
Bengyella Louis
Department of Biochemistry, University of Yaounde I, BP812, Yaounde, Cameroon
Sayanika D. Waikhom
Institute of Bioresources and Sustainable Development, Takyelpat, 795001, Imphal, Manipur, India
Wakambam M. Singh
Institute of Bioresources and Sustainable Development, Takyelpat, 795001, Imphal, Manipur, India
Narayan C. Talukdar
Institute of Bioresources and Sustainable Development, Takyelpat, 795001, Imphal, Manipur, India
Pranab Roy
Department of Biotechnology, Haldia Institute of Technology, 721657, Haldia, West Bengal, India
Plant Pathology Journal
Year: 2014 | Volume: 13 | Issue: 1 | Page No.: 18-27







ABSTRACT
Potato cultivation is hampered by a host of factors among which fungal diseases play a leading role. Nevertheless, our knowledge on phytofungal diversity inciting diseases at the early developmental stages of potato is limited at present to some few traditionally recognized pathotypes. In this present study, potato farms were surveyed for two and a half years and predominant fungi causing disease at the early stages of potato development were identified using standard taxonomic monographs and ribosomal DNA locus. It was found that Ascomycetes viz. Colletotrichum gloeosporioides, Cochliobolus lunatus, Aspergillus terreus, Aspergillus fumigatus, Nigrospora oryzae, Fusarium culmorum, Pestalotiopsis vismiae, Phomopsis asparagi, Dothiorella gregaria, Didymella cucurbitacearum, Stagonosporopsis cucurbitacearum and Penicillium citrinum incite diseases of potato at the early developmental stages. This study provide a comprehensive overview of the genetic diversity of Ascomycetes thriving on potato plants and evidence that Aspergillus species are harmful pathogens causing loses to potato cultivation.
PDF Abstract XML References Citation
Received: September 02, 2013;
Accepted: November 04, 2013;
Published: March 18, 2014
How to cite this article
Bengyella Louis, Sayanika D. Waikhom, Wakambam M. Singh, Narayan C. Talukdar and Pranab Roy, 2014. Diversity of Ascomycetes at the Potato Interface: New Devastating Fungal Pathogens Posing Threat to Potato Farming. Plant Pathology Journal, 13: 18-27.
DOI: 10.3923/ppj.2014.18.27
URL: https://scialert.net/abstract/?doi=ppj.2014.18.27
DOI: 10.3923/ppj.2014.18.27
URL: https://scialert.net/abstract/?doi=ppj.2014.18.27
INTRODUCTION
The continuing decrease in field surveillance program and mining of emergent pathogens has led to our poor understanding of Ascomycetes diversity and the threat they pose in potato (Solanum tuberosum L.) fields. Usually, the use of pathogen contaminated tubers for seed favour diseases at the early developmental stages of potato. Congruently, most of the organisms causing disease at the early developmental stages go unidentified because of incorrect diagnosis since all parts of the plant are usually affected with rare characteristic symptoms.
Common blight pathogens of potato are Phytophthora infestans (causing late blight diseases), Alternaria solani (causing early blight disease), Verticillium spp. (causing early dying disease), Helminthosporium solani (causing silver scab disease), Colletotrichum coccodes (causing black dot disease), Synchytrium endobioticum (causing wart disease), Fusarium spp. (causing Fusarium wilt disease) and Alternaria alternata (causing brown spot and black pit disease). Although, these pathogens causes early blight diseases, the term ‘early-blight’ is misleading because these diseases are often observed on older leaves at the late stages of development not at the early developmental stages of the plant. Based on this pre-existing data for early blight diseases, farmers, especially those of Burdwan, West Bengal, India, subjectively apply fungicides such as mancozeb, copper oxychlorides and mancozebmetalaxyl to control potato diseases (Bengyella et al., 2012). This subjective approach is usually based on the assumption that common phytofungi of potato prevails. In most cases, this control measure fails leading to substantial loses and thus; indebting farmers. Addressing this problem requires a strategic surveillance of potato field and identifying recurring phytofungi so that appropriate control measure can be established. In this study, potato fields were surveyed for phytofungi causing diseases at the early developmental stages of potato. Here, it is observed that Ascomycetes poses severe threat to potato farming.
