supernatant fractions collected were combined and made to volume. To a suitable aliquot from this was added 0.2 mL of 80% phenol. This was followed by rapid addition of 5 mL concentrated sulphuric acid. The solution was allowed to cool at room temperature and the color developed during the reaction period was read at 490 nm using 1 cm cell and blank reagent setting of 0.0. Starch concentration has been expressed as percentage per unit fresh weight.
Carbohydrates: For the estimation of sugar and starch plant material was fixed in boiling 80% ethanol in the proportion of 1:10. The fixed material was crushed in a pestle and mortar. Alcohol soluble and insoluble fractions were separated by repeated centrifugation at 8,000 g. In the alcohol soluble fractions, sugar was estimated while alcohol insoluble fraction was used for the determination of starch.
Essential oil contents and chemical constituents: Essential oil content was determined by hydro-distillation (Langenau, 1948) of the powdered leaves in Clevenger type apparatus (1928), for 4 h, from which v/w.
![]()
Statistical analysis: Statistical method and test of significance appropriate to the design were applied to the data for discriminating the treatment effects. To elucidate the nature and magnitude of effect, the level of significance was calculated by one way ANOVA and `F` test in the analysis of variance (Panse and Sukhatme, 1967).
RESULTS
Plant height and dry matter yields: Results indicate that plants height significantly decreased at high ESP in comparison to control while at low and medium ESP it was non-significant (Fig. 1a). The percent decreased in plant height was more steep at higher ESP levels which was 28.4% decrement was observed while at lower and medium ESP level it was non-significant. The number of branches decreased non-significantly on increasing ESP (Fig. 1b).
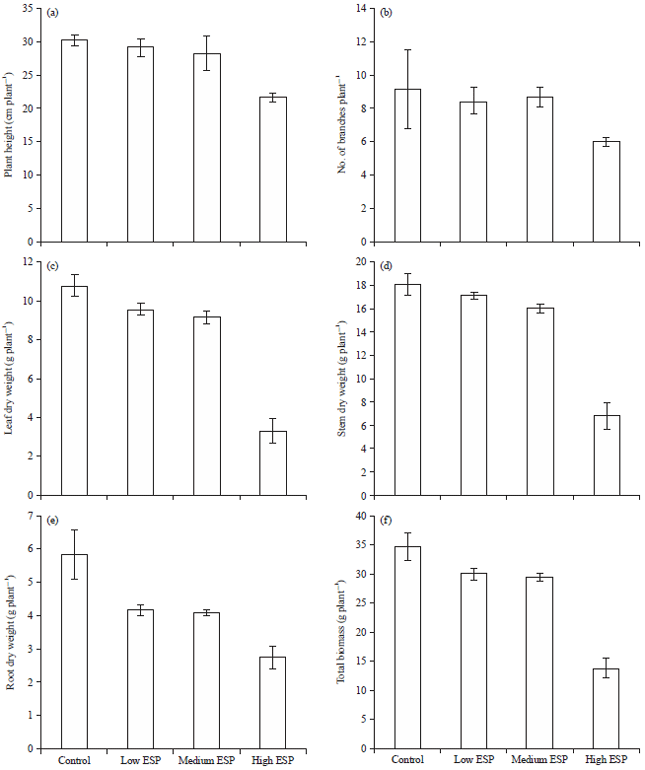 | |
| Fig. 1(a-f): | Effect of soil sodicity on vegetative growth and dry matter yields of Ocimum sanctum, (a) Plant height, (b) No. of branches, (c) Leaf dry weight, (d) Stem dry weight, (e) Root dry weight and (f) Total biomass |
Leaf and stem dry weight were decreased significantly in comparison with control while at lower ESP level it was non-significant in both (Fig. 1c-f). Root dry weight was also significantly decreased on increasing the ESP levels.
Cell sap pH, Electrical Conductivity (EC) and water contents: The cell pH and EC were significantly decreased on increasing the soil sodicity (Fig. 2a and b). Relative Water Contents (RWC) was observed highest in control (93.39%) and it was decreased on increasing ESP levels but it was non-significant decreased (Fig. 2c). The Specific Water Content (SWC) was maximum in control (5.386) and minimum in lower ESP (Fig. 2d).
