
Mercy G. Ajuru
Department of Biology, Ignatius Ajuru University of Education, Port Harcourt, Rivers, Nigeria
Bosa E. Okoli
Department of Plant Science and Biotechnology, University of Port Harcourt, P.M.B. 5323, Port Harcourt, Nigeria
Research Journal of Botany
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 15-23







ABSTRACT
The detailed anatomy of four species and three varieties of melons in an interesting and somewhat unusual family, Cucurbitaceae is presented. Parts of each species (root, stem and leaf) were sectioned with a rotary microtome at 20 to 24 μm thickness after fixation and wax embedding. Sections were observed using a photomicroscope. There were similarities in the arrangement and distribution of cells and tissues in the organs under investigation. Nonetheless, the number of layers of these cells and tissues (collenchyma, parenchyma, pericycle, sclerenchyma and vascular bundles) differed which is taxonomically important. Comparative anatomical studies of the melons in this family is presented together for the first time with variations in the number of tissues in the different varieties of Citrullus lanatus-watermelon found in Nigeria.
PDF Abstract XML References Citation
Received: August 16, 2012;
Accepted: December 11, 2012;
Published: May 24, 2013
How to cite this article
Mercy G. Ajuru and Bosa E. Okoli, 2013. Comparative Vegetative Anatomy of Some Species of the Family Cucurbitaceae Juss in Nigeria. Research Journal of Botany, 8: 15-23.
URL: https://scialert.net/abstract/?doi=rjb.2013.15.23
URL: https://scialert.net/abstract/?doi=rjb.2013.15.23
INTRODUCTION
The Cucurbitaceae family consists of herbaceous climbing and trailing plants, found abundantly in the warmer regions of the world (Berrie et al., 1987; Rubatzky and Yamaguchi, 1997). There are about 120 genera and more than 800 species, but in Nigeria, it is represented by 21 genera and 41 species (Hutchinson and Dalziel, 1954).
The species are annual or perennial herbs, characterized by a very extensive root system which ramifies in the surface soil. They have hollow angled stems which may be glabrous, hairy or prickly and their vascular bundles are bicollateral. The leaves are large, simple, but often deeply lobed, alternate or spirally arranged on long petioles. In each leaf axils, there is a flower bud, a vegetative bud and a tendril. The plants are monoecious or dioecious with unisexual flowers occurring singly in the leaf axils. Monoecious plants are protandrous with male flowers produced before the females and in much greater numbers. Fruits are usually large and fleshy, berry or pepo, or a fleshy or dry capsule. Seeds are without endosperm. Cucurbitaceae is of high economic value, being a major source of food for man. Their fruits are eaten mainly as vegetables, though some species produce fruits which are eaten as a dessert. Others such as Lagenaria siceraria (calabash, bottle gourd) and Luffa cylindrica (loofah) are cultivated for making items of utility such as drinking vessels, cooking pots, utensils, bath sponge, in industrial filters and in sound insulation (Heiser Jr., 1979). Some species are cultivated as garden ornamentals. Members of the family contain cucurbitacins, which are sometimes bitter (Okoli, 1984).
According to Stace (1980), the anatomical structure of plants are useful in the taxonomic characterization of plants since they are less affected by environmental changes and therefore, highly conservative in taxa variations. The distribution and arrangement of plant cells and tissues such as sclerenchyma, vascular bundles and other anatomical features have been reported and utilized at different systematic levels for delimitation of taxa (Metcalfe and Chalk, 1979; Stace, 1980; Fahn, 1990; Ndukwu and Okoli, 1992; Agbagwa and Ndukwu, 2004; Ajuru, 2012). Comparative and systematic studies on the anatomy of the root, stem and leaf of all the melon species belonging to the Cucurbitaceae family in Nigeria is being presented together for the first time. The three species, Cucumis melo, Cucumeropsis mannii, Cucurbita moschata and the different varieties of the species, C. lanatus studied are economically important. This study forms part of a research carried out on various aspects of the biosystematics of these species. The aim of this study is to carry out in depth anatomical studies of the root, stem and leaf of the different species of the family Cucurbitaceae with a view to providing features that may be of taxonomic value in their identification and classification.