MATERIALS AND METHODS
Study area, sampling and identification: The study was performed in potato farms of Burdwan District (23°14’N, 87°51’E, altitude 150 m, 102.1 km from Kolkata), West Bengal, India, during winter 2010 to 2012. The area receives mean annual rainfall of about 1173-1442 mm and temperature of 10-20°C during potato farming season. Only four to six week’s old potato plants were sampled following development of true leaves. Diseased plants that amounted to more than 5% of the sampled plot with uniform pattern of symptoms were considered for further studies. Diseased potato leaves and stems were excised and treated with 2% NaClO solution (2 min) and rinsed in sterile water with three changes. The tissue pieces were aseptically plated on PDA (HiMedia®) amended with 250 mg L-1 chloramphenicol and incubated at 25°C in dark. Developed colonies were further purified on V8 agar medium and subsequently identified morphologically based on standard monographs taxonomic keys. However, non sporulating isolates that could not be mapped into any taxonomic position but produced significant crop damage were only characterized molecularly. Microscopy observation was performed under a phase contrast Olympus BX61 microscope couple with DP7M5.0.0.5 software and an Olympus DP70 camera.
Pathogenicity test: Pathogenicity test were performed on three weeks old disease free potato plants (cv. Kufri Jyoti) grown in a 7 L capacity pots under greenhouse conditions. Autoclaved soil derived from a blend of rice stalk vermicompost and sand (1:2% w/w) was used. The soil was amended with 1 g NPK (1:1:1% w/w) fertilizer after one week of sprouting. A fungal suspension of 106 conidia mL-1 was sprayed on the leaves using a hand compression sprayer. Control plants were sprayed with sterile water only. Plants were covered with polyethylene bags (at near 100% humidity) and incubated for four days at 20±2°C. Fungi isolates that produced disease symptoms similar to the field symptoms were re-isolated and characterized further using molecular tools.
DNA phylogenetic analysis: Total genomic DNA was extracted from mycelium mat using UltraCleanTM Microbial DNA isolation kits (Mo Bio Laboratories, Inc., Carlsbad, CA, USA) essentially following the manufacturer protocol. The purity of DNA was checked by the absorbance measurements at 260/280 nm using a Shimadzu® BioSpec-nanodrop spectrophotometer. The integrity of the DNA was confirmed by agarose gel electrophoresis. The ribosomal DNA internal spacer region 2 was amplified using ITS4 (5’-tcctccgctt attgatatgc-3’) and ITS3 (5’-gcatcgatgaag aacgcagc-3’) primers. The PCR mix contained 10 ng DNA, 5 μL Green GoTaq® reaction buffer (Promega®), 0.2 mM of each deoxyribonucleoside triphosphate (dNTP), 0.2 μM of each primer and 1.1 U of GoTaq® DNA polymerase (Promega®) in a total reaction volume of 25 μL in triplicates (conditions: 5 min at 95°C; 38 cycles of 1 min at 94°C, 1 min annealing at 56°C, 2 min for extension and a final 5 min at 72°C). The quality of the amplification products was checked on 2% agarose gel electrophoresis. The amplicons were purified and sequenced. The sequences have been submitted to the GenBank® nucleotide database based on 99.5 to 100% sequence similarity and have been awarded accession numbers (Table 1). To establish the diversity among these pathogens, sequences were aligned in Muscle software (Edgar, 2004). Substitution model parameters were determined based on Akaike Information Criterion, corrected (AICc) and Bayesian Information Criterion (BIC). The phylogenetic tree was inferred using the maximum likelihood method in MEGA5.2 software (Tamura et al., 2011).
RESULTS AND DISCUSSION
Among the diseases recorded, only three cases of early blight caused by Alternaria species were observed in the month of November to December (Table 1) and the conidial morphological variations are represented (Fig. 1). Alternaria blight caused by A. alternata has been reported in India on Rumex vesicarius (Sankar et al., 2012) and in neighbouring Pakistan (Siddiqui et al., 2009). However, based on the literature Alternaria solani (Fig. 1a) is herein reported for the first time to cause early blight disease of potato in India. Symptoms under greenhouse conditions were scattered brown spots on leaflet which often coalesced. Because the occurrence of Alternaria blight was low within the two and a half years surveillance, this indicated other unidentified pathogens are actively involved accounting for substantial loses of potato plants at the early developmental stages.