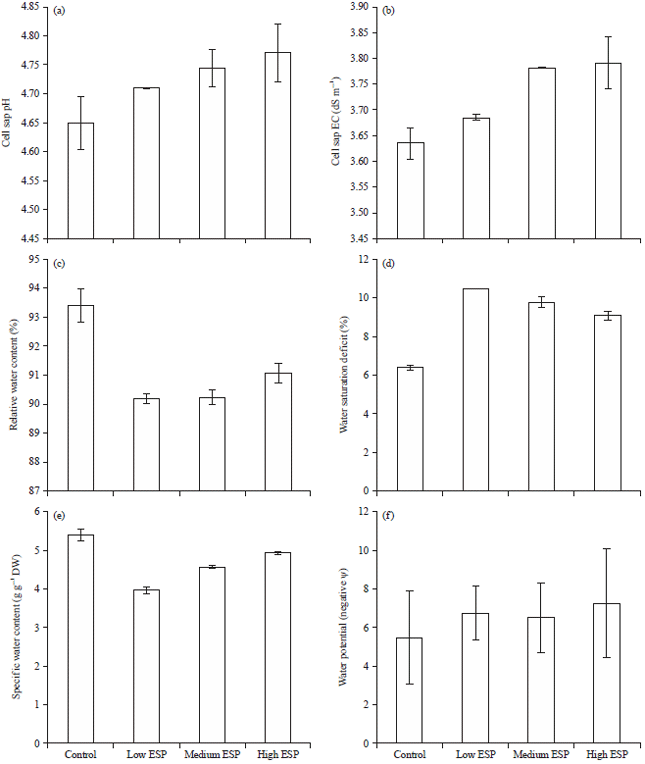 | |
| Fig. 2(a-f): | Effect of soil sodicity on (a) Cell sap pH, (b) Cell sap EC, (c) Relative water content, (d) Water saturation deficit, (e) Specific water content and (f) Water potential of Ocimum sanctum plant |
The SWC was decreased on increasing the ESP level. Comparison of control with lower ESP levels it was significant decrease observed but comparison with medium and it was non-significant. Comparison of lower ESP level with high ESP the SWC was significant increased. The water saturation deficit (WSD) was maximum at lower ESP (10.503) and minimum in control (6.393). The WSD was non-significantly increased on increasing the ESP levels (Fig. 2e). The water potential was decreased (became more negative) significantly on increasing soil sodicity (Fig. 2f).
Photosynthetic pigments: Photosynthetic pigment i.e., chl ‘a’, chl ‘b’ and total chlorophyll were significantly decreased on increasing the soil sodicity. Carotenoid content was significantly decreased at low and medium ESP levels in comparison to control while at high ESP levels it was also significant decreased than the control. The carotenoid/chlorophyll ratio was significantly decreased in low and high ESP but slightly increased in medium ESP than the control. There was no clear trend was observed in it (Table 2).
Enzymatic antioxidants: The antioxidant activities i.e., catalase, peroxidase, SOD and glutathione reductase activities were significantly increases on increasing soil sodicity. In case of catalase activity was prominently visible at medium and high ESP level while in low ESP it was no significant than the control (Fig. 3a). Peroxidase activities was significant at high ESP while in low and medium ESP level it was non significant (Fig. 3b). The SOD activity was significantly increases at all ESP levels (Fig. 3c). While glutathione reductase activity was significantly decreases on increasing the soil ESP (Fig. 3d). The difference in the mean values among the treatment groups are greater than the expected, there was statistically significant difference (p = 0.003).
Proline, total phenol and hydrogen peroxide (H2O2) contents: Non-enzymatic antioxidant like proline and total phenol contents were significantly increases on increasing the soil sodicity while H2O2 content was significantly decreased (Fig. 3e-g). Proline is stress protein which accumulation in the plant showed tolerance against stress. There was a statistically significant difference (p = 0.001).
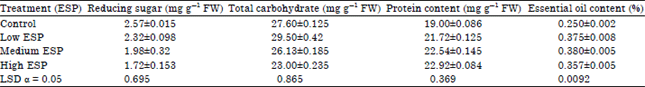
Reducing sugar, total carbohydrate, protein content and essential oil content: The reducing sugar was significantly decreased on increasing the soil sodicity while total carbohydrate was significantly increased in low ESP than the control while in medium and high ESP it was decreased. Soluble protein content in the leaf tissue was significantly increased on increasing the soil sodicity but it compare within the treatment low and medium ESP level were higher than the high ESP.
| Table 2: | Effect of different levels of soil sodicity on the photosynthetic pigments and its ratio in the leaf of Ocimum sanctum |
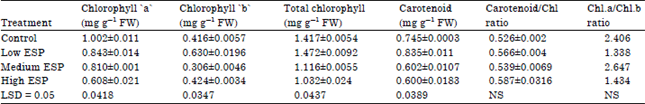 | |
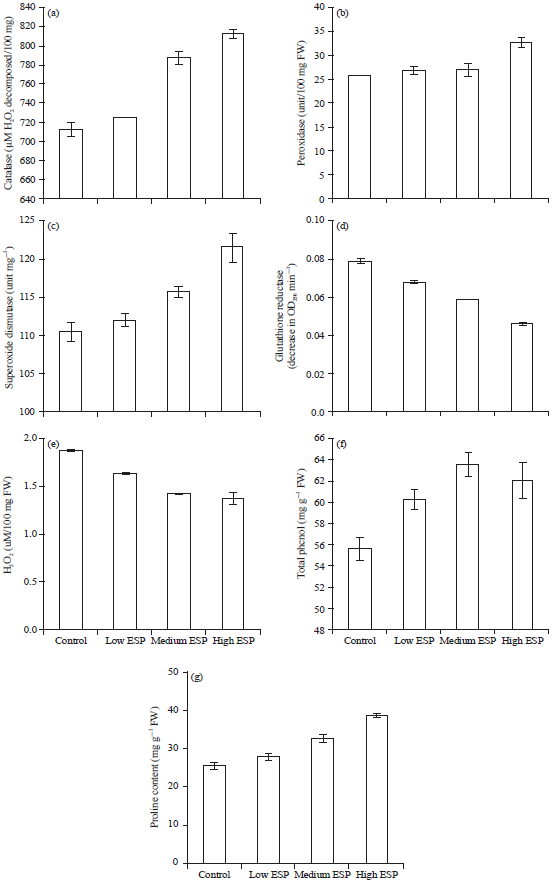 | |
| Fig. 3(a-g): | Effect of soil sodicity on antioxidative enzymes (a) Catalase, (b) Peroxidase, (c) Superoxide dismutase, (d) Glutathione reductase, (e) H2O2, (f) Total phenol and (g) Proline contents in the leaves of Ocimum sanctum plant |
