
H. Mahmoodzadeh
Departmentof Biology, Faculty of Science, Islamic Azad University, Mashad Branch, Mashad, Iran
F. Abbasi
Departmentof Biology, Faculty of Science, Islamic Azad University, Mashad Branch, Mashad, Iran
S. Rohani
Departmentof Biology, Faculty of Science, Islamic Azad University, Mashad Branch, Mashad, Iran
Research Journal of Environmental Sciences
Year: 2010 | Volume: 4 | Issue: 4 | Page No.: 407-413







ABSTRACT
In vitro seeds germination and plantlet establishment of Zinnia elegans were studied in this report. The seeds of Z. elegans were sterilized and cultured on Murashige and Skoog (MS) solid media supplemented with 1-3 μM of 6-benzylaminopurine (BAP) or Kinetin (KIN) and 3% (w/v) sucrose. The presence of KIN in the medium significantly affects seeds germination. High percentage of seeds germination (up to 90%) was successfully achieved after 3 days of culture on medium supplemented with 1 μM KIN. The average of 3.8 leaves per explant obtained on media treated with 3 μM BAP. Result showed that medium supplemented with 2 μM KIN promote the highest growth with an average of 2.72 cm shoot length and 1.6 cm root length after 3 weeks. In addition, medium added with 2 μM BAP and supplemented with 3-4% (w/v) of sucrose promote the best growth i.e., 8.6 mm leaf length. In conclusion, present results showed that it is possible to improve the production of Zinnia elegans plants in vitro with using different cytokinins and carbon sources.
PDF Abstract XML References Citation
How to cite this article
H. Mahmoodzadeh, F. Abbasi and S. Rohani, 2010. In vitro Germination and Early Seedling Growth of Zinnia elegans. Research Journal of Environmental Sciences, 4: 407-413.
DOI: 10.3923/rjes.2010.407.413
URL: https://scialert.net/abstract/?doi=rjes.2010.407.413
DOI: 10.3923/rjes.2010.407.413
URL: https://scialert.net/abstract/?doi=rjes.2010.407.413
INTRODUCTION
Zinnia elegans (Asteraceae) is the most well known of the 20 or so species in the Zinnia genus. The wild form is a coarse, upright, bushy annual, to 30 in (76 cm) high, with solitary daisylike flowerheads on long stems and opposite, sandpapery, lance shaped leaves. The ray flowers are purple, the discs yellow and black and the entire head is about 2 in (5 cm) across. Gardeners love zinnias and there are at least a hundred cultivars in a diversity of flower colors and types, some with flowerheads up to 6 in (15 cm) across. There are zinnias with white, cream, green, yellow, apricot, orange, red, bronze, crimson, purple and lilac flowers; zinnias with striped, speckled and bicolored flowers; zinnias with double, semi-double and dahlia-like pompon flowers; zinnias that range from dwarfs that don't exceed 6 in (15 cm) in height to cut flower beauties that get 3 ft (0.9 m) tall. Newer varieties are resistant to powdery mildew and other diseases. Old Mexico is like the wild plant with single, daisylike flowerheads with wide purple rays (www.floridata.com).
Several researchers have noted certain advantages of using seeds that is, intact seedlings as primary explants (Malik and Saxena, 1992a, b; Victor et al., 1999). If this method is followed, quiescent embryonic cells are directly exposed to a hormonal stimulus, dedifferentiation of parenchymatous cells and callus induction are bypassed, the seedlings’ integrity is retained and wounding is avoided.
Cytokinins promote cell division and cell expansion in plant cell culture. In clonal propagation of tropical fruit trees, growth is stimulated by adding a cytokinin to a proliferation medium. Many researchers have been reported suitable cytokinin types and their concentration for fruit crop species.
The activity of various cytokinins in morphoge-netic phenomena reflects their concentration in the tissue, which depends on the capacity for their synthesis and the activity of various inactivation mechanisms. The most important role in cytokinin inactivation is assigned to the enzyme cytokinin oxidase/dehydrogenase (CKX) (Galuszka et al., 2001; Bilyeu et al., 2003), formerly described as cytokinin oxidase (Galuszka et al., 2000).
Neibaur et al. (2008) showed induction of highly regenerable callus with approximately 400 shoots per cultured immature inflorescence (1 cm in length) was achieved by culturing 0.2 cm segments on media with 3, 6-dichloro-2-methoxybenzoic acid (dicamba) and benzylaminopurine (BA). A multifactorial experiment demonstrated the combination of dicamba and BA for induction is suitable.
Shoot formation from seedling explants of Pinus heldreichii was induced by pulse treatment with benzyladenine at different concentration, followed by culture on medium lacking plant growth regulators. After treatment with 222 μM benzyladenine (BA) an average of 4.6 shoots per explant was obtained. Shoots, detached from explants, rooted with a frequency of about 10% and rooted plantlets were successfully transferred to ex vitro conditions (Stojicic and Budimir, 2004).
