
M. S. Dar
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
Irtefa Mohammed
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
T. A. Sofi
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
F. A. Ahanger
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
M. D. Shah
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
Mushtaq Ahmad
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
Aflaq Hamid
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
A. A. Mir
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
Asha Nabi
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
B. A. Padder
Plant Virology and Molecular Plant Pathology Laboratory, Division of Plant Pathology, SKUAST, 190 025, Shalimar Srinagar, Kashmir
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 196-201







ABSTRACT
Apple scab caused by a fungus Venturia inaequalis cause enormous losses to growers both qualitatively and quantitatively. In order to manage the crop, farmers routinely spray 8-12 fungicides from pink bud till harvest. Hence, the best strategy to manage the disease is planting apple scab resistant cultivars. In the present study, four races viz., (0), (1), (2) and (1, 2) were reported from various commercial apple growing regions of Kashmir. Apple scab races were spatially distributed in all apple growing regions of Kashmir. Race 1 was most prevalent among the four races. Screening of 31 apple genotypes under controlled conditions revealed majority of commercial cultivars susceptible. Genotypes carrying RVi3 to RVi13 scab resistance genes along with the cultivars American Apirouge, H27, Shireen and Firdous (latter two contain RVi6 scab resistance gene) were found resistant to all the four races present in Kashmir. Resistant genotypes reported in the present study can easily be exploited by breeders for management of apple scab resistance using marker assisted selection approach.
PDF Abstract XML References Citation
Received: June 25, 2015;
Accepted: August 03, 2015;
Published: September 29, 2015
How to cite this article
M. S. Dar, Irtefa Mohammed, T. A. Sofi, F. A. Ahanger, M. D. Shah, Mushtaq Ahmad, Aflaq Hamid, A. A. Mir, Asha Nabi and B. A. Padder, 2015. Distribution of Apple Scab Race Flora and Identification of Resistant Sources against Venturia inaequalis in Kashmir. Plant Pathology Journal, 14: 196-201.
DOI: 10.3923/ppj.2015.196.201
URL: https://scialert.net/abstract/?doi=ppj.2015.196.201
DOI: 10.3923/ppj.2015.196.201
URL: https://scialert.net/abstract/?doi=ppj.2015.196.201
INTRODUCTION
Apple scab caused by Venturia inaequalis is a catastrophic disease world over causing enormous loss to growers. It is the most important disease of apple in Kashmir, a North Western Himalayan state of India. Like other apple growing regions worldwide, scab is currently being managed by fungicidal sprays from pink bud to harvest in the State (Padder et al., 2013). Public concerns over pesticide residue in food have generated a great deal of interest in reducing fungicide use on all food crops. Several strategies like disease predictors or forecasters (Fisher and Lillevik, 1977; Ellis et al., 1984; Jones et al., 1984) combined with ergosterol biosynthesis inhibiting fungicides have been employed in scab management with varying degree of success (Sutton, 1996). The latter strategy readily promotes selection of fungicidal resistant strains leading to failure of disease management in various apple growing regions. In spite of the fact, control of apple scab in most of countries heavily depends on foliar application of fungicides. Thus, the best strategy to manage disease is planting resistant cultivars, which is most effective, least expensive and easiest for farmers to adopt. Many scab resistant cultivars have been bred in various countries (Crosby et al., 1992; MacHardy et al., 2001) majority of which contain Rvi6 (previously known as Vf) resistance gene. Development of these varieties had no market impact and hence did not resulted in displacement of major susceptible cultivars (MacHardy, 1996). Therefore, efforts to produce durable scab resistant cultivars with market acceptability should be given priority in breeding programmes. In order to devise such programmes with success, apple genotypes have to be screened for scab resistance under in vitro condition with fungus races present in particular region. Although various commercial apple cultivars have been reported susceptible to scab under natural infection (Didelot et al., 2007; Le Van et al., 2011) however, work on elucidating resistance in the greenhouse conditions against individual races is diminutive. Hence, there is a need to search for scab resistant cultivars against different races for fungus.