MATERIALS AND METHODS
Fresh plant materials collected during field trips to various parts of Nigeria and germinated in plastic bags in the botanical garden of the University of Port Harcourt were used (Table 1). The plants were transplanted directly into the soil in the botanical garden. They are: Cucurbita moschata (Duch. Ex Lam.) Duch. Ex Poiret (Squash melon), Cucumeropsis mannii Naud. (syn. C. edulis (Hooker f.) Cogn.) (White-seeded melon), Cucumis melo L. (Sweet melon), Citrullus lanatus Thunb. Matsum and Nakai (Brown-seeded melon) and Citrullus lanatus Thunb. Matsum and Nakai (Watermelon of three varieties-Light green exocarp color (A), Deep green exocarp color (B) and Light green exocarp with deep green vertical lines (C). Fresh root, stem and leaf of all the species studied were fixed in Formal-Acetic-Alcohol (FAA) for 48 h, washed in several changes of distilled water, dehydrated through ethanol-water mixtures (30, 50, 70, 95 and 100%), 2 h in each solution, cleared by transferring to 25, 50 and 75% alcohol-chloroform mixtures for two hours in each, followed by a mixture of pure chloroform and wax flakes and embedded in wax. Sections were cut on a Leitz 1512 rotary microtome at thickness between 20 and 22 μm. The sections were dewaxed with pure xylene and rehydrated by first of all rinsing in a bath of absolute ethanol, drained and passed through alcohol series to water. Sections were stained by dipping the slides in 1% safranin for 5 min and washed with distilled water. They were then mounted permanently in DPX (Dee Pex) according to the method employed in the, “Anatomical studies in the leaf and probract of Telfairia Hooker” (Okoli, 1987).
| Table 1: | Sources of plant materials used for the study |
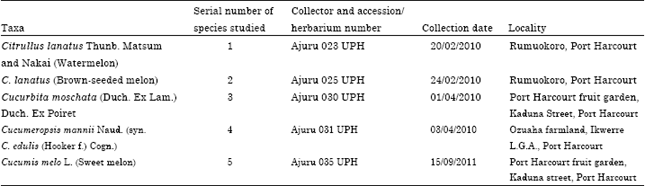 | |
Photomicrographs of good sections were taken with a Leitz Diaplan photomicroscope fitted with Leica WILD MPS 52 camera at x10 microscope objective lens. Microscope measurements of the sizes of cells and tissues in root, stem and leaf were made at x10 microscope objective lens following Radford et al. (1974).
RESULTS
Root: There were observable differences in the number of layers of tissues (Fig. 1a-g). In C. moschata, the root anatomy (Fig. 1a) starts with one layer of epidermal cells followed by 3 cell layers of irregularly shaped epiblema cells, 5-6 layers of cortical cells and l layer of thick walled endodermal cells.
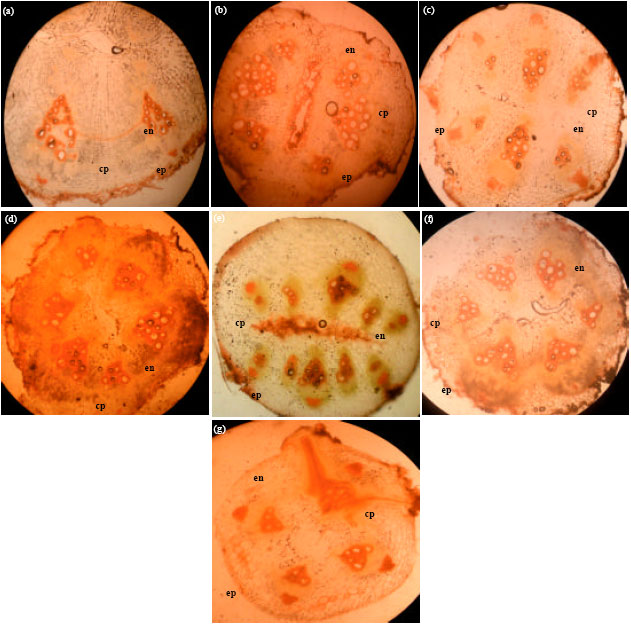 | |
| Fig. 1(a-g): | Cross-sections of the root anatomy of species of Cucurbitaceae (x100), (a) C. moschata, (b) C. lanatus-brown seeded melon, (c) C. mannii, (d) C. lanatus-light green watermelon, (e) Deep green watermelon, (f) Light green with deep green vertical stripe watermelon and (g) C. melo, cp: Cortical parenchyma, ep: Epidermis, en: Endodermis |
