
S.H. Mohammad
Faculty of Education, Suez Canal University, Egypt
Research Journal of Environmental Sciences
Year: 2008 | Volume: 2 | Issue: 2 | Page No.: 100-107







ABSTRACT
Vertical zonation has a great effect on the intertidal snail Cerithium scabridum. This was clear from studying the abundance, population structure and spatial distribution of this snail from intertidal to subtidal zones in Lake Timsah, Suez Canal. The results showed that there was a significant difference in the abundance and prevailing size groups. Also, highly significant variations among depths and between sites were recorded for the growth of shell height, shell width and total weight.
PDF Abstract XML References Citation
How to cite this article
S.H. Mohammad, 2008. Vertical Zonation and Biometric Parameter of the Gastropod Cerithium scabridum in Suez Canal. Research Journal of Environmental Sciences, 2: 100-107.
DOI: 10.3923/rjes.2008.100.107
URL: https://scialert.net/abstract/?doi=rjes.2008.100.107
DOI: 10.3923/rjes.2008.100.107
URL: https://scialert.net/abstract/?doi=rjes.2008.100.107
INTRODUCTION
Genus Cerithium is represented in the Mediterranean and Red Sea by several species. Mediterranean (Cerithium repestre) and Red Sea (Cerithium scabridum) gastropods were studied in details by Rilov et al. (2004). Cerithium scabridum was reported from the Suez Canal by Keller (1882) and has established large stable populations from Egypt to South Eastern Turkey, Cyprus and Tunisia (Galil and Zenetos, 2002). It has been reported also from Italy (Vitturi et al., 2002). Distribution of other species such as Cerithium vulgatum in Atlantic coast of South Europe, Mediterranean Aegean, Marmara and Black sea was studied by Butakov et al. (1977). Grundel and Parent (2001) recorded other species (Cerithium diadeatum) in the uppermost Jurassic of Germany. Also, Cerithium bornii is found among the molluscan shells of Hong Kong (King and Ping, 1931) and Cerithium rubus was among four commonly abundant gastropod in Karachi (Rehman, 2002). Davidson et al. (2003) stated that Cerithium lutosum was among the dominant molluscs in Texas.
Cerithium sp. formed an essential food items in the food chains of many animals. Shell fragments of these snails were observed in the stomach of some crustaceans, gastropods, sea stars and fishes (Houbrick, 1974). Most fishes in the family libridae feed by crushing shelled prey in their powerful pharyngeal jaws (Wainwright, 1991). The snail Cerithium is a favored food item to the spiny lobsters Panulirus argus (Ndiaye and Bassène, 1996). It was also used as a recorder of change in water chemistry parameters primarily salinity. This expands the understanding of the effect of these parameters on the reproduction, growth and overall survival of the snails (Murray and Wingard, 2006). On the other hand, the small hermit crabs used mainly shells of Cerithium sp. as shelters. In addition, Cerithium sp. was used to camouflage sea Urchin tests. Medically, these snails served as intermediate hosts for numerous species of larval trematods (Sousa, 1992).
The aim of present study is to provide information on vertical distribution of Cerithium scabridum in Lake Timsah. Morphometric measurements and total weights were used to evaluate the changes in the gastropod growth and biometric aspects.
MATERIALS AND METHODS
The Cerithium samples were collected from Lake Timsah that lies between 30°33’ and 30° 35’ N latitude and 30°16’ and 32°19’ E longitude, Ismailia, Egypt. Six different sites were chosen along Lake Timsah (Fig. 1) in order to study this species.