In India, particularly Kashmir valley, there is little information on the susceptibility of apple cultivars to scab races, including the cultivars such as Lal Ambri, Gulshan, Shreen, Firdous, Akbar, Shalimar 1 and Shalimar 2 which have been bred in the state. Deployment of these cultivars spatially and temporally depends on understanding of the population biology, virulence spectrum, genomics and evolutionary process of V. inaequalis in a region. Elucidation of virulence structure showed four distinct races viz., (0), (1), (2) and (1, 2) in Kashmir. Population structure based on RAPD, ISSR and CAPS revealed low genetic differentiation in the different scab pathogen population’s thereby indicating occurrence of frequent gene flow in the region (Padder et al., 2011, 2013). This information is a vital component for deployment of resistant cultivars spatially, in organic farming and of any integrated pest and disease management program. Study under investigation was designed with the following objectives to (1) Assess the pathogenic variability in V. inaequalis with an aim to identify any shift in virulence pattern from the already reported races in the region (Padder et al., 2013), (2) Distribute the apple scab race flora (from the present investigation and previously reported) spatially in commercial apple growing regions of Kashmir valley and (3) Evaluate apple genotypes against each individual race under controlled conditions with an intention to identify scab resistant cultivars and their distribution/deployment in time and space.
MATERIALS AND METHODS
Collection of new scab samples and distribution of race flora: Seventy one isolates characterized by Padder et al. (2013) revealed the existence of 4 races viz., (0), (1), (2) and (1, 2) within the 71 of isolates. These isolates were collected during the year 2009 and 2010. In order to know whether there has been any pathogen virulence shift, commercial apple growing areas (45 locations) were surveyed during 2014 in 10 districts viz., Anantnag, Bandipora, Badgam, Pulwama, Shopian, Kulgam, Kupwara, Ganderbal, Srinagar and Baramulla of Kashmir valley with majority of V. inaequalis isolates from Red Delicious cultivar. All the isolates were collected from the same locations surveyed previously. Mono-conidial isolations were done using single spore technique and each isolate was inoculated on an international differential set comprising 14 apple accessions (Patocchi et al., 2009). The inocula, produced as per Barbara et al. (2008), were each adjusted to 5×105 conidia per milliliter and the conidial suspensions applied till runoff with a manual atomizer. Three one-year-old potted trees were inoculated for each isolate and incubated in a Hi-Tech glasshouse (Rajdeep Agri Products, New Delhi, India) set at 20°C with 85% relative humidity. To ensure constant leaf wetness, the potted trees were placed in polythene covered structure and incubated for 48 h in dark at a constant temperature of 20°C; leaf wetness was maintained using a humidifier. Symptoms were assessed 14 days after inoculation. The potted trees were classified as per 0-4 scale of Chevalier et al. (1991), wherein trees in classes 0, 1, 2 and 3a were graded as resistant and trees showing class 3b and 4 were graded as susceptible.
In order to know the prevalence of particular race(s) spatially in different commercial apple growing areas, previous races characterized earlier by authors and the new races characterized in the present study (a total of 116 isolates) were distributed to various districts according to their location.
Resistance evaluation under glass house conditions: For resistance evaluation under controlled conditions, 31 genotypes of apple were inoculated with 4 races (single isolate). Inoculum from each isolate was produced as described by Barbara et al. (2008). The inoculum of each race was adjusted to 5×105 conidia per milliliter and the conidial suspensions applied till runoff with a manual atomizer. One-year-old potted trees were inoculated (three sets for each race), in a Hi-Tech glasshouse (Rajdeep Agri Products, New Delhi, India), set at 20°C with 85% relative humidity. To ensure constant leaf wetness, the potted trees were placed in polythene covered structure and incubated for 48 h in dark at a constant temperature of 20°C; leaf wetness was maintained using humidifier. Symptoms on potted trees were assessed 14 days after inoculation. The potted trees were classified according to the grading system of (Chevalier et al., 1991) with 0-4 class scale: Trees with class 0, 1, 2 and 3a were graded as resistant and trees showing class 3b and 4 were graded as susceptible.
RESULTS AND DISCUSSION
Among the 45 V. inaequalis isolates inoculated on apple scab differentials, 32 (71.11%) were categorized into race (1). These isolates were able to breach the resistance gene Rvi1 present in Golden Delicious in addition to Royal Gala. Seven isolates were able to overcome two scab resistance genes Rvi1 and Rvi2 present in Golden Delicious and TSR34T15, respectively and were designated as a complex race (1, 2). Three isolates each were grouped into race (0) and race (2) (Table 1). Emphases is being given to monitor the virulence alleles throughout the world under the umbrella of “Monitoring of Venturia inaequalis virulences” by Patocchi et al. (2009). Many researchers across the globe are monitoring the new virulence’s in apple scab fungus and data is being validated and maintained (www.vinquest.ch).