Several layers of sclerified phloem fibers are buried in the tissues with some obliterating the endodermis. Xylem occurs towards the centre. The pith is hollow but is surrounded by sclerified parenchyma cells. In C. lanatus-brown-seeded melon, the root anatomy (Fig. 1b) is made up of an outer layer of epidermal cells followed by 2 layers of compactly arranged and rectangular epiblema cells, 4-5 layers of cortical parenchyma cells, 1 layer of endodermal cells. The phloem fibres occasionally obliterates the endodermis. In C. mannii, the root anatomy (Fig. 1c) consists of an outer epidermal layer followed by 2-3 layers of isodiametric and rectangular epiblema cells, 4 layers of thin walled, cylindrical cortical parenchyma cells and 1-2 layers of endodermal cells. The pith is characterized by sclerified parenchyma cells. In C. lanatus-light green, deep green and light green with deep vertical stripe watermelon varieties, the root anatomy (Fig. 1d-f) begins with a layer of epidermal cells followed by a layer of thick walled epiblema cells, 6 layers of thin walled cortical cells in light green watermelon, 7-8 layers in deep green watermelon and 8-9 layers in light green with deep green vertical stripe watermelon. The endodermal layer is one celled thick. The pith is hollow and surrounded by sclerified parenchyma cells in light green and light green with deep vertical stripe watermelon but filled with sclerified parenchyma cells in the deep green watermelon. In C. melo, the root anatomy (Fig. 1g) starts with an epidermal layer that is one celled thick, followed by 1-2 layers of epiblema cells, 5-6 layers of thin walled, loosely packed cortical cells, 1-2 layers of endodermis which is obliterated by a vascular tissue which extends to the cortical cells. The pith is surrounded by sclerified parenchyma cells.
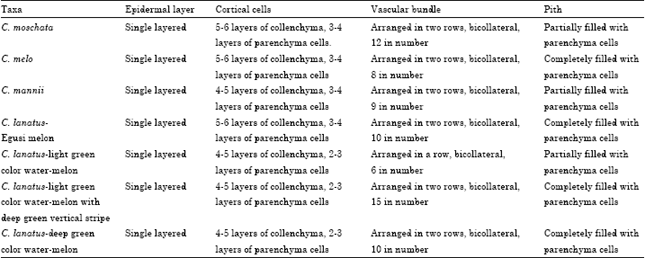
Stem: Generally, the stem of the cucurbit species studied are hollow with five ridges and five furrows (Fig. 2a-g, Table 2). The vascular bundles are arranged in two rows, those of the outer row corresponds to the ridges and those of the inner to the furrows.
The epidermis in all the species is single layered passing over the ridges and furrows. The hypodermis or collenchyma lies immediately below the epidermis and consists of 5-6 layers of collenchyma cells in C. moschata, C. lanatus-egusi melon and C. melo and 4-5 layers in C. mannii and C. lanatus-watermelons. The cells contain chloroplasts. The cortical parenchyma which lies below the collenchyma, forms a narrow zone in the middle, 3-4 layers thick in C. moschata, C. lanatus-egusi melon, C. melo and C. mannii; and 2-3 in C. lanatus-watermelons.