Compact Embryogenic Tissues (CET) obtained after 6-8 months of periodic subculture on Callus Proliferation Medium (CPM) and brought to liquid CPM supplemented with 0.1 mg L-1 diamba alone or in combination with 0.1, 0.5 and 1.0 mg L-1 BA or KIN. After 6 times of subculture (2-3 weeks interval) dicamba with KIN (0.1 mg L-1) gave small aggregates composed of round, dense in cytoplasm led to establishment of fine suspensions in short period of time. The two PGR were proved to play an important role in both cell viability and formation of chloroplasts in embryogenic cell suspension culture.
Callus capable of plant regeneration was initiated at a higher frequency from slices of cormel sprouts of Gladiolus primulinus cv. Golden Wave, cultured in MS with 4.0 mg L-1 NAA,2.0 mg L-1 BAP or1.0 mg L-1 2, 4-D. Callus was maintained in callus initiation medium. Shoots regenerated from the callus in MS with 0.5 - 2.0 mg L-1 KIN or/and 0.5 - 2.0 mg L-1 BAP (Sinha and Roy, 2002).
The aim of the present research was to evaluate in vitro germination and plant growth from Zinnia elegans seeds.
MATERIALS AND METHODS
Plant Material and Sterilization Method
The experiment was conducted in faculty of Science,Islamic Azad University, Mashad Branch, Iran (2008).
The seeds of Zinnia elegans were obtained from Agricultural Research Center of Mahalat (Iran). The seeds were then submerged in 100% (v/v) Clorox (5.25% sodium hypochlorite) for 15 min and finally rinsed three times with sterile distilled water to remove traces of chlorine.
Culture Media and Culture Conditions
The basal medium consisted of full strength MS basal medium (Murashige and Skoog, 1962) and supplemented with 3% (w/v) sucrose. The pH of the media was adjusted ton 5.8 prior to the addition of 0.8% (w/v) agar. To melted the agar, the media were heated in the oven and then distributed into conical flasks and autoclaved for 20 min at 121°C and 15 psi. After autoclaving, the media were allowed to cool at room temperature. All cultures were grown in a plant growth chamber at 25±2°C with 16 h photoperiod provided by 36 μmol/m2/s1 cool light fluorescent tube.
The seed coat was removed using scalpel blade and the seeds were cut longitudinally prior to culturing. The halved cotyledons with embryos were aseptically placed on 10 mL of seed germination media containing of basal media supplemented with 1, 2 or 3 μM BAP or KIN. The medium without Plant Growth Regulators (PGRs) served as a control medium.
The stages of seed germination and development were observed weekly. The seeds considered to be germinated by the emergence of radicle from the seeds. Percentage of seeds germination, numbers of leaves as well as shoot height and root length were determined after 3th week of incubation. Percentage of seed germination was calculated.
Statistical Analysis
The experiments were conducted in 10 replicates per treatment. Data collected in all experiments were analyzed by SPSS and subjected to Analysis of Variance (ANOVA). The mean differences were tested using Duncan multiple range test (DMRT) with significant value of p<0.05 (Khan et al., 2004).
RESULTS
The germination process started after 3 days of culture (Fig. 1). The elongation of epicotyls was observed within 5 to 7 days on all the germination media. Maintaining seeds in the same media up to 7 weeks resulted to formation of numerous green structures observed along epicotyls. Shoots were then elongated along with expanded leaves 2-3 weeks after culture (Fig. 2).
Based on the result summarized in Table 1, the highest percentage of germination (82.6%) was achieved in PGR-free medium followed by 82.3 or 69.6% on media supplemented with 2, 1 and 3 μM BAP, respectively. However, the presence of KIN (1 μM )has significantly promoted of embryo germination (Table 2). The highest number of leaves (3.8±0.2 leaves per explant) was observed on the medium supplemented with 3 μM BAP followed by media supplemented with 1 μM (2.83±0.2 leaves) and 2 μM BAP (2.7±0.3 leaves), respectively.