| Table 1: | Discerning the 45 isolates of Venturia inaequalis into different races based on reaction on International differential set proposed by Patocchi et al. (2009) |
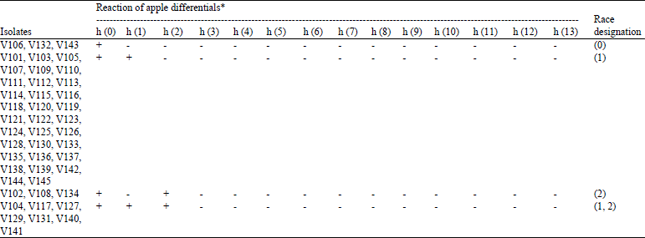 | |
| H0: Royal gala, h (1): Golden Delicious, h (2): TSR34T15, h (3): Geneva, h (4): TSR33T239, h (5): 9-AR2T196, h (6): Priscilla, h (7): M. x floribunda 821, h (8): B45, h (9): K2, h (10): A723-6, h (11): M. baccata jackii, h (12): Hansen’s baccata #2, h (13): Durello di Forlì , +: Susceptible, -: Resistant | |
Thus, our main objective was to identify any shift in the virulence spectrum. This shift in pathogenicity was not exhibited by the isolates collected during 2014. These isolates were collected after three years of our previous study (Padder et al., 2013) which may be attributed to the less time lapse in survey or the absence of scab resistance genes (Rvi3 to Rvi13) in the orchards of Kashmir valley. Additionally, V. inaequalis is described as a good model for host adaptation in plant pathogens (Giraud et al., 2010) because mating only occurs between strains that are able to infect identical hosts, thus facilitating the maintenance of adaptations to a host when host ranges do not overlap (Guerin and Cam, 2004; Guerin et al., 2007; Gladieux et al., 2011). Moreover, population genetic studies have shown that apple resistance genes might induce specialization in V. inaequalis populations, thus favouring host-related adaptations (Guerin and Cam, 2004; Gladieux et al., 2011). Since, majority of area under apple belts in Kashmir is under Red Delicious, Golden Delicious, Star Crimson and Royal Delicious cultivars, thus fungus has adopted and evolved to overcome the resistance specificities present in them. The race (1) was most wide spread (71.11% isolates belonged to this race) and the results are in conformity with our previous study and with many researchers especially from the Europe and the USA (Heaton et al., 1991; Parisi et al., 2004; Bus et al., 2011).
The distribution pattern of races in different apple growing areas of Kashmir valley depicted in disease map (Fig. 1) revealed the presence of one or the other race of scab pathogen, V. inaequalis. Race (1) was most widespread and prevalent in most of the apple growing of area districts. Various locations in district Srinagar and Budgam contained all the four races whereas three out of four races were present in locations of Kupwara district. In addition to race (1), race (0) was also observed in areas of Shopian and Bandipora whereas, locations in district Baramulla, Kulgam and Anantnag only race (1) isolates were present. In India, Gupta (1990) speculated the presence of only 2 races of apple scab pathogen in various apple growing regions of Kashmir and Himachal Pradesh however, Padder et al. (2013) first time reported presence of 4 different races of apple scab from Kashmir valley and were reconfirmed in present study. So, presence of these races in Himachal Pradesh cannot be unnoticed and the occurrence of many more (both simple and complex races) cannot be ruled out from North Western Himalayan region because of its close proximity to the centre of origin of apple. Since, discerning pathogenic variability in Venturia inaequalis populations forms the back bone of any breeding strategy, timely monitoring of different virulence’s should be given priority in India like the European DARE programme (Lespinasse, 1989; Patocchi et al., 2009).
Screening of 31 apple genotypes under glass house conditions with 4 races of scab pathogen prevalent in the Kashmir valley revealed majority of commercially grown genotypes susceptible to one or the other race of pathogen (Table 2). Genotypes, which contain well characterized scab resistance genes were resistant to all the 4 races. Among the commercially grown apple cultivars American Apirouge, H27, Shireen and Firdous exhibited resistance to all the races, whereas Saharanpuri showed resistance towards 3 races viz., race (0), (1), (1, 2). Two cultivars Star Crimsion and Sunhari were resistant to race (0) whereas, White Dotted Red and Golden Delicious were resistant to two race (0), (1) and race (0, 2), respectively. Commercially grown apple cultivars viz., Ambri, Cox Orange Pippen, Gala, Gulshan, Lal Ambri, Red Delicious, Red Gold and Spartan were susceptible to all the 4 races of pathogen (Table 2). Most of the differentials and few cultivars exhibited pin point hypersensitivity reaction (type 1 symptoms) and chlorosis (type 2) on inoculation with 4 races.