| Table 2: | Summary of stem anatomical characters in the Cucurbitaceae species studied |
 | |
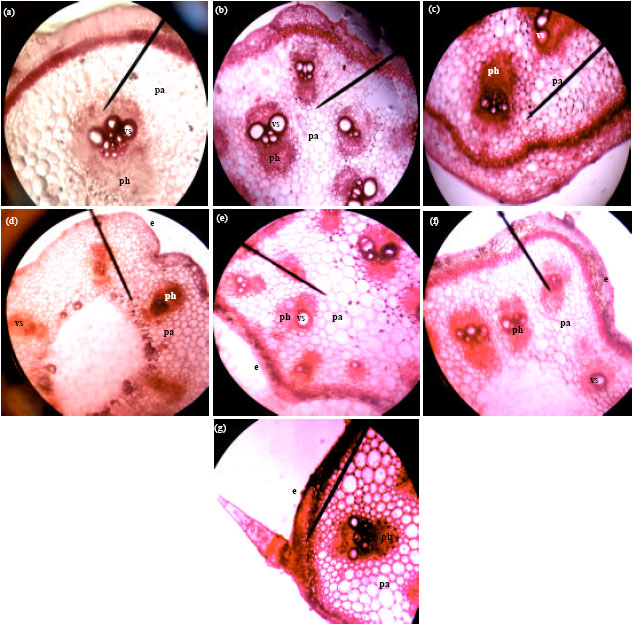 | |
| Fig. 2(a-g): | Cross-sections of the stem anatomy of Cucurbitaceae species (x400) (a) C. moschata, (b) C. lanatus-brown-seeded melon, (c) C. mannii, (d) C. lanatus-light green watermelon, (e) C. lanatus-deep green watermelon, (f) C. lanatus-light green with deep green vertical stripe watermelon and (g) C. melo, e: Epidermis, pa: Parenchyma, vs: Vessels, ph: Phloem |
Chloroplasts are abundant in it. The endodermis, which is the innermost layer of the cortex, lies immediately outside the pericyclic sclerenchyma. It is single layered in all the species and contains starch grains. Directly below the endodermis is a zone of sclerenchyma, which represents the pericycle. It is 4-5 layers thick in all the species and is made up of thick-walled, lignified cells, which are polygonal. There is a continuous mass of thin-walled, parenchymatous cells extending from below the sclerenchyma to the pith cavity which is completely filled up in C. melo, C. lanatus-egusi melon, C. lanatus-deep green exocarp color watermelon and light green with deep green vertical stripe watermelon, but partially filled up with a central hollow in C. mannii, C. moschata and C. lanatus-light green exocarp color watermelon. In all the species, the vascular bundles are bicollateral and arranged in two rows. Each bundle consists of xylem, two strips of cambium and two patches of phloem. The vascular bundles are twelve in C. moschata, ten in C. lanatus-egusi melon and C. lanatus-deep green exocarp color watermelon, nine in C. mannii, eight in C. melo and fifteen in C. lanatus-light green with deep green vertical stripe watermelon. In C. lanatus-light green exocarp watermelon, the vascular bundles are six in number, arranged in a single row. The xylem tissue occupies the centre of the bundle. It has protoxylem, which are smaller vessels on the inner side and metaxylem, which are slightly bigger vessels a little higher up. The cambium tissue occurs in two strips, one on each side of the xylem. It forms a narrow strip between the phloem and the xylem to the inside and between the xylem and the phloem to the outside. The phloem tissue occurs in two patches. The outer phloem is plano convex and the inner one is semi lunar. Each patch of phloem consists of sieve-tubes, companion cells and phloem parenchyma.
Leaf: The upper epidermis of the leaves of all the species is a single layer of cells with a thick cuticle which checks excessive evaporation of water from the surface (Fig. 3a-g, Table 3). The lower epidermis is also a single layer of cells but with a thin cuticle. There are two to three layers of palisade parenchyma in all the species which are elongated, cylindrical cells, closely packed together, with narrow intercellular spaces here and there. They contain numerous chloroplasts. The spongy parenchyma cells are irregular in shape, loosely arranged towards the lower epidermis, with numerous intercellular air spaces between them. There are about eight vascular bundles in C. melo, six in C. lanatus-deep green exocarp color watermelon, four in C. moschata and C. lanatus-egusi melon, three in C. lanatus-light green with deep green vertical stripe watermelon and C. lanatus-light green exocarp color watermelon and two in C. mannii. The vascular bundles are bicollateral. Each consists of xylem, which lies towards the upper epidermis and phloem, which lies towards the lower epidermis. The xylem consists of various kinds of vessels, tracheids and xylem parenchyma. The phloem consists of some narrow sieve-tubes, companion cells and phloem parenchyma. Surrounding each vascular bundle is the border parenchyma or bundle sheath, which is a compact layer of thin-walled parenchymatous cells. There are one to two patches of sclerenchyma associated with the vascular bundles.