| Table 3: | Effect of different levels of soil sodicity on the quality (i.e., reducing sugar, total carbohydrates, protein and essential oil content in leaf) of Ocimum sanctum |
 | |
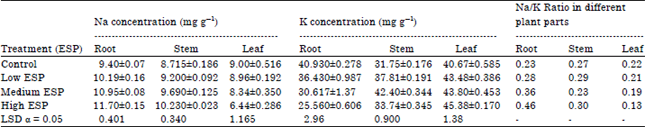
| Table 4: | Effect of different levels of soil sodicity on the sodium (Na) and potassium (K) concentration and Na/K ratio in different plant parts of Ocimum sanctum |
 | |
Essential oil content was significantly increased on increasing the sodicity. Essential oil content was in the range of 0.25-0.38%. Maximum oil content was observed in medium ESP while minimum in control (Table 3).
Cations concentration
Na and K concentration: The Na concentration in root and stem were significantly increased than the control on increasing the soil sodicity while in leaf it was non-significantly decreased in low ESP and medium ESP (Table 4). The K concentration was significantly decreased in root on increasing soil sodicity while in stem and leaf it was significantly increased. Although in stem maximum K concentration (42.42 mg g–1) was observed at medium ESP level and minimum (31.75 mg g–1) was in control while in leaves maximum concentration of K was observed at high ESP level and minimum in control (Table 4). The Na/K ratio was maximum in the root in comparison to stem and leaf. In root, it was increased on increasing the soil sodicity. Same trend was also observed in stem where Na/K ratio increases on increasing the sodicity. While in leaf Na/K ratio was decreases than the control on increasing the sodicity which just reverse to root and stem.
Fe, Mn, Zn, Cu and Pb concentration: The Fe, Mn, Zn, Cu and Pb concentrations in different plant parts were significantly affected by soil sodicity (Fig. 4). The Fe concentration ranged between 251-730 μg g–1 in different plant parts. In root, Fe content was significantly increased at low and medium ESP level but at high ESP level it was significantly decreased. In stem it was non-significant increment while in leaf significant decrease was observed. The Mn concentration was varied 18-49.5 μg g–1 in different plant parts. In root, no any trend was observed. In stem and leaf Mn concentration was significantly increased on increasing soil sodicity. The Zn concentration in different plant parts were ranged 46.3-93.8 μg g–1 in different plant parts. There was no trend observed in root while in stem and leaf it was significantly decreased on increasing soil sodicity. The Cu concentration in different plant parts were ranged from 10.3-27.23 μg g–1. In root Cu concentration was non-significantly decreased in low ESP but it was significantly decreased in medium and high ESP than the control. Pb concentration in different plant parts were ranged from 65.83-102.6 μg g–1 in different plant parts.
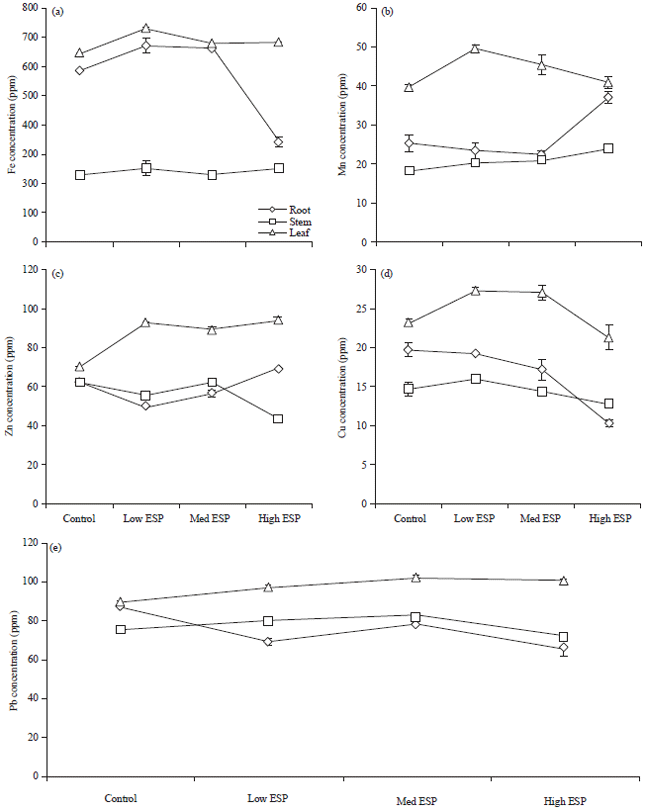 | |
| Fig. 4(a-e): | Effect of soil sodicity on cations content, (a) Fe, (b) Mn, (c) Zn, (d) Cu and (e) Pb concentrations in the leaves of Ocimum sanctum plant |
In root, Pb content was significantly decreased than control on increasing the sodicity while in stem it was significantly increased in low and medium ESP than the control but in high ESP it was significantly decreased. In leaf, Pb concentration was significantly increased than the control on increasing the sodicity.