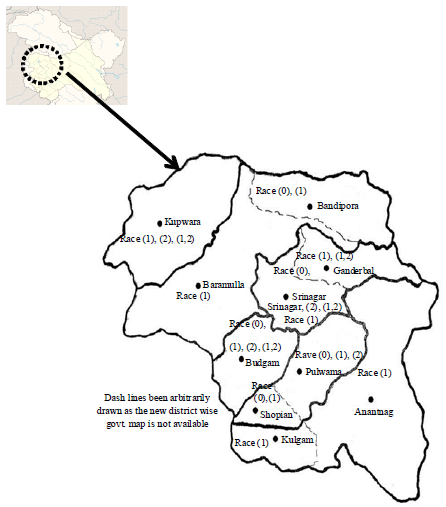 | |
| Fig. 1: | Distribution of Venturia inaequalis race flora in different districts of Kashmir |
| Table 2: | Resistant and susceptible apple genotypes to 4 races of Venturia inaequalis in Kashmir |
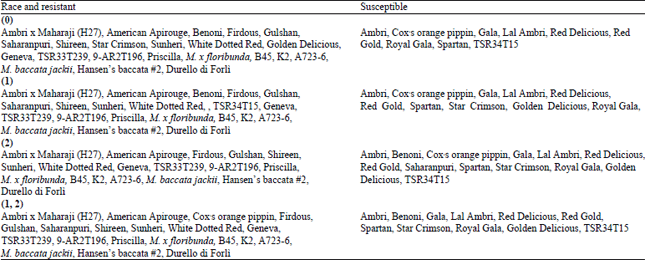 | |
Apple scab resistance is the most preferred one among the various management practices in containing the scab disease particularly under the circumstances of organic agriculture and the fungicide resistant pathogen strains (Schnabel and Jones, 2001; Martinez-Bilbao et al., 2012). Many resistant sources against apple scab has been reported from India and abroad, mainly based on evaluation of apple genotypes under natural conditions (Mercier et al., 2000; Blazek et al., 2003; Lefrancq et al., 2004; Gelvonauskiene et al., 2006; Brun et al., 2008; Biggs et al., 2010; Farooqui et al., 2014). Such findings may sometime be erroneous owing to disease escape and mixed infection by many pathotypes of pathogen. In order to remove such unambiguity from present investigation, genotypes were evaluated under artificial conditions with all the races present in the valley. A few promising genotypes like American Apirouge, H27, Shireen and Firdous were resistant to all the races. However their consumer unacceptability renders them unfit for spatial and temporal deployment. However, presence of Rvi6 resistance gene in Shireen and Firdous can be exploited easily to bred scab resistance cultivars using the aid of marker assisted backcrossing. The other useful scab resistance genes present in the differentials (Rvi3 to Rvi13) seems good candidates for durable scab management as the genes were not defeated by the pathogen and will be harder for the pathogen to defeat them in combination. These genes can be combined together provided comprehensive breeding strategy will be initiated and such strategies has already begun in many countries (Bus et al., 1999; Gleichauf et al., 2009; Flachowsky et al., 2011). A few cultivars showed resistance response to one or the other race of pathogen suggesting a good source of scab resistance in the collection. In such cultivars, pathogen germinated and penetrated the host (class 1 to 3a) but, further growth of the fungus was halted resulting in the hypersensitivity, suggesting that resistance in these cultivars is not due to host barriers but rather existence of effective defence mechanism which nevertheless needs to be elucidated by further studies at molecular level. Quantitative scab resistance in apple genotypes is well documented (Gessler et al., 2006; Jha et al., 2010; Bowen et al., 2011; Bus et al., 2011) and might be present in the cultivars like White Dotted Red, Saharanpuri and H27 as they have been reported moderately resistant under natural conditions (Farooqui and Dalal, 2003). Apple cultivar Golden Delicious exhibited susceptibility against race (1). Since, this cultivars carries Rvi1 scab resistance gene which is easily breached by the race (1) and (1, 2).
Development of high quality disease resistant apple cultivars is the preferred long-term means of reducing fungicide use in apples, in the short term growers are restricted by the lack of suitable resistant cultivars. This study has provided information about the susceptibility of a range of apple cultivars to scab, which should be useful in planning integrated pest and disease management programs for Kashmir fruit growers. For example, this information can be used by growers when planning new plantings of apples. If growers consider that for their enterprise, it is important to minimise applications of fungicides, then they have a rationale for making a decision between cultivars. Growers of ‘organic’ fruit would also find this information helpful in planning new plantings. In addition, this information should be a useful guide to selecting the most susceptible cultivars to target the scouting activities in, thus saving time and money.