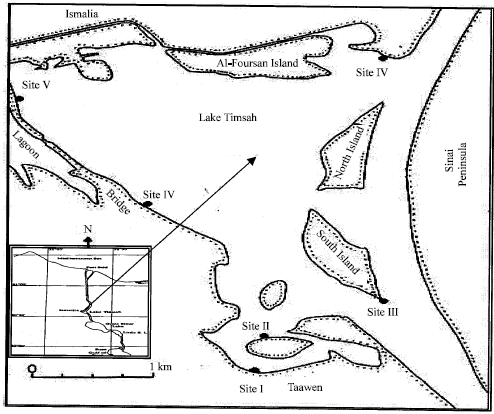 | |
| Fig. 1: | Map showing the investigated sites in Lake Timsah |
Cerithium scabridum was found only in the first three sites. At each site, four different depths were chosen in order to compare the population of this gastropod among them. Due to the narrow tidal range in the Suez Canal (<1 m), tidal depths were chosen at narrow range. These were: the beach (0-depth), 25, 50 and 80 cm depth. The samples were collected using a 20x20 cm quadrate in April 2006. These samples were washed with sea water through a sieve of 2 mm mesh size, counted and the average numbers were calculated for each site. For each specimen, height and width were measured to the nearest 0.1 mm by a Vernier Caliper. Also, total weights were calculated to the nearest 0.001 g. The data was analyzed by SPSS package. One-way ANOVA was performed to compare number of individuals and means of: shell height, shell width and total weight among depths and between sites. Some of water qualities such as salinity and pH value were measured. It did not differ greatly between sites. The salinity ranged from 43-43.5‰ and pH values were between 8.5 and 8.9 for the three sites.
RESULTS
Abundance
The present results revealed that number of individuals varied from one site to another. Site II constitute 89% of the total number whereas site I constitute only 8.16%. Site III had the lowest number (2.83%). By pooling the data for the three sites, Fig. (2) showed a bimodal structure with abundance of size groups 9-10 mm (16.2%) and 11-12 mm (14.7%).
Variation in the individual number was observed with depth. The highest number was recorded at 50 cm depth followed by 25 and 80 cm then the beach (site I). Site II revealed another pattern, the highest number was found at 80 cm followed by 50 cm and the beach, each of which contained the same number and finally at 25 cm depth.
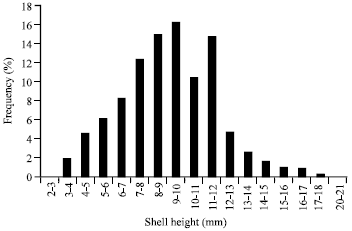 | |
| Fig. 2: | Size-frequency of Cerithium scabridum in the three sites |
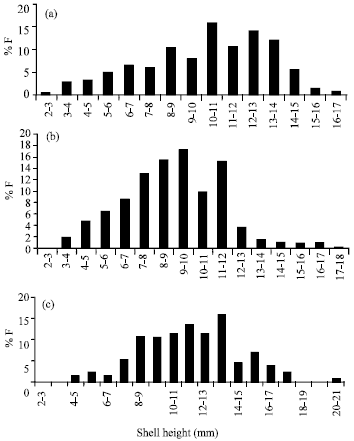 | |
| Fig. 3: | Size-frequency of Cerithium scabridum in site I (a), site II (b) and site III (c) |
A reverse pattern was detected in site III. The arrangement in a descending manner was as follow: the beach, 25 cm, 80 cm then 50 cm.
Population Structure
Figure (3a) revealed a gradual increase in the percentage number of individuals with increasing size. After size group 10-11 mm, the number of individuals suddenly declined then returned to increase at size group 12-13 mm. Thereafter, the number of specimens declined as the size increased. The other sites revealed the same pattern with little differences. Peaks of abundance were at the size groups 9-10 and 11-12 mm (site II) and at 11-12 and 13-14 mm (site III). Few individuals were observed at the smallest and largest size groups. Also, spat were recorded only at site I (Fig. 3a). Meanwhile, the largest snails were at site III (Fig. 3c).
Biometric Studies
Minimum shell height, shell width and total weight were recorded at 50 cm depth in sites I and III and at 25 and 80 cm depth in site II (Table 1). On the other hand, the maximum values for the three variables were restricted between 25 cm and the beach.