DISCUSSION
Increasing soil sodicity profoundly affected the survival, growth and metabolic process of Ocimum species. The detrimental effects of increased ESP were distinct from transplanting to harvest. Plant height was significantly decreases on increasing the ESP levels but maximum detrimental effect was observed at high ESP (ESP 54.4) level and in low and medium ESP levels Ocimum was tolerant because decreased was non-significant. Similar finding was observed in other parameters i.e., number of branches per plant, root, stem and leaf dry weight per plant and total biomass in which detrimental effect was observed at high ESP but in low and medium ESP levels it was non-significant. Similar finding was also observed by Singh et al. (2014) and Garg et al. (2005) in fennel plant who have reported that no detrimental effect was observed upto 25 ESP . The detrimental effects of alkalinity and salinity on growth, dry matter production and grain yield of different crops have been reported by several workers (Bhatnagar and Yadav, 1980; Singh and Abrol, 1985; Abrol, 1986; Manchanda et al., 1985; Singh and Singh, 1990; Chodhary et al., 1996; Dharmendra and Bijendra, 1997; Khanduja et al., 1980) who have conducted experiment at graded level of soil ESP on grapevine. Qadar (1995) observed that the increasing sodicity stress adversely affected growth with delayed initiation of tillering and reduced grain filing period in rice (Oryza sativa). Dua (1998) observed six characters (plant height, number of tillers/plant, number of pods/plant and days to 50% flowering and 100-seed weight and grain yield/plant) in Vicia faba 27 genotypes grown under normal, saline and sodic soil conditions. Rajpar and Wright (2000) reported that the grain yield and straw dry weight per plant of spring wheat (Triticum aestivum) cv. Kharchia-65 were not affected up to 20 ESP however at (39-40 ESP) were decreased. Garg et al. (2000) indicated the potential of growing coriander and fennel in moderate sodic soils. Thus, Ocimum plant can survive up to 35 ESP without detrimental effect in the present study.
The decrease in dry matter production may also due to decreased photosynthetic capacity of plants. Photosynthetic efficiency is related to the photosynthetic pigments of plants growth at high ESP, which showed decrease in our study. In our study both forms of chlorophyll ‘a’ and ‘b’ along with carotenoids declined such decreased in chlorophyll a and b contents as a result of increase in soil sodicity which is also observed by several workers (Singh et al., 2014; El-Sharkawi et al., 1986; Tewari and Singh, 1991). This reduction in chlorophyll content may be related to the enhanced activity of chlorophyllase (Reddy and Vora, 1986).
In our study, the photosynthetic pigment e.i., chl ’a’ and total chlorophyll were significantly decreased on increasing soil sodicity. The carotenoid content was significantly decreased. The carotenoid/chlorophyll ratio was significantly decreased in low and high ESP but slightly increased in medium ESP than the control. There was no clear trend was observed in it. Similar finding was also observed by other worker (Singh et al., 2014). Reduction in chlorophyll content plants treated with NaHCO3 was more than in those treated with Na2CO3 during early growth of maize (Garg and Garg, 1982). Reddy and Das (1978) observed accumulation of Na and Cl in the chloroplast of groundnut (Arachis hypogaea) in response to salinity (NaCl) and only of Na in alkalinity (Na2 CO3) was more detrimental. The conditions adversely affected Hill reaction activity, photophoshorylation (starch and sugars) and protein nitrogen in Arachis hypogaea chloroplast was noted under both salinity (NaCl) and sodicity (Na2CO3) and there was an increase in soluble nitrogen. Huang et al. (1995) reported that salinity that salinity reduced photosynthesis rate stomatal conductance and leaf chlorophyll content in summer squash (Cucurbita pepo). Wang et al. (1999) studied that Amaranthus tricolor plants to salt stress (0.3 M NaCl) for 5 days and observed that the level of chlorophyll and activity of PSI, PSII, ribulose-1,5-biphosphate carboxygenase and the CO2 fixation rate decreased by about 20-35% in the green region whereas those in the non-green regions remained unaffected.
Oxygen radicals are generated during plant metabolism and they need to be scavenged for maintenance of normal growth. Large evidence has accumulated from various plant systems showing the environmental stresses, especially drought and salt stress, alter the amounts and the activities of enzymes involved in scavenging oxygen radicals. In the ascorbate-glutathione or Halli-Well-Asada cycle (Asada, 1994). Two key enzymes are assumed to deal with the detoxification of reactive oxygen species in plants. The primary scavenger is the enzyme superoxide to hydrogen peroxide. This toxic products of SOD is eliminated by ascorbate per oxidase, at the expense of oxidizing ascorbate to monodehydroascorbate. Both enzymes exist as several isozymes and are active in the chloroplast and the cytosol (Asada, 1994).