ACKNOWLEDGMENT
The authors are highly thankful to DST, India for financial assistance (SR/SO/PS-47/08).
REFERENCES
- Barbara, D.J., A.L. Roberts and X.M. Xu, 2008. Virulence characteristics of apple scab (Venturia inaequalis) isolates from monoculture and mixed orchards. Plant Pathol., 57: 552-561.
CrossRefDirect Link - Biggs, A.R., G.W. Sundin, D.A. Rosenberger, K.S. Yoder and T.B. Sutton, 2010. Relative susceptibility of selected apple cultivars to apple scab caused by Venturia inaequalis. Plant Health Prog.
CrossRefDirect Link - Blazek, J., J. Krelinova and J. Blazkova, 2003. Results of a trial with 17 chosen apple cultivars bred in the Czech Republic that was evaluated 1996-2002 at Holovousy. Vedecke Prace Ovocnarske, 18: 7-23.
Direct Link - Bowen, J.K., C.H. Mesarich, V.G.M. Bus, R.M. Beresford, K.M. Plummer and M.D. Templeton, 2011. Venturia inaequalis: The causal agent of apple scab. Mol. Plant Pathol., 12: 105-122.
CrossRefPubMedDirect Link - Brun, L., F. Didelot and L. Parisi, 2008. Effects of apple cultivar succeptibility to Venturia inaequalis on scab epidemics in apple orchards. Crop Protect., 29: 1009-1019.
CrossRefDirect Link - Bus, V., C. Ranatunga, S. Gardiner, H. Bssett and E. Rikkerink, 1999. Marker assisted selection for pest and disease resistance in the New Zealand apple breeding programm. Acta Horticulturae, 538: 541-547.
CrossRefDirect Link - Bus, V.G.M., E.H.A. Rikkerink, V. Caffier, C.E. Durel and K.M. Plummer, 2011. Revision of the nomenclature of the differential host-pathogen interactions of Venturia inaequalis and Malus. Annu. Rev. Phytopathol., 49: 391-413.
CrossRefPubMedDirect Link - Chevalier, M., Y. Lespinasse and S. Renaudin, 1991. A microscopic study of the different classes of symptoms coded by the Vf gene in apple for resistance to scab (Venturia inaequalis). Plant Pathol., 40: 249-256.
CrossRefDirect Link - Crosby, J.A., J. Janick, P.C. Pecknold, S.S. Korban and P.A. O'Connon et al., 1992. Breeding apples for scab resistance: 1945-1990. Fruit Varieties J., 46: 145-166.
Direct Link - Didelot, F., L. Brun and L. Parisi, 2007. Effects of cultivar mixtures on scab control in apple orchards. Plant Pathol., 56: 1014-1022.
CrossRefDirect Link - Ellis, M.A., L.V. Madden and L.L. Wilson, 1984. Evaluation of an electronic apple scab predictor for scheduling fungicides with curative activity. Plant Dis., 68: 1055-1057.
Direct Link - Farooqui, K. and A. Dalal, 2003. Breeding apples for resistance to scab-an eco-friendly strategy. Acta Horticulturae, 696: 33-37.
CrossRefDirect Link - Farooqui, K.D., G. Hussain and M.A. Mir, 2014. Evaluation of some scab resistant apple hybrids as early season genotypes. Applied Biol. Res., 16: 161-168.
CrossRefDirect Link - Flachowsky, H., P.M. Le Roux, A. Peil, A. Patocchi, K. Richter and M.V. Hanke, 2011. Application of a high-speed breeding technology to apple (Malus × domestica) based on transgenic early flowering plants and marker-assisted selection. New Phytol., 192: 364-377.
CrossRefPubMedDirect Link - Gelvonauskiene, D., A. Sasnauskas, B. Gelvonauskis, J.B. Siksnianiene, G. Sabajeviene and P. Duchovskis, 2006. Investigation of biological important traits of apple selections. Sodininkyste Darzininkyste, 25: 20-31.
Direct Link - Gessler, C., A. Patocchi, S. Sansavini, S. Tartarini and L. Gianfranceschi, 2006. Venturia inaequalis Resistance in Apple. Crit. Rev. Plant Sci., 25: 473-503.