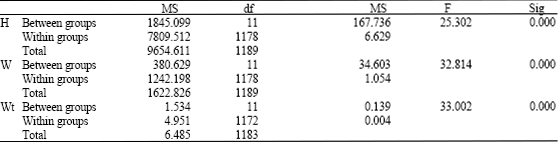
One-way ANOVA revealed highly significant differences between and within groups for each variable separately (Table 2).
| Table 1: | Average, maximum and minimum values of shell height (H), shell width (W) and total weight (Wt) of Cerithium scabridum at different depths |
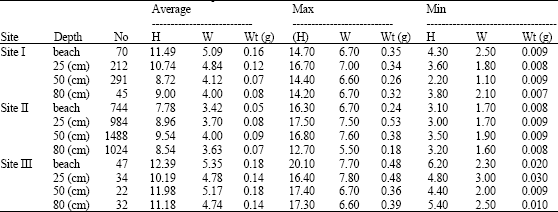 | |
| No: No. of specimens | |
| Table 2: | One-way ANOVA for three variables; shell height (H), shell width (W) and total weight (Wt) of Cerithium scabridum |
 | |
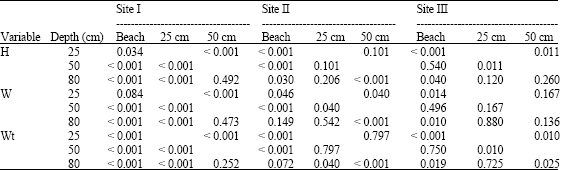
| Table 3: | Intraspecific variation for three variables; shell height (H), shell width (W) and total weight (Wt) of Cerithium scabridum |
 | |
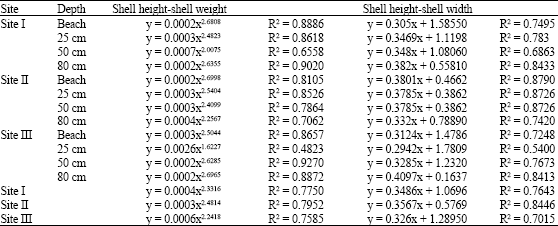
| Table 4: | Regression equations and correlation coefficient between shell height and each of shell weight and shell width of Cerithium scabridum |
 | |
| SiteDepthShell height-shell weightShell height-shell width | |
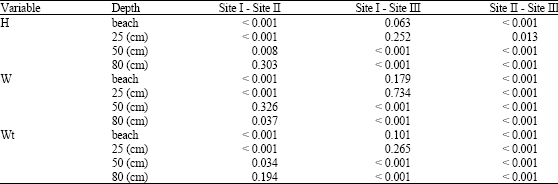
| Table 5: | Interspecific variation for three variables; shell height (H), shell width (W) and total weight (Wt) of Cerithium scabridum |
 | |
Intraspecific Relationship Site I:
This site revealed variation in the three variables (height, width, weight) among depths except those between 50 and 80 cm (Table 3).
Site II:
As site I, site II exhibited variation among depths for all variables. However, some depths showed non significant differences (among 25 and 50 cm and among 25 and 80 cm for height; among 80 cm and either of the beach and 25 cm for width and among the beach and 80 cm and among 25 and 50 cm for total weight).
Site III:
Both of shell height and shell width exhibited little variation among depths, whereas shell weight exhibited wide variation (Table 3).
The relationships between shell height and either of shell width and total weight were summarized in Table 4. It exhibited negative allometric growth with shell height.
Site-specific Relationships
Table 5 revealed that there were generally (significant-to highly significant) variations between sites at all depths. In spite of this, site I and site II exhibited similarity at beaches and at 25 cm depth. Also, sites I and III did not vary significantly at 50 cm (for the width variable) and at 80 cm (for height and weight variables). Whereas sites II and III were completely differed from each other at all depths.