Following inoculation of Colletotrichum gloeosporioides under greenhouse condition, atypical symptoms similar to those of the field developed. No symptoms were observed on control plants. Often, plant progressively shriveled followed by discoloration (Fig. 2). Here, damping off and curling of apical leaves associated with occasional black spot on decolorized leaves were observed. This disease was encountered once in the field only in the month of October (Table 1). Colletotrichum gloeosporioides has been reported on Jasmimum grandiflorum in India (Sharma et al., 2012). To our knowledge, this is the first report of Colletotrichum gloeosporioides causing disease of S. tuberosum L. in India.
 | |
| Fig. 1(a-c): | Diversity of Alternaria, the main causal of early blight diseases identified in Burdwan potato farms, (a) Alternaria solani, (b) Alternaria sp. and (c) Alternaria sp. cultured on V8 agar medium |
| Table 1: | Ascomycetes causing diseases of potato at the early development stages reveals high occurrence level in the month of November and December |
 | |
| *Isolates that were characterized morphologically solely | |
Among all the pathotypes identified, the occurrence of Cochliobolus lunatus (Fig. 3) was high. C. lunatus has been reported to cause severe foliar necrosis at all stages of potato development (Bengyella et al., 2013a).
Under field conditions Cochliobolus lunatus, Aspergillus spp., Colletotrichum gloeosporioides and Alternaria solani caused significant losses of about 46-60%.
 | |
| Fig. 2: | Potato disease characterized by damping-off and occasional black spots caused by Colletotrichum gloeosporioides |
 | |
| Fig. 3: | A typical conidia morphological variations of Cochliobolus lunatus (GenBank accession JX477595) cultured on V8 agar medium |
Disease caused by Aspergillus spp. and Penicillium citrinum were all similar in nature to those of Aspergillus terreus under greenhouse conditions as described in Bengyella et al. (2013b). Nonetheless, A. terreus was more virulent and symptoms developed within three days of inoculation (Fig. 4).
 | |
| Fig. 4: | Leaf blight of potato caused by Aspergillus terreus characterized by brown apex, spots and chlorosis |
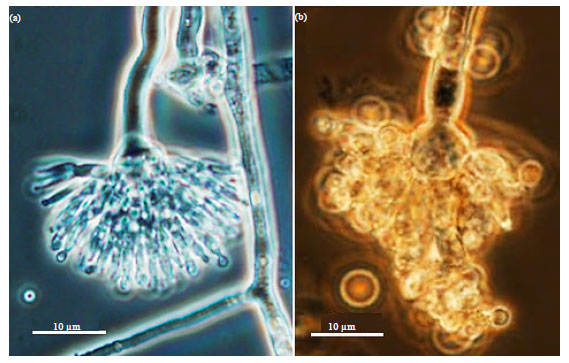
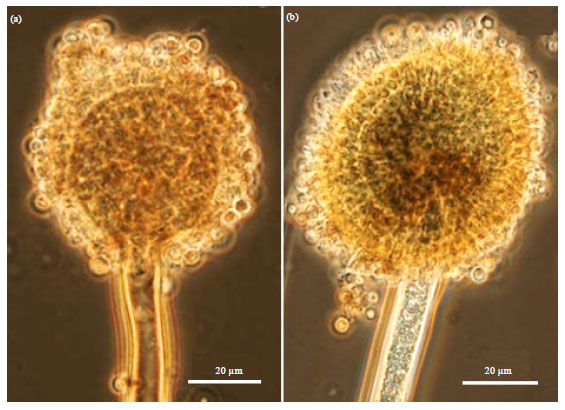
Although Aspergillus spp. and Penicillium spp. are often considered as weak pathogen of plants (Gugnani, 2003; Pitt and Hocking, 1997), it was found that Penicillium citrinum (Fig. 5a), Aspergillus aculeatus (Fig. 5b), Aspergillus terreus (Fig. 6a), Aspergillus sydowi (Fig. 6b), Aspergillus fumigatus (Fig. 7a) and Aspergillus niger (Fig. 7b) were successfull in inciting diseases of potato.