CrossRefDirect Link - Giraud, T., P. Gladieux and S. Gavrilets, 2010. Linking the emergence of fungal plant diseases with ecological speciation. Trends Ecol. Evol., 25: 387-395.
CrossRefDirect Link - Gladieux, P., F. Guerin, T. Giraud, V. Caffier and C. Lemaire et al., 2011. Emergence of novel fungal pathogens by ecological speciation: importance of the reduced viability of immigrants. Mol. Ecol., 20: 4521-4532.
CrossRefPubMedDirect Link - Gleichauf, B., T. Koller, K. Hunziker, P. Duffy, J. Frey and M. Kellerhals, 2009. Modern methods of apple breeding. Obst-und Weinbau, 145: 10-13.
Direct Link - Guerin, F. and B. Le Cam, 2004. Breakdown of the scab resistance gene Vf in apple leads to a founder effect in populations of the fungal pathogen Venturia inaequalis. Phytopathology, 94: 364-369.
CrossRefPubMedDirect Link - Guerin, F., P. Gladieux and B.L. Cam, 2007. Origin and colonization history of newly virulent strains of the phytopathogenic fungus Venturia inaequalis. Fungal Genet. Biol., 44: 284-292.
CrossRefDirect Link - Heaton, J.B., S.R. Dullahide, L.B. Baxter and A.D. McWaters, 1991. Race determination of nine Australian isolates of the apple black spot fungus Venturia inaequalis. Australasian Plant Pathol., 20: 139-141.
CrossRefDirect Link - Jha, G., K. Thakur and P. Thakur, 2010. The Venturia apple pathosystem: Pathogenicity mechanisms and plant defense responses. J. Biomed. Biotechnol.
CrossRefDirect Link - Le Van, A.L., C.E. Durel, B. Le Cam and V. Caffier, 2011. The threat of wild habitat to scab resistant apple cultivars. Plant Pathol., 60: 621-630.
CrossRefDirect Link - Lefrancq, B., M. Lateur and A. Rondia, 2004. Screening method for polygenic scab resistance within an apple breeding programme: Relationship between early greenhouse screening test on young seedlings and their scab susceptibility in natural field conditions. Acta Horticulturae, 2: 793-797.
CrossRefDirect Link - MacHardy, W.E., D.M. Gadoury and C. Gessler, 2001. Parasitic and biological fitness of Venturia inaequalis: Relationship to disease management strategies. Plant Dis., 85: 1036-1051.
CrossRefDirect Link - Martinez-Bilbao, A., A. Ortiz-Barredo, E. Montesinos and J. Murillo, 2012. Venturia inaequalis resistance in local Spanish cider apple germplasm under controlled and field conditions. Euphytica, 188: 273-283.
CrossRefDirect Link - Mercier, V., F. Combe, G. Fauvel, G. Marboutie and Y. Lespinasse, 2000. A four year experiment on the development of mite populations on scab resistant apples. Acta Hortic., 525: 325-330.
CrossRefDirect Link - Padder, B.A., M.D. Shah, M. Ahmad, T.A. Sofi, F.A. Ahanger and A. Hamid, 2011. Genetic differentiation among populations of venturia inaequalis in kashmir: A North-Western State of India. Asian J. Plant Pathol., 5: 75-83.
CrossRefDirect Link - Padder, B.A., T.A. Sofi, M. Ahmad, M.D. Shah, A. Hamid, S. Saleem and F.A. Ahanger, 2013. Virulence and molecular diversity of Venturia inaequalis in commercial apple growing regions in Kashmir. J. Phytopathol., 161: 271-279.
CrossRefDirect Link - Parisi, L., V. Fouillet, H. Schouten, R. Groenwold and F. Laurens et al., 2004. Variability of the pathogenicity of Venturia inaequalis in europe. Acta Horticulturae, 663: 107-113.
CrossRefDirect Link - Patocchi, A., A. Frei, J.E. Frey and M. Kellerhals, 2009. Towards improvement of marker assisted selection of apple scab resistant cultivars: Venturia inaequalis virulence surveys and standardization of molecular marker alleles associated with resistance genes. Mol. Breed., 24: 337-347.
CrossRefDirect Link - Schnabel, G. and A.L. Jones, 2001. The 14α-demethylasse(CYP51A1) gene is overexpressed in Venturia inaequalis strains resistant to myclobutanil. Phytopathology, 91: 102-110.
CrossRefPubMedDirect Link - Sutton, T.B., 1996. Changing options for the control of deciduous fruit tree diseases. Annu. Rev. Phytopathol., 34: 527-547.
CrossRefDirect Link