DISCUSSION
Cerithium scabridum, in the present study, occurred from the intertidal to the sublittoral zone in Lake Timsah. Migotto et al. (1993) also observed this gastropod in intertidal and sublittoral zones in Brazil. Cerithium scabridum showed variations in the numbers, morphometric measurements (shell height and width) and total weights in different tidal level. Few individuals were recorded in the deepest zone of sites I and III. In addition, the lowest values of shell measurements and total weights were restricted between 50 and 80 cm depth. So, it could be concluded that Cerithium scabridum preferred more exposed area (intertidal) than that which was continuously immersed with water (sublittoral). This was confirmed by Fujioka et al. (2007) who stated that Cerithium coralium decreased evidently as the depth increased. Also, Arruda (2000) found most records of this genus from the lower intertidal region in spite of their occurrence from the intertidal to the sublittoral zone in Brazil. On the other hand, the highest values of morphometric measurements as well as total weights of these snails, of the present study, were restricted between the beach (zero-level) and 25 cm depth. This, together with the detection of spat in the deep zone only, might led to the hypothesis that snails moved to the deep zones for spawning, thereafter they returned back to their habitat. This was ensured by Denadai et al. (2004) who stated that this genus bred mainly in spring (the time of collection in the present study). Then spat gradually migrated to less depth zones perhaps for the availability of food. This assumption was cited by Cannon (1979) who found snails of Cerithium migrated to higher exposed levels as they grew. Such migratory behavior is common in marine gastropods, which tend to translocate from settlement area to the adult habitat (Noda and Nakao, 1995). On the base of the previous conclusions, the present results might be elucidated. Time of spawning might vary between sites and among depths. The number of individuals was quite few in the deepest region but the highest was as 50 cm depth. Then the number gradually decline as they go the beach (quarter and half of the previous number in site I and II, respectively). It can be assumed that few individuals spawned and moved toward the beach while the majority was in the spawning condition. However, site III differed from the other sites. Most individuals were recorded at the beach and the number declined with increase depth. This may mean that most snails in this site might spawn in the former period, migrated already to the beach where they could feed and grow. The highest number of the largest size groups comparing with the other sites supports the previous hypothesis. This vertical movement was not only associated with reproduction but also with other factors such as habitat. Members of genus Cerithium are generally epifaunal and live in association with vegetation such as algae and sea grass (Houbrick, 1992). Denadai et al. (2004) concluded that the greater abundance of sea grasses and seaweeds indicated that this was the preferred habitat for adults Cerithium scabridum, where they can have greater reproductive success. Analysis of the results of Davidson et al. (2003) demonstrated that sea grass biomass influenced the abundance of the herbivore and epifaunal groups of Cerithium lutosure. This might illustrate the largest size to which Cerithium scabridum attained at site III in the present study. So, this site was more suitable for their growth comparing with the other sites. This may be due to the continuous water movement, resulted from ships motion at this site, which may refresh feed source of the snails. A similar result was detected by Cannon (1979) who found little seasonally related change in the abundance of Cerithium moniliferum, but number and shell morphs were correlated with sand movements. Not only availability of food affect on Cerithium sp. abundance but also the microhabitat. Chapman (1994) found correlation between snails' densities and sizes with the distribution of particular microhabitat such as the slope of the surface. So, more studies are needed on the nature of these sites in Lake Timsah.
ACKNOWLEDGMENTS
The author would like to thank Prof. Dr. S. Zakaria for valuable comments on the manuscript.
REFERENCES
- Denadai, M.R., A.C.Z. Amaral and A. Turra, 2004. Biology of a tropical intertidal population of Cerithium atratum (Born, 1778) (Mollusca, Gastropoda). J. Nat. History, 38: 1695-1710.
Direct Link - Fujioka, Y., T. Shimoda and C. Srithong, 2007. Diversity and community structure of macrobenthic fauna in shrimp aquaculture ponds of the gulf of Thailand. JARQ, 41: 163-172.
Direct Link - Grundel, J. and H. Parent, 2001. Lower and Middle Tithonian marine gastropods from the Neuquén-Mendoza basin, Argentina. Boletin del Instituto de Fisiografiay Geologia, 71: 13-18.
Direct Link - Rilov, G., Y. Benayahu and A. Gasith, 2004. Prolonged lag in population outbreak of an invasive mussel: A shifting-habitat model. Biol. Invasions, 6: 347-364.
CrossRef - Vitturi, R., M. Colomba, L. Castriota, A.M. Beltrano, A. Lannino and N. Volpe, 2002. Chromosome analysis using different staining techniques and fluorescent in situ hybridization in Cerithium vulgatum (Gastropoda: Cerithiidae). Hereditas, 137: 101-106.
Direct Link