Importantly, it was observed that Didymella cucurbitacearum, Stagonosporopsis cucurbitacearum, Diaporthe phaseolorum, Nigrospora oryzae, Phomopsis asparagi and Gibberella zeae failed to incite diseases under greenhouse conditions. This result suggested that these pathogens required some abiotic or biotic factors to thrive on the plant which was excluded under greenhouse conditions or were simply opportunistic pathogens. Furthermore, these pathogens were associated with decomposition of plant tissues. Diseases caused by the Fusarium sp. under greenhouse conditions were severed. For instance, Fusarium culmorum caused stem wilting, stunting, chlorosis and leaf necrosis (Fig. 8). The pathogenicity of the pathogen developed slowly and disease persisted to the advanced stages of development, colonizing the xylem vascular tissues and producing browning of stem tissues as indicated by arrows (Fig. 8).
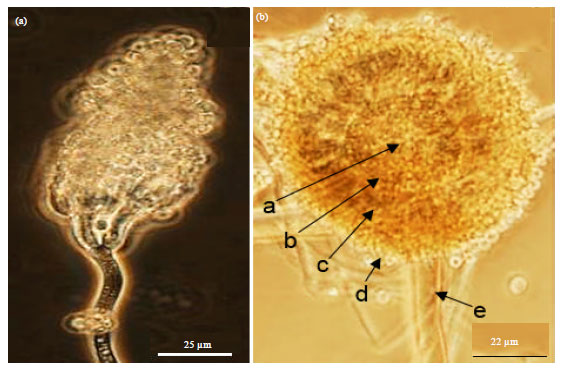 | |
| Fig. 5(a-b): | (a) A head of typical Penicillium citrinum and (b) A typical conidiophore head of Aspergillus aculeatus consisting of a central vesicle (a) which produces a dense layer of cells called metulae (b) which develps into phialides (c) from which conidia develops (d) and the whole structure is supported by stipe (e) Isolates were cultured on V8 agar medium and imaged at 1000X |
 | |
| Fig. 6(a-b): | (a) A typical conidiophore head of Aspergillus terreus characterized by a smooth columnar broom-like conidiophore and (b) Aspergillus sydowii characterized by slightly brownish conidiophore and spherical conidia (2.84 μm), Isolates were cultured on V8 agar medium and capture at 1000X |
 | |
| Fig. 7(a-b): | (a) Conidiophore head of Aspergillus fumigatus and (b) Conidiophore head Aspergillus niger, Isolates were cultured on V8 agar medium and capture at 1000X |
 | |
| Fig. 8: | Blight disease characterized by wilting under greenhouse conditions caused by Fusarium culmorum strain btl18IBSD GenBank accession KC937044, Image acquired with Nikon CoolpixS6200 under greenhouse condition |
According to the literature F. oxysporum variants has been reported in India on potato (Mandhare et al., 2011), but not F. culmorum. This is a first report demonstrating that F. culmurum causes disease of potato in India.
To date, Dothiorella gregaria (Fig. 9) is essentially reported to cause Dothiorella canker disease of avocado (Hartill and Everett, 2002). In potato the pathogen caused leaf discoloration associated with apex distortion. Often, chlorosis originated on or near the leaf apex (Fig. 10).
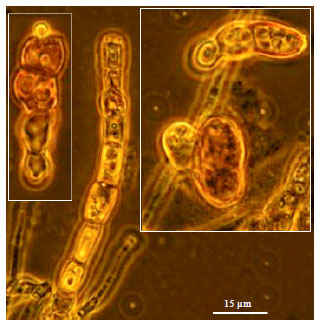 | |
| Fig. 9: | A typical conidia morphological variations of Dothiorella gregaria (GenBank accession KC937051) cultured on V8 agar medium imaged at 1000X |
It was observed that the disease does not reduce the plant vigour; however, surface discoloration could amount to 30-50% of the total leaf surface. This observation indicated that photosynthesis is severely hampered. To our knowledge, this is the first report of foliar disease of potato caused by Dothiorella gregaria in India.