 | |
| Fig. 1: | Embryo germination of Z. elegans on MS medium |
 | |
| Fig. 2: | Elongated shoots with expanded leaves of Z. elegans on in vitro condition |
| Table 1: | Effect of BAP concentration on seed germination of Z. elegans after 1 week of culture on MS medium supplemented with BAP, pH 5.8, 3% (w/v) sucrose, 0.8% (w/v) agar and grown at 25±2°C with 16 h photoperiod |
 | |
| Values represent Mean±SD of 10 replicates. Means followed by the same superscripted letter did not differ significantly at (p<0.05) | |
| Table 2: | Effect of kinetin concentration on seed germination of Z. elegans after 1 week of culture on MS medium supplemented with KIN, pH 5.8, 3% (w/v) sucrose, 0.8% (w/v) agar and grown at 25±2°C with 16 h photoperiod |
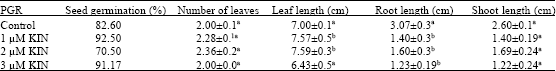 | |
| Values represent Mean±SD of 10 replicates. Means followed by the same superscripted letter did not differ significantly at (p<0.05) | |
The KIN was superior compared to BAP in term of mean shoots height and roots height per explant. The explants cultured on the medium supplemented with KIN obtained the longest shoot (2.72±0.15 cm) and the highest root height (1.6±0.6). Otherwise, BAP produced larger leaves as compared to another (Fig. 2).
DISCUSSION
Cytokinins are plant hormones implicated in the regulation of various processes of growth and development. Among their multiple activities, the effects of cytokinins on seed germination stand apart from their role in shoot morphogenesis. Modern analytical methods have shown their very active metabolism in all phases of germination, from imbibition to radicle emergence and the start of seedling establishment (Stirk et al., 2005; Chiwocha et al., 2005). On the other hand, exogenous cytokinins have various effects on seed germination in different species. Their promotive effects are mostly related to the alleviation of stress factors. Kinetin was one of the growth regulators that alleviated both innate and salinity-induced seed dormancy in many halophytes (Khan and Ungar, 1997; Khan et al., 2004). However, kinetin did not affect germination in some halo-phytic grasses (Gulzar and Khan, 2002). Kinetin was also reported to prolong the viability of recalcitrant seeds, which was interpreted as possible protection of cell membranes against oxidative stress (Chaitanya and Naithani, 1998). In the range of concentrations used in our experiments, cytokinins can be divided into two groups, according to their physiological activities: BAP exhibited rather weak effects, whereas KIN constituted a very active group. In promoting seed germination, KIN occupy the first place. The differences of leaf sizes between explant cultured on control medium and media supplemented with various cytokinins revealed the absolute requirement of cytokinins during leaf formation. During this stage, cytokinins are required to drive the cell division cycle at a normal speed and to obtain the required number of cell divisions to reach a normal leaf size. Therefore, in the absence or deficient of cytokinins, a reduced leaf size may cause mainly by a reduced rate of cell division (Werner et al., 2001).
Mackay et al. (1995) noted the poor effect of KIN on shoot growth of Cercis canadensis. Contrary to the present observation regarding the superiority of KIN, earlier report indicated that the better growth and elongation of cotyledonary node culture of Pterocarpus marsupium was obtained on BAP-supplemented medium (Anis et al., 2005). It was also found that BAP was the most effective for shoot formation among other cytokinins in the culture of Polygonatum odoratum seedling explants (Yoon and Choi, 2002). In other plant species such as Myrica esculenta, KIN showed the best performance with regards to shoots proliferation (Bhatt and Dhar, 2004).
The fact that in our experiments KIN acted as a very active cytokinin in seed germination and elongation of roots and shoots, but as a weak cytokinin in leaf formation, is hard to explain. It has frequently been reported that cytokinin action is different in different types of cells, for instance in root and shoot apical meristems (Werner et al., 2001). Differential uptake of exogenous cytokinins can be considered one of the causes of this phenomenon (Auer et al., 1999). Seed germination is most likely initiated in quiescent embryonic cells and these cells perhaps become determined at the same time for subsequent regeneration. However, the stimulus for root and shoot elongation is probably perceived in the apical meristematic cells. They may differ from embryonic cells in endogenous cytokinin content and in uptake of exogenous hormones. Spichal et al. (2004) have recently demonstrated that similar cytokinin receptors differ in recognizing cytokinins as ligands, as well as in transmitting their signals to biological processes.
CONCLUSION
In conclusion, the results presented here show that it is possible to improve the production of Zinnia elegans plants in vitro. Moreover, Zinnia elegans seeds and seedlings seem to be rewarding objects for further fundamental studies concerning the relationship between cytokinins and the systems that regulate their activities. Dry seeds containing quiescent embryonic cells represent novel objects on which to study the development of these functions.
ACKNOWLEDGMENTS
This study is financially supported by Islamic Azad University, Mashad Branch, Iran (Grant No. Ms project/1387).
REFERENCES
- Anis, M., H.M. Kashif and S. Anwar, 2005. In vitro plant regeneration of Pterocarpus marsupium Roxb., an endangered leguminous tree. Curr. Sci., 88: 861-863.
Direct Link - Auer, C.A., V. Motyka, A. Brezinova and M. Kaminek, 1999. Endoge-nous cytokinin accumulation and cytokinin oxidase activity during shoot organogenesis of Petunia hybrida. Physiol. Plant, 105: 141-147.