In the present study, the antioxidative enzymes i.e., the catalase, peroxidase and SOD activities were significantly increased on increasing the sodicity while glutathione reductase activity was decreased. That showed tolerance against sodic stress environment. It is suggested that the inactivation of catalase was possibly due to enhancement of H2O2 levels. Similar finding was observed that SOD activity was unregulated after NaCl treatment and the product of this reaction is H2O2 (Parida et al., 2004). Similar results are observed in a different species B. gymnorrhiza. In this study, catalase-activity assays were done before and after salt stress treatments in 500 mM NaCl stress medium for 9 days. It was shown that catalase activity increased to about 4.9 times more that in unstressed control plants. In a different study, NaCl tolerance in Suaeda nudiflora callus cultures was evaluated to investigate if alterations in antioxidant activity occur at cell level (Cherian and Reddy, 2003). Plant growth was retarded with respect to both fresh and dry mass with increase in NaCl concentration. Maximum reductions of 92 and 94% were found for fresh and dry mass at 200 mM NaCl. There was 60% decrease in total catalase activity. These experiments showed the declining capacity of callus tissues to scavenge H2O2 with increasing salt stress. In another study, total catalase activity decreased with increasing NaCl concentration in Crithmum maritimum (Amor et al., 2005). Though studies on catalase in mangroves are few, these clearly showed a decrease in activity. It is possible that catalases respond similarly to salt stress conditions in mangroves and halophytes as in other glycophytes. Plants contain abundant Amounts Of Peroxidases (POX) that are also involved in H2O2 scavenging. They are associated with the cell wall where they generate phenoxy compounds from cinnamic acids. The POX are also involved in to her plant defense mechanisms, including responses to insect (Salin, 1988; Bi and Felton, 1995) and in a coordinated response known as the oxidative burst (Kawano, 2003). Similar finding was also observed by different workers (Cherian et al., 1999; Amor et al., 2005; Cherian and Reddy, 2003). Increased POX activity was observed in root and shoot tissues in Avicennia marina in comparison to control under NaCl stress conditions; however, no significant changes in POX activity were observed in leaf tissues (Cherian et al., 1999). Enhanced activity of POX was also observed in callus cultures of Suaeda nudiflora Moq at higher salt concentrations. The NaCl-induced enhancement of POX activity in salinized cells of S. nudiflora indicated that these cells had a higher capacity for decomposition of H2O2 and that POX perform a major role in this particular reaction (Cherian and Reddy, 2003). A transient increase of POX activity was also observed in halophytes like Crithmum maritimum at different NaCI concentrations studied (Amor et al., 2005).
Superoxide dismutase (SOD) is an important antioxidant enzyme and is the first line of defense against oxidative stress in plants. The SOD causes dismutation of superoxide radicals at almost diffusion-limited rates to produce H2O2 (Salin, 1988). It plays an important part in determining the concentration of O2 and H2O2 in plants and hence performs a key role in the defense mechanism against free-radical toxicity (Bowler et al., 1992). Similar trend were also observed by several workers. According to him salt stress in cultivars differing in salt tolerance has revealed increased SOD activity in salt-tolerant genotypes of pea, cotton and tomato and induction of SOD enzyme activity was suggested as a reason for improved tolerance to salinity in these cases (Hernandez et al., 1993, 1995, 1999; Gosset et al., 1994; Mittova et al., 2003). Steep increase in total SOD activity levels has been recorded in Bruguiera gymnorrhiza and B. parviflora during salt stress (Takemura et al., 2000; Parida et al., 2004). When B. gymnorrhiza seedlings were placed in 500 mM NaCl for 9 days, the total SOD activity increased by almost 8.1 times in comparison to unstressed control (Takemura et al., 2000). Again, analysis of the total SOD activity in B. parviflora seedlings subjected to 45 days of NaCl stress at 200 and 400 mM NaCl revealed an increase of 128% (Parida et al., 2004). In Avicenna marina it was observed that the total SOD activity increased in leaf tissues after six weeks of application of salt stress (Cherian et al., 1999). According to Cherian and Reddy (2003), the presence of a strong antioxidative response mechanism combined with physiological specialization in the plant contribute to the difference in salt tolerance capacity between whole plants and calli in S. nudiflora. Thus, Ocimum plant also showed antioxidative response against sodic stress environment.
In addition to antioxidative enzymes, nonenzymatic antioxidants also play a key role in scavenging free radicals in plants during salinity stress. The nonenzymatic component comprises molecules such as ascorbic acid, glutathione, tocopherol and carotenoids that can scavenge ROS (Arora et al., 2002). Reduced glutathione is a powerful reductant and hence a very efficient scavenger of ROS (May et al., 1998). Carotenoids scavenge free radicals that are generated owing to excess excitation energy from chlorophyll during photosynthesis (Arora et al., 2002). Osmolytes play an important functional role in osmotic adjustment under hyper osmotic stress caused by salinity stress (Yancey et al., 1982). Osmolytes are synthesized as a metabolic response to salt stress and include algras, polyols, amino acids and tertiary and quaternary ammonium and sulphonium compounds (Chen and Murata, 2002).