It is worth noting that all the pathotypes belonged to the phylum Ascomycota. In essence Penicillium, Aspergillus and Alternaria, Cochliobolus produced huge numbers conidia which on a putative host lead to rapid colonization; however, does not explain the gain of new host. Primitive genes involved in pathogenicity are enzymatic such as cellulose encoders for plant cell wall decomposition. Meanwhile, advance pathogenicity and virulence factors are toxins. Ascomycota such as Penicillium, Aspergillus, Alternaria, Cochliobolus, Fusarium has been shown to produce host specific toxins (Yoder et al., 1997; Salas et al., 1999) and concurrently exhibit high level of virulence. Meanwhile, certain factors such as temperature fluctuation stress associated with insects damages and drought can readily weaken plant resistance to these emerging pathogens; thereby facilitating fungal invasion and subsequent toxin production (McMillian et al., 1998; Salas et al., 1999).
 | |
| Fig. 10: | Leaf discoloration of potato caused by Dothiorella gregaria under greenhouse condition and image was acquired with Nikon CoolpixS6200 |
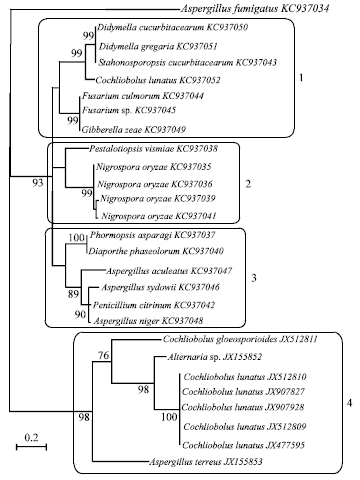 | |
| Fig. 11: | Molecular Phylogenetic analysis by Maximum Likelihood method. The ribotype tree is drawn to scale and was inferred by using the Maximum Likelihood method based on the General Time Reversible (GTR+G+I) model (Nei and Kumar, 2000), Branches are supported by 1000 bootstrap test of replicates and Aspergillus fumigatus strain btl81BSD KC937034 forms the out-group |
Some of the above mention factors were absence under the greenhouse condition and could explain why some pathotypes failed to incite disease. Importantly, Thompson (1994) argued that Ascomycetes are excellent ‘ecological opportunist’, suggesting in a given biota, they could coevolved and thrived on any putative host.
Final sequence alignment matrix for 5.8S-ITS2-28S rDNA locus provided 355 information patterns out of a total of 393 sites and 84 sites without polymorphism (21.37%). The GTR+G+I nucleotide substitution model was used base on AICc (2801.958) and BIC (3160.941). The rate of nucleotide transitional substitution were (A-G) = 10.28, (C-T/U) = 10.28, (T/U-C) = 10.28 and (A-G) = 10.28. Initial tree(s) for the heuristic search were obtained by applying the Neighbor-Joining method to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach. A discrete Gamma distribution was used to model the evolutionary rate differences among sites (5 categories (+G, parameter = 8.5974)). The rate variation model allowed for some sites to be evolutionarily invariable ((+I), 0.0000% sites). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (Fig. 11). The analysis involved 27 fungal isolates identified and sequenced. All positions containing gaps and missing data were eliminated. The percentage of trees in which the associated isolates clustered together is shown next to the branches following 1000 bootstrap test of replicates.
More than 90% of pathogens that thrives on S. tuberosum L. under field conditions goes identified. This is probably because fungicides are applied sequentially in most cases at the early stages of development till harvest in order to optimize yield. As a consequence, necrotrophs and heminecrotrophs which are chiefly Ascomycetes are severely suppressed, favouring the detection of biotrophs such as the Oomycetes which are specific to potato. Unequivocally, fungicide application eliminates the episode of Ascomycetes host gains as well as their pathogenicity evolutionary history. Base on this argument, pathogens that have coevolved with host or switch host to adapt in their biota in a given farming season eludes detection and characterization. Based on the ribotype tree (Fig. 11) clade 1 was made of Pezizomycetes, Sordariomycetes and Dothideomycetes, clade 2 was composed of Pezizomycetes and Sordariomycetes, clade 3 was composed of Sordariomycetes and Eurotiomycetes and clade 4 was made of Sordariomycetes, Dothideomycetes and Eurotiomycetes (Table 1). Previously, robust phylogenetic analysis revealed plant pathogens were concentrated at the level of Sordariomycetes while animal pathogens were focused in the Eurotiomycetes (Berbee, 2001). Our result show changes in the pathogenicity host-gains of Ascomycota; where Pezizomycetes, Sordariomycetes, Eurotiomycetes and Dothideomycetes are involve in inciting diseases of potato.