| Table 3: | Summary of leaf anatomical characters in the Cucurbitaceae species studied |
 | |
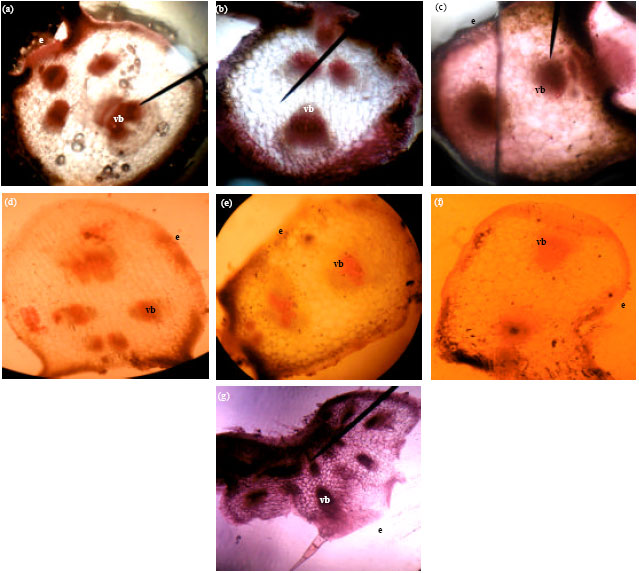 | |
| Fig. 3(a-g): | Cross-sections of the leaf anatomy of Cucurbitaceae species (x10) (a) C. moschata, (b) C. lanatus-brown-seeded melon, (c) C. mannii, (d) C. lanatus-light green watermelon, (e) C. lanatus-deep green watermelon, (f) C. lanatus-light green with deep green vertical stripe watermelon and (g) C. melo, e: Epidermis, vb: Vascular bundle |
DISCUSSION
The variations that exist among the species of Cucurbit studied is of taxonomic value as reported by Stace (1980), AbdulRahaman and Oladele (2010a, b), Ogunkunle and Oladele (2008) and Ndukwu and Okoli (1992). The anatomical structures of all the species as observed in this study is in agreement with the findings of Esau (1965) on herbaceous plants. The primary plant body is made up of the dermal system, the epidermis, on which are glandular multicellular trichomes, the fascicular tissue system composed of the xylem and phloem and the fundamental or ground system represented by the cortex and pith. C. moschata, C. mannii and C. lanatus-light green exocarp color watermelon exhibit amphiphilic type of siphonostele while C. lanatus-egusi melon, deep green exocarp color watermelon, light green exocarp with deep green vertical stripe color watermelon and C. melo exhibit ectophloic type of siphonostele, with bicollateral vascular bundles of various number in all the species. The pith lies to the inside of the stele and as in other herbaceous stems, is wide in diameter and continues to occupy a high proportion of the stem throughout life. This is also seen in Helianthus tuberosus L. and H. annuus L. (Metcalfe and Chalk, 1979).
All the species in this study are said to be drought resistant (Oomen and Grubben, 1977). The presence of the cuticle, sclerenchymatous sheath in the leaves and increased number of glandular hairs in the stems and leaves support this view. The medullary rays alternate with the bundles linking up with the cortex instead of forming a continuous cylinder. This then explains why the plants are creeping/trailing and also a climber in the case of C. mannii. The diagnostic features of all the species therefore as belonging to the family Cucurbitaceae include the presence of bicollateral vascular bundles, arrangement of the vascular bundles in two rows to coincide with the five furrows and ridges and presence of siphonostele.
The observed anatomical similarities among the cucurbit species studied indicate phylogenetic relatedness of the taxa. These anatomical differences observed in each species must have been as a result of evolution, conferring heritable variation that could be exploited for taxonomic purposes. The present study is an attempt to provide anatomical data on the species of Cucurbit earlier described by Hutchinson and Dalziel (1954).
In conclusion, based on anatomical features, the species of melons studied can readily be distinguished from one another as a result of variation in the number of layers of tissues and vascular bundles present in them. This information can be useful in herbal medicine and Forensic Science where, identification and authentication of plant specimens are essential.
ACKNOWLEDGMENT
We are grateful to the Curators at the Obafemi Awolowo University of Ife, Ille-Ife, Osun State, for their assistance during the study.
REFERENCES
- Abdulrahaman, A.A. and F.A. Oladele, 2010. Stomatal complex types and epidermal cells in Jatropha species L. (Euphorbiaceae). Nig. J. Pure Applied Sci., 23: 2160-2163.
Direct Link - Agbagwa, I.O. and B.C. Ndukwu, 2004. The value of morpho-anatomical features in the systematics of Cucurbita L. (Cucurbitaceae) species in Nigeria. Afr. J. Biotechnol., 3: 541-546.
Direct Link - Ogunkunle, A.T.J. and F.A. Oladele, 2008. Leaf epidermal studies in some Nigerian species of Ficus L. (Moraceae). Plant Syst. Evolution, 274: 209-221.
CrossRef - Okoli, B.E., 1984. Wild and cultivated cucurbits in Nigeria. J. Econ. Bot., 38: 350-357.
CrossRefDirect Link