Proline is a stress protein which accumulation in the plant showed tolerance against stress. In the present study, which indicated that proline content accumulation was increased on increasing the sodicity in O. sanctum Maximum proline content was observed in the plant grown in high ESP while minimum in control. Similar finding also prove the finding which indicates that the mechanism of action of compatible solutes in conferring protection against free radicals during stress conditions is still largely unknown (Chen and Murata, 2002). However, recent studies suggest a link between increased osmolyte content (e.g., proline) in halophytes with decreased ROS production and better tolerance to salt stress. The amino acid proline is a good osmoticum and helps in membrane stability (Hanson and Burnet, 1994). A correlation was also shown for increased proline content and improved tolerance to abiotic stress factors (Matysik et al., 2002). Increased levels of proline accumulation have also been observed in salt-stressed calli of Suieda nudiflora. The study suggested that proline protects the callus cells from membrane damage caused by during salt stress (Cherian and Reddy, 2003). In plants, proline also scavenges singlet oxygen and free-radical-induced damages and performs an important role in protection of proteins against denaturation (Alia et al., 1991).
Total phenol content was significantly increased than the control on increasing the sodicity in O. sanctum while it was non-significant on comparison with low ESP to medium and high ESP. Sugar alcohols like pinitol also play an important role in intracellular osmotic adjustment as well as in scavenging free radicals. Pinitols accumulate in response to salinity stress in Mesembryanthemum crystallinum myo-Inositol O-methyltransferase (IMT) is the principal enzyme in the pinitol biosynthesis pathway (Vernon and Bohnert, 1992).
Qualitative aspect showed that reducing sugars significant decrease was observed than the control on increasing sodicity while total carbohydrate was significantly increased on than the control on increasing sodicity. Protein content in the leaf tissue was significantly increased on increasing the sodicity stress. Thus it appears that in sodic stress condition reducing sugars convert in carbohydrates, on the basis of these finding we can say that O. sanctum shows maximum tolerance than the others two. Hydro-distillation results indicated that the essential oil content of O. sanctum was significantly increased on increasing the sodicity. Similar finding was also observed by Singh et al. (2014). They stated that essential oil content of fennel plant increases on increasing soil sodicity.
Exchangeable sodium caused in the absorption of sodium, nitrogen and molybdenum and a decrease in the absorption of calcium, potassium, sulfur, manganese of Ca, Mg, K in different plant parts as well as total uptake decreased with increase in exchangeable sodium in root zone of the plant has been reported by various workers (Das and Mehrotra, 1971; Abrol, 1986, Garg and Srivastava, 1986; Singh and Singh, 1990; Odar, 1990; Graifenberg et al., 1996; Botrini et al., 2000; Singh et al., 2014). A decrease in accumulation of Fe, Cu, Mn and Zn has been shown in plants expose to salt stress (Singh and Singh, 1990). The problem of Zn, Fe and Cu uptake is very acute under sodic conditions where carbonate and bicarbonate are supposed to be major anions interfering with their availability and may cause precipitation of these nutrients as insoluble salts due to high soil pH.
CONCLUSION
Thus, Ocimum sanctum showed tolerance in sodic stress environment and can be easily grown up to 35 ESP. Dry matter yield reduction in sodic stress condition appeared to be due to both soil physico-chemical changes and nutritional changes. The nutritional changes involved the combined effects of nutrient deficiency and Na toxicity.
ACKNOWLEDGMENT
The author is thankful to Department of Science and Technology (DST), New Delhi for financial assistance through SERC Fast Track Scheme (SR/FT/L-172/2004).
REFERENCES
- Alia, P.P. Saradhi and P. Mohanty, 1991. Proline enhances primary photochemical activities in isolated thylakoid membranes of Brassica juncea by arresting photoinhibitory damage. Biochem. Biophys. Res. Commun., 181: 1238-1244.
CrossRefDirect Link - Arora, A., R.K. Sairam and G.C. Srivastava, 2002. Oxidative stress and antioxidative system in plants. Curr. Sci., 82: 1227-1238.
Direct Link - Barrs, H.D. and P.E. Weatherley, 1962. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci., 15: 413-428.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link - Amor, N.B., K.B. Hamed, A. Debez, C. Grignon and C. Abdely, 2005. Physiological and antioxidant responses of the perennial halophyte Crithmum maritimum to salinity. Plant Sci., 168: 889-899.
CrossRefDirect Link - Bhatnagar, C.P.B. and J.S.P. Yadav, 1980. Effect of varying levels of soil salinity and alkali on growth and nutrient uptake of barley. Agrokemia Talajtan, 29: 447-455.
Direct Link - Bi, J.L. and G.W. Felton, 1995. Foliar oxidative stress and insect herbivory: Primary compounds, secondary metabolites and reactive oxygen species as components of induced resistance. J. Chem. Ecol., 21: 1511-1530.