It was observed that all Cochliobolus lunatus strains failed to cluster indicating divergent evolutionary pattern. Akin to this pattern, Aspergillus terreus and Aspergillus fumigatus failed to cluster with other Aspergilli. Among all the identified fungi, Cochliobolus lunatus, Aspergillus sp. and Fusarium sp. has been reported to cause disease in both plants and animals. And as such, they represent potent dangers to many organisms and are generally called cross-kingdom pathogens (Berbee, 2001). Agricultural loses in potato caused by Ascomycota can sum up to be significant to those of Oomycetes such as Phytophthora spp. if the latter is controlled and the former is neglected. A significant damage caused by an Ascomycota is the Great Bengal rice famine caused by Cochliobolus spp. that lead to the loss of over 2 million lives (Scheffer, 1997). This case demonstrates the destructive potential of Ascomycetes when uncontrolled.
An efficient integrated disease management strategy to protect crops can only be implemented when the prevailing pathogens are identified. In this study, newly identified pathogens of potato are Cochliobolus lunatus, Aspergillus terreus, Colletotrichum gloeosporioides, Aspergillus fumigatus, Nigrospora oryzae, Fusarium culmorum, Pestalotiopsis vismiae, Phomopsis asparagi, Dothiorella gregaria, Didymella cucurbitacearum, Stagonosporopsis cucurbitacearum and Penicillium citrinum. Future surveillance of potato farms untreated with fungicides would supplement the current knowledge of diversity of Ascomycota and the magnitude of damages they caused in potato farms.
ACKNOWLEDGMENTS
This study was funded by The Academy of Sciences for Developing World (TWAS) and Department of Biotechnology (DBT), Government of India (Program No. 3240223450). The authors extend their gratitude to D.K. Hore, R.C. Rashmi and D.G. Momin for proofreading the text.
REFERENCES
- Sharma, P., N. Singh and O.P. Verma, 2012. First report of Colletotrichum gloeosporioides on Jasminum grandiflorum in India. J. Plant Prot. Res., 52: 91-92.
CrossRefDirect Link - Berbee, L.M., 2001. The phylogeny of plant and animal pathogens in the Ascomycota. Physiol. Mol. Plant Pathol., 59: 165-187.
CrossRef - Edgar, R.C., 2004. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res., 32: 1792-1797.
CrossRefDirect Link - Bengyella, L., R. Pranab, E.L. Yekwa and S.D. Waikhom, 2012. The farmers cry: Impact of heat stress on Fusarium oxysporum f. sp. dianthi, interaction with fungicides. Asian J. Plant Pathol., 6: 19-24.
CrossRef - Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei and S. Kumar, 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol. Biol. Evol., 28: 2731-2739.
CrossRefPubMedDirect Link - Gugnani, H.C., 2003. Ecology and taxonomy of pathogenic Aspergilli. Frontiers Biosci., 8: S346-S357.
PubMed - Sankar, N.R., M.N. Devamma and D. Giridhar, 2012. First report of Alternaria alternata causing leaf spot on Rumex vesicarius in India. Aust. Plant Dis. Notes, 7: 17-18.
CrossRef - Siddiqui, I., R. Bajwa and A. Javaid, 2009. First report of Alternaria alternata causing leaf spot on Rumex dentatus in Pakistan. Plant Dis., 93: 431-431.
CrossRef - Hartill, W.F.T. and K.R. Everett, 2002. Inoculum sources and infection pathways of pathogens causing stem‒end rots of 'Hass' avocado (Persea americana). New Zealand J. Crop Hort. Sci., 30: 249-260.
CrossRef - Nei, M. and S. Kumar, 2000. Molecular Evolution and Phylogenetics. Oxford University Press, United Kingdom, ISBN-13: 9780195350517, Pages: 352.
Direct Link - Yoder, O.G., V. Macko, T.J. Wolpert and B.G. Turgeon, 1997. Cochliobolus spp. and their Host-Specific Toxins. In: The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research, Corroll, G.C., P. Tudzynski and P.A. Lemke (Eds.). Vol. 5, Springer, Germany, pp: 145-166.