CrossRef - Bhatt, I.D. and U. Dhar, 2004. Factors controlling micropropagation of Myrica esculenta Buch.-Ham. Ex D. Don: A high value wild edible of Kumaun Himalaya. Afr. J. Biotechnol., 3: 534-540.
Direct Link - Bilyeu, K.D., J.G. Laskey and R.O. Morris, 2003. Dynamics of expression and distribution of cytokinin oxidase/dehydrogenase in developing maize kernels. Plant Growth Regul., 39: 195-203.
CrossRef - Chaitanya, K.S.K. and S.C. Naithani, 1998. Kinetin-mediated prolonga-tion of viability in recalcitrant sal (Shorea robusta Gaertn f.) seeds at low temperature: Role of kinetin in delaying membrane deterioration during desiccation-induced injury. J. Plant Growth Regul., 17: 63-69.
CrossRef - Chiwocha, S.D.S., A.J. Cutler, S.R. Abrams, S.J. Ambrose, J. Yang, A.R.S. Ross and A.R. Kermode, 2005. The etr1-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during maintenance of seed dormancy, moist-chilling and germination. Plant J., 42: 35-48.
PubMedDirect Link - Galuszka, P., I. Frebort and M. Sebela, 2000. Degradation of cytokinins by cytokinin oxidases in plants. Plant Growth Regul., 32: 315-327.
CrossRef - Galuszka, P., I. Frebort, M. Sebela and P. Sauer, 2001. Cytokinin oxidase or dehydrogenase? Mechanism of cytokinin degradation in cereals. Eur. J. Biochem., 268: 450-461.
PubMed - Gulzar, S. and M.A. Khan, 2002. Alleviation of salinity-induced dor-mancy in perennial grasses. Biol. Plant., 45: 617-619.
CrossRefDirect Link - Khan, M.A. and I.A. Ungar, 1997. Alleviation of seed dormancy in the desert forb Zygophyllum simplex L. from Pakistan. Ann. Bot., 80: 395-400.
Direct Link - Khan, M.A., B. Gul and D.J. Weber, 2004. Action of plant growth reg-ulators and salinity on seed germination of Ceratoides lanata. Can. J. Bot., 82: 37-42.
CrossRef - Mackay, W.A., J.L. Tipton and G.A. Thompson, 1995. Micropropagation of Mexican redbud, Cercis canadensis var. mexicana. Plant Cell Tissue Org. Cult., 43: 295-299.
Direct Link - Malik, K.A. and P.K. Saxena, 1992. Somatic embryogenesis and shoot regeneration from intact seedlings of Phaseolus acutifolius A., P. aureus L. Wilezek., P. coccineus L. and P. wrightii L. Plant Cell Rep., 11: 163-168.
CrossRef - Malik, K.A. and P.K. Saxena, 1992. Thidiazuron induces high-fre-quency shoot regeneration in intact seedlings of pea (Pisum sativum), chickpea (Cicer arietinum) and lentil (Lens culinaris). Aust. J. Plant Physiol., 19: 731-740.
CrossRef - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Neibaur, I., M. Gallo and F. Altpeter, 2008. The effect of auxin type and cytokinin concentration on callus induction and plant regeneration frequency from immature inflorescence segments of Seashore paspalum (Paspalum vaginatum Swartz). In vitro Cell. Dev. Biol. Plant, 44: 480-486.
CrossRefDirect Link - Sinha, P. and S.K. Roy, 2002. Plant regeneration through in vitro cormel formation from callus culture of Gladiolus primulinus baker. Plant Tissue Cult., 12: 139-145.
Direct Link - Spichal, L., N.Y. Rakova, M. Riefler, T. Mizuno and G.A. Romanov, 2004. Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay. Plant Cell Physiol., 45: 1299-1305.
Direct Link - Stirk, W.A., J.D. Gold, O. Novak, M. Strnad and J. van Staden, 2005. Changes in endogenous cytokinins during germination and seedling establishment of Tagetes minuta L. Plant Growth Regul., 47: 1-7.
CrossRef - Stojicic, D. and S. Budimir, 2004. Cytokinin-mediated axillary shoot formation in Pinus heldreichii. Biol. Plant., 48: 477-479.
CrossRef - Victor, J.M.R., S.J. Murch, S. KrishnaRaj and P.K. Saxena, 1999. Somatic embryogenesis and organogenesis in peanut: The role of thidiazuron and N6-benzylaminopurine in the induction of plant morphogenesis. Plant Growth Regul., 28: 9-15.
CrossRefDirect Link - Werner, T., V. Motyka, M. Strnad and T. Schmulling, 2001. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA., 98: 10487-10492.
CrossRefDirect Link