CrossRefDirect Link - Huang, B., D.S. NeSmith, D.C. Bridges and J.W. Johnson, 1995. Responses of squash to salinity, waterlogging and subsequent drainage: I. Gas exchange, water relations and nitrogen status. J. Plant Nutr., 18: 127-140.
CrossRefDirect Link - Botrini, L., A. Graifenberg and M.L. di Paola, 2000. Potassium affects sodium content in tomato plants grown in hydroponic cultivation under saline-sodic stress. HortScience, 35: 1220-1222.
Direct Link - Bowler, C., M.V. Montagu and D. Inze, 1992. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Mol. Biol., 43: 83-116.
CrossRefDirect Link - Brennan, T. and C. Frenkel, 1977. Involvement of hydrogen peroxide in the regulation of senescence in pear. Plant Physiol., 59: 411-416.
Direct Link - Chen, T.H.H. and N. Murata, 2002. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol., 5: 250-257.
CrossRefPubMedDirect Link - Cherian, S. and M.P. Reddy, 2003. Evaluation of NaCl tolerance in the callus cultures of Suaeda nudiflora Moq. Boil. Plant., 46: 193-198.
Direct Link - Cherian, S., M.P. Reddy and J.B. Pandya, 1999. Studies on salt tolerance in Avicennia marina (Forstk.) Vierh.: Effect of NaCl salinity on growth, ion accumulation and enzyme activity. Indian J. Plant Physiol., 4: 266-270.
Direct Link - El-Sharkawi, H.M., F.M. Salame and A.A. Mazen, 1986. Chlorophyll response to salinity, sodicity and heat stresses in cotton, rama and millet. Photosynthetica, 20: 204-211.
Direct Link - Garg, V.K., P.K. Singh and P. Pushpangadan, 2005. Exchangeable sodium induced changes in yield, water relation and cation composition of fennel (Foeniculum vulgare Mill). J. Environ. Biol./Acad. Environ. Biol. India, 26: 335-340.
PubMedDirect Link - Gosset, D.R., E.P. Millhollon and M.C. Lucas, 1994. Antioxidant response to NaCl stress in salt-tolerant and salt-sensitive cultivars of cotton. Crop Sci., 34: 706-714.
CrossRefDirect Link - Graifenberg, A., L. Botrini, L. Giustiniani and M.L. Di Paola, 1996. Salinity affects growth, yield and elemental concentration of fennel. HortScience, 31: 1131-1134.
Direct Link - Hernandez, J.A., A. Campillo, A. Jimenez, J.J. Alarcon and F. Sevilla, 1999. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol., 141: 241-251.
CrossRefDirect Link - Hernandez, J.A., F.J. Corpas, M. Gomez, L.A. Rio and F. Sevilla, 1993. Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria. Physiol. Plant., 89: 103-110.
CrossRefDirect Link - Hernandez, J.A., E. Olmos, F.J. Corpas, F. Sevilla and L.A. del Rio, 1995. Salt-induced oxidative stress in chloroplasts of pea plants. Plant Sci., 105: 151-167.
CrossRefDirect Link - Jablonski, P.P. and J.W. Anderson, 1978. Light-dependent reduction of oxidized glutathione by ruptured chloroplasts. Plant Physiol., 61: 221-225.
PubMedDirect Link - Kawano, T., 2003. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep., 21: 829-837.
CrossRefPubMedDirect Link - Khanduja, S.D., K.N. Chaturvedi and V.K. Garg, 1980. Effect of exchangeable sodium percentage on the growth and mineral composition of Thompson Seedless grapevines. Scientia Horticulturae, 12: 47-53.
CrossRefDirect Link - Kumaran, A. and R.J. Karunakaran, 2007. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. LWT-Food Sci. Technol., 40: 344-352.
CrossRefDirect Link - Kikuzaki, H. and N. Nakatani, 1993. Antioxidant effects of some ginger constituents. J. Food Sci., 58: 1407-1410.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Manchanda, H.R., S.K. Sharma and J.P. Singh, 1985. Effect of increasing levels of residual sodium carbonate in irrigation water on the exchangeable sodium percentage of a sandy loam soil and crop yield. J. Indian Soc. Soil Sci., 33: 366-371.
Direct Link - Matysik, J., Alia, B. Bhalu and P. Mohanty, 2002. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci., 82: 525-532.
Direct Link - May, M., T. Vernoux, C. Leaver, M. van Montagu and D. Inze, 1998. Glutathione homeostasis in plants: Implications for environmental sensing and plant development. J. Exp. Bot., 49: 649-667.
CrossRefDirect Link - Mittova, V., M. Tal, M. Volokita and M. Guy, 2003. Up-regulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii. Plant Cell Environ., 26: 845-856.
CrossRefDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Odar, A., 1990. Effect of seedling age of two rice varieties on their sodicity tolerance. Indian J. Plant Physiol., 33: 162-164.
Direct Link - Parida, A.K., A.B. Das and P. Mohanty, 2004. Defense potentials to NaCl in a mangrove, Bruguiera parviflora: Differential changes of isoforms of some antioxidative enzymes. J. Plant Physiol., 161: 531-542.
CrossRefPubMedDirect Link - Qadar, A., 1995. Requirement of rice crop for phosphorus and potassium at varying sodicity levels. J. Plant Nutr., 18: 2291-2303.
CrossRefDirect Link - Rajpar, I. and D. Wright, 2000. Effects of sowing method on survival, ion uptake and yield of wheat (Triticum aestivum L.) in sodic soils. J. Agric. Sci., 134: 369-378.
Direct Link - Reddy, M.P. and A.B. Vora, 1986. Changes in pigment composition, Hill reaction activity and saccharides metabolism in Bajra (Pennisetum typhoides S & H) leaves under NaCl salinity. Photosynthetica, 20: 50-55.
Direct Link - Rengasamy, P., 2006. World salinization with emphasis on Australia. J. Exp. Bot., 57: 1017-1023.
CrossRefDirect Link - Sairam, R.K. and A. Tyagi, 2004. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci., 86: 407-421.
Direct Link - Salin, M.L., 1988. Toxic oxygen species and protective systems of the chloroplast. Physiol. Plant., 72: 681-689.
CrossRefDirect Link - Sarkar, A., S.C. Lavania, D.N. Pandey and M.C. Pant, 1994. Changes in the blood lipid profile after administration of Ocimum sanctum (Tulsi) leaves in the normal albino rabbits. Indian J. Physiol. Pharmacol., 38: 311-312.
PubMedDirect Link - Serrano, R., J.M. Mulet, G. Rios, J.A. Marquez and I.F. de Larrinoa et al., 1999. A glimpse of the mechanisms of ion homeostasis during salt stress. J. Exp. Bot., 50: 1023-1036.
CrossRefDirect Link - Dharmendra, S. and S. Bijendra, 1997. Influence of residual sodium carbonate in sodic water on the yield, oil content and oil production of lemon grass (Cymbopogon cytratus). J. Indian Soc. Soil Sci., 45: 354-357.
Direct Link - Singh, P.K., P. Kumar and P.K. Tandon, 2014. Soil sodicity alters antioxidative enzymes, photosynthetic pigments, water content and essential oil quality of fennel (Foeniculum vulgare Mill.). Res. J. Soil Biol., 6: 1-16.
CrossRefDirect Link - Abrol, I.P., 1986. Effect of soil sodicity on growth yield and chemical composition of soybean. J. Indan Soc. Soil Sci., 34: 568-571.
Direct Link - Singh, S.B. and M.V. Singh, 1990. Effect of exchangeable sodium on the growth, yield and mineral composition of barley varieties. J. Indian Soc. Soil Sci., 38: 135-138.
Direct Link - Singh, S.B. and I.P. Abrol, 1985. Effect of exachangeable sodium percentage on growth, yield and chemical composition of onion and garlic. J. Indian. Soc. Soil Sci., 33: 358-361.
Direct Link - Smirnoff, N., 1993. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol., 125: 27-58.
CrossRefDirect Link - Takemura, T., N. Hanagata, K. Sugihara, S. Baba, I. Karube and Z. Dubinsky, 2000. Physiological and biochemical responses to salt stress in the mangrove, Bruguiera gymnorrhiza. Aquatic Bot., 68: 15-28.
CrossRefDirect Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link - Tewari, T.N. and B.B. Singh, 1991. Stress studies in lentil (Lens esculenta Moench). II. Sodicity-induced changes in chlorophyll, nitrate and nitrite reductase, nucleic acids, proline, yield and yield components in lentil. Plant Soil, 136: 225-230.
CrossRefDirect Link - Vernon, D.M. and H.J. Bohnert, 1992. A novel methyl transferase induced by osmotic stress in the facultative halophyte Mesembryanthemum crystallinum. EMBO J., 11: 2077-2085.
PubMedDirect Link - Wang, Y., Y.L. Meng, H. Ishikawa, T. Hibino, Y. Tanaka, N. Nii and T. Takabe, 1999. Photosynthetic adaptation to salt stress in three-color leaves of a C4 plant Amaranthus tricolor. Plant Cell Physiol., 40: 668-674.
Direct Link - Xiong, L. and J.K. Zhu, 2002. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ., 25: 131-139.
CrossRefPubMedDirect Link - Yadav, I.S.P., 1993. Salt affected soils and their management with special reference to Uttar Pradesh. J. Indian Soc. Soil Sci., 41: 623-629.
Direct Link - Yancey, P.H., M.E. Clark, S.C. Hand, R.D. Bowlus and G.N. Somero, 1982. Living with water stress: Evolution of osmolyte systems. Science, 217: 1214-1222.
CrossRefPubMedDirect Link - Zhu, J.K., P.M. Hasegawa, R.A. Bressan and H.J. Bohnert, 1997. Molecular aspects of osmotic stress in plants. Crit. Rev. Plant Sci., 16: 253-277.
CrossRefDirect Link - Batta, S.K. and G. Santhakumari, 1971. The antifertility effect of ocimum sanctum and hibiscus rosa sinensis. Indian J. Med. Res., 59: 777-781.
PubMedDirect Link